Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Клетки и молекулы за работой
Нервы в действии
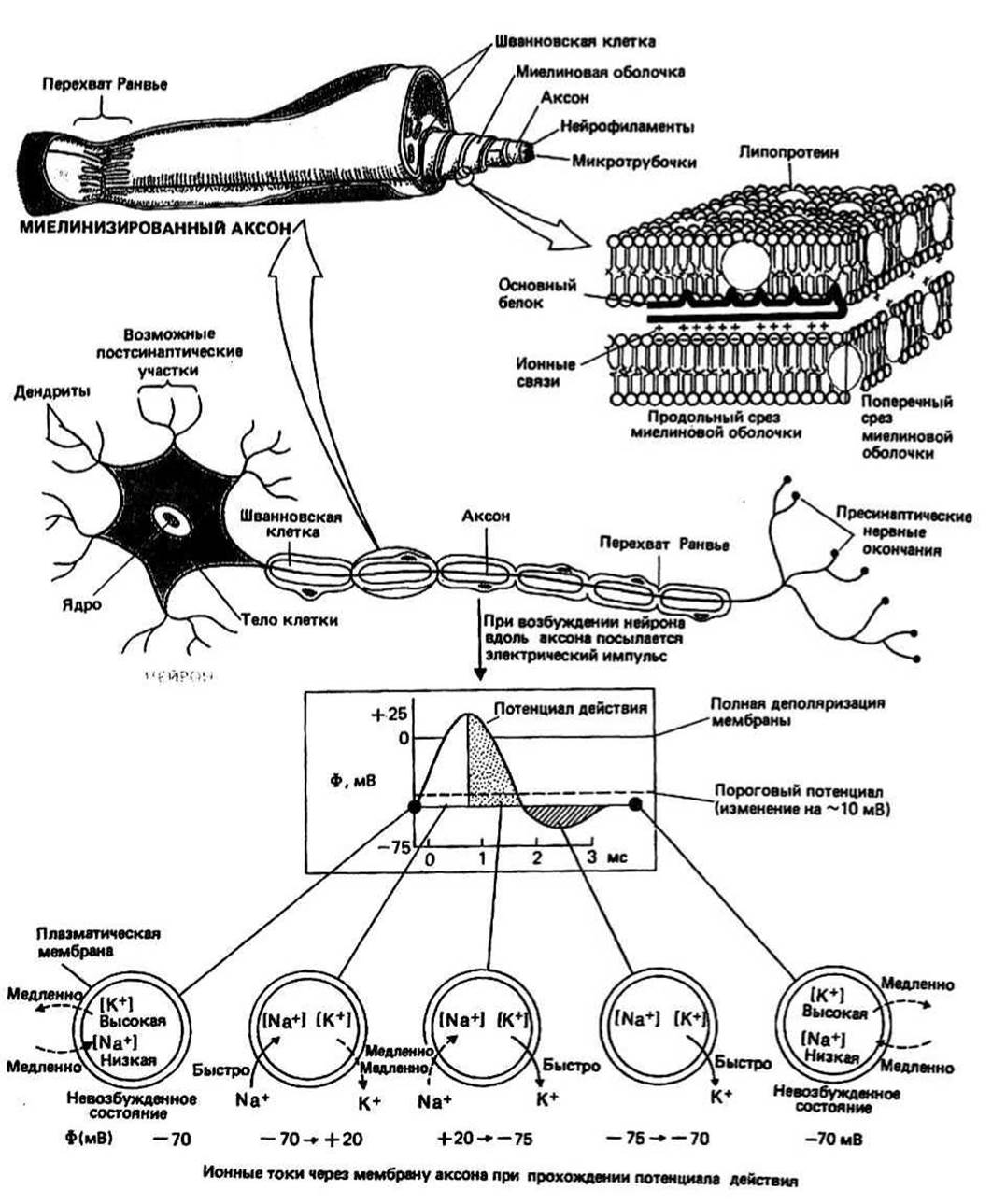
Рис. 36.1.
НЕРВЫ — длинные специализированные структуры, осуществляющие координацию функций организма и опосредующие его реакцию на различные воздействия. Нервные клетки (их называют также нейронами) есть у многих организмов. Каждая нервная клетка имеет тело и длинный отросток — аксон. Нервную систему позвоночных разделяют на центральную нервную систему (ЦНС) и периферическую нервную систему (ПНС). В ЦНС сходится огромное число нервов и осуществляется передача информации от одних нервов к другим. Два основных элемента ЦНС — головной и спинной мозг. ПНС состоит из двух частей: афферентных нервов, в которых аксоны идут от точек, воспринимающих внешние стимулы, к ЦНС, и эфферентных нервов, аксоны которых идут от ЦНС к тканям и органам-мишеням. Управляемые нервами органы и ткани называются иннервируемыми. Простейший механизм нервной регуляции — моносинаптический рефлекс.
МОНОСИНАПТИЧЕСКАЯ РЕФЛЕКТОРНАЯ ДУГА представляет собой нервную цепь, образованную двумя нейронами. Первый нейрон функционирует как принимающий элемент, к которому поступает стимул от чувствительной к растяжению области мышечного веретена: при растяжении мышцы механическое движение генерирует химический сигнал, который в свою очередь индуцирует электрический импульс, посылаемый по аксону этого первого, чувствительного нейрона (афферентный путь рефлекторной дуги). На конце аксона, расположенном в спинном или головном мозге, имеется особый контакт — синапс — со вторым, мотонейроном. Синапс состоит из пресинаптического элемента, которым является окончание афферентного нерва, и постсинаптического элемента, расположенного на теле мотонейрона. Эти два элемента разделены синаптической щелью. Посредством специального вещества, называемого нейромедиатором, импульс передается через синаптическую щель и затем распространяется по аксону мотонейрона (эфферентный путь рефлекторной дуги) к эффекторному окончанию, расположенному на мышце. Передача импульса к мышце через следующий синапс приводит к выделению кальция и сокращению мышцы (гл. 37). Таким образом растяжение мышечного веретена индуцирует рефлекторную сократительную реакцию мышцы.
Пресинаптический элемент — специализированная мембранная структура на конце нерва. В нервном окончании имеются пузырьки, содержащие нейромедиатор. Импульс, приходящий к окончанию, индуцирует слияние пузырьков с пресинаптической мембраной и выделение нейромедиатора в синаптическую щель.
Нейромедиаторы — небольшие заряженные молекулы, которые после высвобождения из пресинаптических мембран связываются с рецепторами на постсинаптической мембране. Примерами нейромедиаторов могут служить глутамат [+NH3-CH(CH2CH2COO)—СООН], встречающийся главным образом в синапсах ЦНС, и ацетилхолин [СН3—СОО—СН2—СН2— +N(CH3)3], использующийся преимущественно в ПНС.
Постсинаптический элемент образуется мембраной дендритов нервных клеток (в синапсах ЦНС) или чувствительной мембраной клеток ткани-мишени, иннервируемой мотонейроном (в синапсах ПНС). Мембрана мышечной концевой пластинки, заполненная множеством тесно расположенных рецепторов для нейромедиатора ацетилхолина, является постсинаптическим элементом периферического синапса на конце моносинаптической рефлекторной дуги.
Аксоны —длинные волокна, по которым передаются электрические сигналы от одного нервного окончания к следующему. У каждой нервной клетки есть только один аксон. Длина аксонов варьирует в широком диапазоне; аксоны многих мотонейронов, расположенных в ЦНС, доходят до самых удаленных частей иннервируемых ими мышц. Если аксоны покрыты изолирующей оболочкой, миелином (см. ниже), их называют миелинизированными. Диаметр аксонов в миелинизированных нервах варьирует в сравнительно узком диапазоне 1—20 мкм, тогда как немиелинизированные аксоны могут иметь диаметр от 0,1 до 500 мкм. Благодаря изолирующим свойствам миелина скорость прохождения импульса по миелинизированным нервам выше, чем по немиелинизированным. У млекопитающих, например, для миелинизированного нерва диаметром 20 мкм скорость проведения достигает 120 м/с, тогда как для немиелинизированного нерва диаметром 1 мкм она может составлять всего лишь 2 м/с. Отсутствие миелина может компенсироваться увеличением диаметра аксона: так, аксон кальмара (немиелинизированный) имеет диаметр 500—1000 мкм и проводит импульс со скоростью около 20 м/с.
Миелинизированные аксоны окружены специализированными глиальными клетками, вырабатывающими липопротеиновый мембранный миелин. Мембрана этих клеток наматывается на аксон, формируя многослойный чехол, который выполняет функцию изолятора. Для нейронов, входящих в состав ЦНС, образующими миелин клетками являются клетки олигодендроглии, а для нейронов ПНС — шванновские клетки. Аксон нейрона, изображенного на рис. 36.1, входит в состав периферического миелинизированного нерва. Миелиновая оболочка построена из липидной мембраны, состоящей из лецитин-холестерола и сфингомиелин-холестерола (гл. 32, 33), связанных главным образом с двумя белками, один из которых — липопротеин, а другой — основный белок. Основный белок находится на наружной поверхности мембраны (гл. 34) и благодаря этому может стабилизировать последовательные мембранные слои в миелиновом «рулете»: его положительно заряженный С-концевой участок, выступающий в межмембранное пространство, образует ионные связи с отрицательно заряженными липидами следующего мембранного слоя. Таким образом, каждый мембранный слой прочно удерживается в контакте с соседними слоями.
Проведение нервного импульса осуществляется в виде волны деполяризации, распространяющейся вниз по аксону, от тела клетки к нервному окончанию. В каждой точке невозбужденной мембраны аксона — до того как в нее придет волна деполяризации — имеется трансмембранный потенциал (ф) около —70 мВ (с внутренней стороны больше отрицательных зарядов). Этот потенциал обусловлен неравномерным распределением Na+ и К+ между внутри- и внеклеточным пространством (гл. 38) и избирательной проницаемостью мембраны. Когда в рассматриваемую точку приходит импульс, достаточно большой, чтобы вызвать изменение ф, превышающее пороговую величину (около 10 мВ), открывается ряд натриевых каналов, расположенных в плазматической мембране аксона. Этим начинается цикл изменений ф, называемый потенциалом действия. У миелинизированных аксонов плазматическая мембрана контактирует с содержащей Na+ внеклеточной жидкостью лишь в определенных точках — перехватах Ранвье — и деполяризация мембраны происходит только в области этих перехватов.
Потенциал действия возникает, когда импульс достигает перехвата и в аксон через чувствительные к трансмембранному потенциалу каналы входит Na+. Мембранный потенциал быстро (менее чем за 1 мс) изменяется от —70 мВ до 0 мВ (полная деполяризация) и далее до приблизительно +20 мВ (с внутренней стороны больше положительных зарядов). Этой фазе потенциала действия соответствует коричневая часть графика на рис. 36.1. Поскольку ионы Na+ переносят положительные заряды внутрь клетки, эффект, вызванный первоначально стимулирующим потенциалом, усиливается, и открывается еще больше натриевых каналов. Когда все каналы в перехвате будут
открыты, величина ф в нем достигнет максимума. Так как аксон покрыт изолятором — миелином, область электрического тока не может эффективно распространяться вдоль аксона по мембране, и ее распространение до следующего перехвата происходит в основном через цитоплазму аксона. Восстановление поляризации происходит в результате выхода из аксона ионов К+, который занимает несколько больше времени (2—3 мс), чем вход Na+ (этой фазе процесса соответствует на рис. 36.1 часть графика, покрытая точками). До тех пор пока реполяризация не закончится, перехват не может возбудиться снова (это так называемый рефрактерный период), а к тому времени, когда перехват опять будет способен к ответу, импульс уже уйдет вниз по аксону слишком далеко, чтобы вновь открыть в рассматриваемом перехвате натриевые каналы. По этой причине импульс распространяется вдоль аксона только в одном направлении. Заключительная стадия реполяризации начинается после того, как осуществляющие активный транспорт ионов АТРазы мембраны аксона восстановят нормальное соотношение концентраций Na+ и К+ (этой стадии соответствует заштрихованная область графика). Следует подчеркнуть, что электрические изменения на мембране аксона вызываются очень небольшими изменениями внутри- и внеклеточной концентрации ионов Na+ и К+. Критическим параметром является скорость движения этих ионов через мембрану; она указана на рис. 36.1 около стрелок, показывающих направление перемещения ионов. Время прохождения потенциала действия может составлять менее 1/5 с. У человека, например, рефлекторный ответ мышцы, разгибающей колено, занимает около 35 мс. Стимулы, поступающие к чувствительным нейронам, могут различаться по силе. Информация об интенсивности входного сигнала кодируется в виде частоты генерации потенциалов действия. Так, интенсивное сокращение мышцы индуцируется высокой частотой потенциалов действия, и наоборот.