Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Клеточный цикл
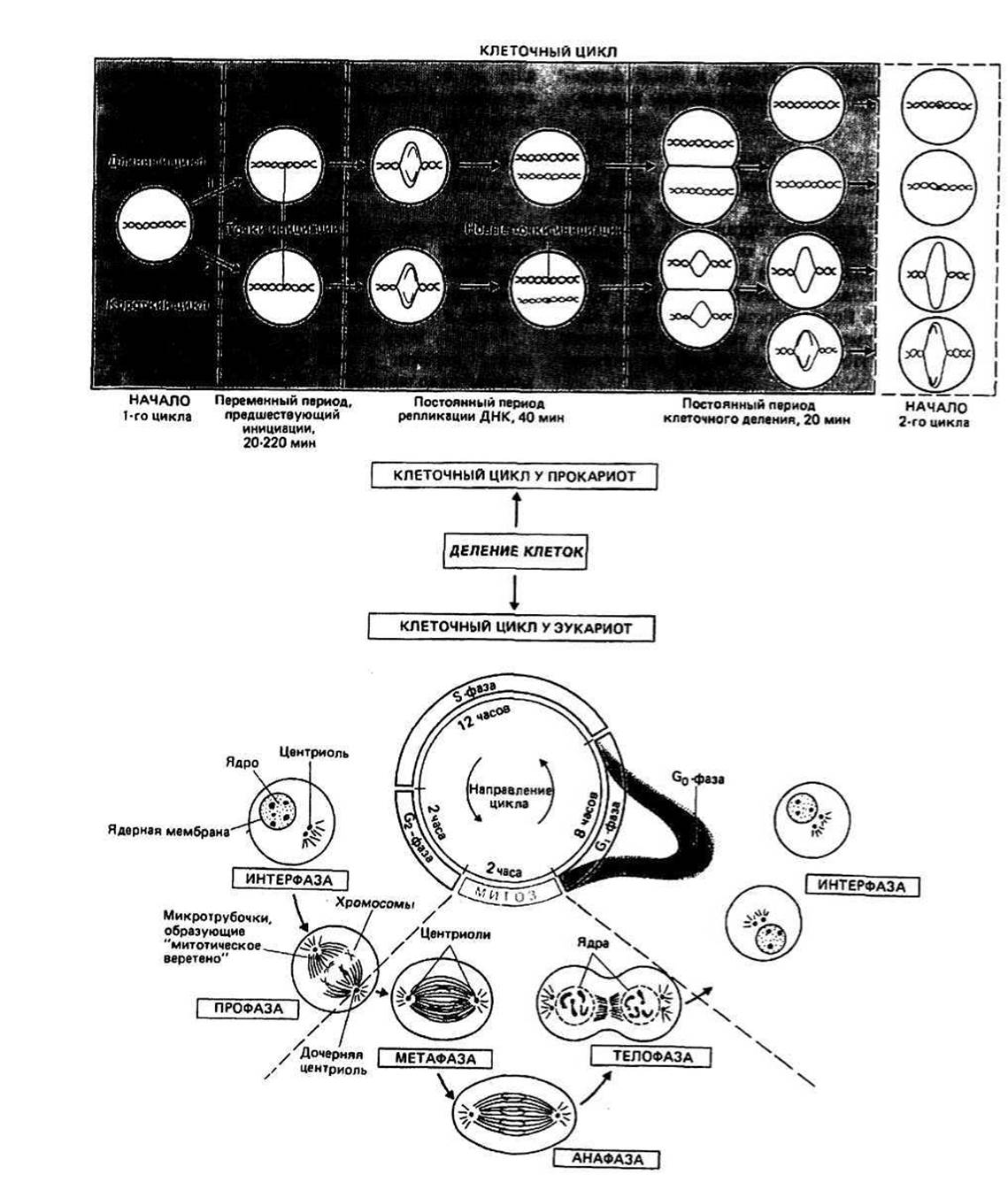
Рис. 29.1. Клеточный цим.
Клеточный цикл — это последовательность событий, происходящих во время деления клетки, в результате которой образуются две дочерние клетки. Элементы клеточного цикла прокариотических клеток пока охарактеризованы недостаточно полно. Напротив, для эукариот четко установлен набор событий в клетке, порядок которых в цикле фиксирован, но время протекания может сильно варьировать.
ДЕЛЕНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ включает репликацию единственной кольцевой хромосомы и всех содержащихся в клетке плазмид, а также удвоение клеточных органелл, в том числе плазматической мембраны и компонентов клеточной стенки. Скорость деления бактерий зависит от содержания питательных веществ в среде, в которой растут клетки. Если недостает каких-либо важных субстратов, уменьшается скорость инициации репликации; при этом скорость самой репликации не меняется.
Продолжительность репликации у большинства прокариотических клеток на удивление постоянна (около 40 мин). Если удвоение ДНК началось, оно продолжается до конца, независимо от любых изменений в составе питательной среды, произошедших в это время. Время, предшествующее инициации репликации ДНК, непостоянно (20—220 мин). В двух примерах на рис. 29.1 отдельно показаны длинный и короткий циклы. Длинный цикл имеет место тогда, когда велико время до начала инициации. В этом случае после начала репликации новые точки инициации в хромосоме не возникают до тех пор, пока не осуществятся все события клеточного цикла. Время, необходимое для появления новых точек инициации, непостоянно, и от него в основном зависит полная длительность клеточного цикла. Короткий цикл имеет место тогда, когда новые точки инициации возникают до полного завершения цикла репликации. Таким образом, еще до окончания деления клетки начинается следующий цикл репликации.
Инициация репликации осуществляется с помощью специфических белков. Если клетки поместить в среду без какого-либо ключевого субстрата, например триптофана, синтез белка сразу остановится. Хотя репликация ДНК и деление клетки продолжаются, новый цикл репликации не начинается до тех пор, пока опять не начнется синтез белка. Отсюда следует, что инициаторы — это белки, взаимодействующие с ДНК и переводящие ее в такое состояние, в котором она способна связывать ДНК-полимеразу.
Деление клетки происходит, когда ее суммарная масса достаточна для двух клеток и когда завершилась репликация. Время, затрачиваемое на процесс деления, относительно постоянно и составляет около 20 мин.
МИТОТИЧЕСКОЕ ДЕЛЕНИЕ ЭУКАРИОТИЧЕСКИХ КЛЕТОК включает в себя ряд фаз клеточного цикла, которые в сумме для одного цикла деления могут составлять около 24 ч.
Интерфаза соответствует отрезку цикла, в течение которого ядерный хроматин распределен по ядру и не удается выявить никаких признаков хромосом. Интерфазу подразделяют на четыре периода: Go, G1, S и G2.
Профаза следует за интерфазой и является тем периодом после G2, когда становятся различимыми конденсированные хромосомы.
Митоз (М) — это период за профазой, во время которого происходит перестройка хромосом, завершающаяся образованием двух ядер и, наконец, двух дочерних клеток. Выделяют три четко различимых подпериода митоза, известных как метафаза, анафаза и телофаза.
Фаза Go непостоянна по длительности. Клетка может перейти к ней сразу после деления. О клетке, находящейся в фазе Go, говорят как о покоящейся, т. е. о пребывающей в неделящемся состоянии. Клетки многих тканей взрослого организма находятся в основном именно в этом состоянии: в них не происходит репликации ДНК и может протекать лишь очень слабый синтез РНК. На переход клетки в фазу Go влияют такие факторы, как уменьшение количества питательных веществ, межклеточное контактное ингибирование деления и внутриклеточные регуляторы (например, в некоторых условиях циклический AMP). Когда клетка, находящаяся в фазе Go, подвергается действию определенных гормонов и других факторов роста, она может перейти из фазы Go в G1.
Фаза G1 — это период, в течение которого клетка готовится к началу синтеза ДНК, осуществляемого в течение фазы S. Длительность фазы G1 непостоянна (от нескольких часов до суток и даже больше). Почему это так, неизвестно, но при добавлении к клеткам в фазе G, гормонов продолжительность данной фазы часто уменьшается. Это можно объяснить увеличением скорости синтеза каких-то ключевых молекул под действием гормонов. Зачастую гормон должен находиться в клетке или на ее поверхности по крайней мере в течение 8 ч, пока сможет начаться репликация ДНК. Причина этого явления неизвестна. К концу фазы G1 клетка полностью подготовлена к переходу к S-фазе.
Фаза S— это период, за который количество ДНК удваивается; он предшествует делению клетки на две дочерние. Длительность фазы S незначительно меняется от клетки к клетке и составляет около 6—8 часов. За репликацией ДНК можно следить, измеряя в этот период скорость включения в ДНК тимидина, меченного тритием (3Н-Т). Одновременно происходит синтез РНК. Если к клеткам, находящимся в S-фазе, добавить 3Н-цитидин, то радиоактивная РНК появляется вначале в ядре, а затем в цитоплазме. Хотя РНК синтезируется во время всей интерфазы, наиболее интенсивно этот процесс протекает в S-фазе. Можно остановить клетки в S-фазе, если добавить ингибиторы, такие, как актиномицин D, который блокирует работу РНК-полимеразы, встраиваясь в ДНК (гл. 43). При этом клеточный цикл не может продолжаться, пока не завершится репликация ДНК. По завершении синтеза нуклеиновых кислот и после удвоения большинства других клеточных компонентов клетка переходит в фазу G2.
Фаза G2 — это часть интерфазы, во время которой не происходит репликации ДНК и может идти лишь очень слабый синтез РНК, т. е. G2 — это некий промежуточный период. В фазе G2 содержание хромосом в клетке вдвое больше, чем в нормальной диплоидной клетке. Длительность фазы G2 примерно 2—6 часов и относительно слабо зависит от типа клеток. В фазе G2 происходит конденсация хроматина и исчезает ядерная мембрана. Между G2 и М клетка проходит состояние профазы.
В профазе высококонденсированные хромосомы на самом деле оказываются состоящими из двух переплетенных, но раздельных структур, называемых хроматидами. Каждая хроматида — это завершенная копия двухцепочечной ДНК в комплексе с белками. Во время профазы центриоли, которые ранее поделились с образованием меньших, дочерних, центриолей, расходятся к противоположным полюсам. Виден сложный пучок микротрубочек (гл. 39), выходящих из центриолей, который называется митотическим веретеном. Оно образовано микротрубочками, расположенными между центриолями, и по форме напоминает яйцо. Эта структура выполняет в клетке функцию строительных лесов. Когда клетка входит в митоз, хромосомы связываются с «лесами», а ядерная мембрана перестает быть видимой. Хромосомы прикрепляются к микротрубочкам своими центромерами. Митоз (М) подразделяется на три периода: метафазу, анафазу и телофазу.
В метафазе хромосомы выстраиваются поперек митотического веретена, образуя метафазную пластинку. Механизм, лежащий в основе этой ориентации, неизвестен.
В анафазе хромосомы разделяются на составляющие их хроматиды. Каждая из хроматид от каждой пары движется к тому или другому концу митотического веретена. Движение хроматид, называемых теперь дочерними хромосомами, обусловлено сокращением микротрубочек, но детальный механизм этого процесса неизвестен. После завершения разделения хроматид клетка входит в телофазу.
В телофазе хроматиды деконденсируются, образуя дисперсный хроматин, и формируются ядерные мембраны. Митотическое веретено разрушается, а плазматическая мембрана перетягивается. В результате получаются две дочерние клетки. Затем каждая дочерняя клетка переходит в новый клеточный цикл в фазе Go или G1, и весь процесс повторяется.