Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Клетки и вирусы
ДНК-содержащие вирусы
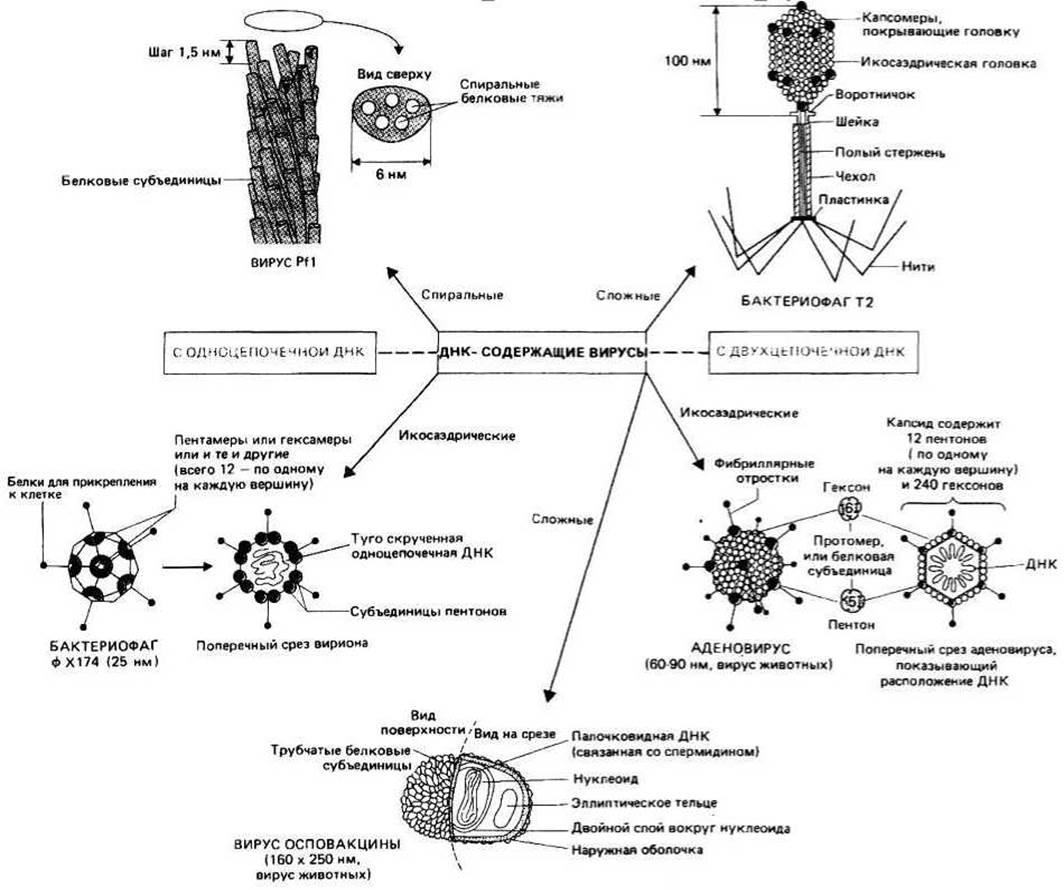
Рис. 4.1.
ВИРУСЫ — частицы, содержащие нуклеиновые кислоты, белки, а иногда и липиды и способные размножаться лишь в клетке-хозяине. Вне клетки вирусы не могут реплицироваться, поскольку у большинства из них нет ферментов, необходимых для полного воспроизведения зрелой вирусной частицы. Диаметр вирусных частиц (их называют также вирионами) равен 20—300 нм. Таким образом, они намного меньше, чем даже мельчайшие из прокариотических клеток. Так как размеры белков и некоторых нуклеиновых кислот находятся в диапазоне 2—50 нм, вирусную частицу можно было бы считать просто комплексом макромолекул. Вследствие их малых размеров и неспособности к самовоспроизведению вирусы часто относят к разряду «неживого».
ДНК-СОДЕРЖАЩИЕ ВИРУСЫ несут в качестве генетического материала либо одно-, либо двухцепочечную ДНК, которая может быть как линейной, так и кольцевой. В ДНК закодирована информация обо всех белках вируса. Вирусы классифицируют в зависимости от того, одно- или двухцепочечная у них ДНК и про- или эукариотической является клетка-хозяин. Вирусы, заражающие бактерии, называются бактериофагами.
Структура вируса в принципе такова: это молекула ДНК в белковой «обертке», называемой капсидом. Существует, однако, множество разных вариантов строения вирусов — от просто покрытой белком ДНК (например, бактериофаг РП) до сложных макромолекулярных комплексов, окруженных мембранными структурами (например, вирус оспы). Если у вируса есть мембрана, говорят, что он в оболочке, а если мембраны нет, вирус называют «раздетым». Различают четыре основных класса капсидов ДНК-содержащих вирусов: спиральные, икосаэдрические, сложные без оболочки, сложные с оболочкой.
Спиральные капсиды обычно встречаются у нитевидных вирусов. Они образуются путем самосборки асимметричных белковых субъединиц (капсомеров), объединяющихся в трубчатую структуру со спиральной симметрией (например, у РП). Субъединицы в большинстве случаев гомогенны, так что поверхность вириона состоит из множества копий одного и того же белка, хотя под наружным капсидом могут находиться и другие белки. ДНК в таких вирусах либо вытянута, либо может быть туго скручена в комплексе со специальными связывающими белками (гл. 26). Икосаэдрические капсиды свойственны большинству сферических ДНК-содержащих вирусов. Икосаэдр — это многогранник с двадцатью треугольными гранями, имеющий кубическую симметрию и приблизительно сферическую форму. Вершины треугольников, соединяясь, образуют соответственно двенадцать вершин икосаэдра; в местах соединения располагаются обычно пентамерные белковые структуры — пентоны; там же могут находиться участки, на которых формируются белковые нити, нередко ассоциированные с вершинами (например, у ф Х174 — см. рис. 4.1). Грани икосаэдра заполнены другими белковыми субъединицами, сгруппированными обычно в гексамерные структуры — гексоны (например, у аденовируса — см. рис. 4.1). Количество субъединиц, необходимое для заполнения граней, определяется размерами вириона в целом, и разные икосаэдрические вирусы содержат поэтому разное число гексонов — обычно при неизменном числе пентонов. ДНК обычно плотно свернута внутри капсида; иногда она связана с белками или полипептидами, способными стабилизировать ее структуру.
Сложные капсиды без оболочки типичны для бактериофагов: они состоят из частей с разными типами симметрии. У бактериофага Т2, например, ДНК находится в икосаэдрической головке, а для «узнавания» бактерии и введения в нее ДНК служат трубчатые и фибриллярные структуры (в узнавании участвует также лизоцим, расположенный на дистальном конце хвостового отростка).
Сложные капсиды с оболочкой есть только у вирусов эукариотических клеток. Они свойственны многим вирусам с нуклеоидом, состоящим из ДНК-белковых комплексов. Эти комплексы окружены одним или несколькими белковыми слоями, имеющими либо икосаэдрическую, либо нерегулярную симметрию, и наружной мембраной, почти все белковые компоненты которой являются по своему происхождению вирусными, а липидные структуры — клеточными. Инфицирование — процесс, посредством которого вирус внедряется в клетку-хозяина и «настраивает» ее метаболический аппарат на воспроизведение вирионов. Зараженные вирусом клетки либо остаются живыми (в этом случае говорят, что вирус невирулентен), либо подвергаются лизису, приводящему к высвобождению вирусных частиц. Неизменным итогом заражения клеток ДНК-содержащими бактериофагами является лизис. ДНК-содержащие вирусы животных вызывают лизис редко; клетки, однако, могут погибнуть из-за возникших при заражении хромосомных повреждений, вследствие иммунологической реакции организма или просто в результате нарушения вирусом нормальных клеточных функций.

Рис. 4.2. Обобщенная схема размножения ДНК-содержащих вирусов.
Размножение вируса — четко очерченный цикл, приводящий в конечном счете, после синтеза новых молекул вирусных белков и большого числа копий вирусной ДНК, к формированию зрелых вирусных частиц. Хотя детали этого процесса могут различаться у разных ДНК-содержащих вирусов, по существу он универсален. У вирусов бактерий весь цикл может завершаться менее чем за час, тогда как у многих вирусов животных он занимает не один день.
Адсорбция вируса на клетке-хозяине — первый этап инфицирования. Она происходит на специфических рецепторных участках (белковых или липидных) клеточной поверхности, которые узнаются особыми выступающими частями вириона и к которым он прочно прикрепляется. У вирусов без оболочки такими частями могут быть белковые отростки (например, у аденовируса и бактериофага Т2), а у вирусов с оболочкой это, как правило, белки, погруженные в вирусную мембрану. В процессе адсорбции осуществляются, в частности, такие белок-белковые взаимодействия, результатом которых является инициация стадии проникновения ДНК в клетку.
Проникновение вирусной ДНК в клетку-хозяина происходит у разных вирусов по-разному. ДНК многих бактериофагов (например, бактериофага Т2) попадает в клетку, как предполагается, следующим образом: белковый стержень сплющивается, подобно телескопической конструкции, и ДНК «вспрыскивается» в бактерию. Из вирусов животных ДНК обычно переходит в клетку в результате как бы слияния наружного слоя вириона с клеточной мембраной. Это легко себе представить для вирусов с оболочкой: должно произойти просто слияние мембран; в случае же вирусов без оболочки все не столь ясно. В отличие от ДНК большинство бактериофагов ДНК вирусов животных всегда входят в клетку вместе с непосредственно прилегающими к ней белками; последующее освобождение ДНК от этих белков осуществляется с помощью ферментов.
Для понимания процессов, описываемых ниже, читателю, возможно, понадобится изучить сначала часть Шэтой книги.
Транскрипция и репликации генетического материала вируса осуществляются обычно с участием ферментов клетки-хозяина. Сначала вирусная ДНК копируется РНК-полимеразами клетки-хозяина, в результате чего образуется мРНК, которая затем транслируется (гл. 25). На некоторых молекулах вирусной ДНК синтезируются также ее ДНК-копии — с помощью либо клеточной, либо кодируемой вирусом ДНК-полимеразы (гл. 20). Эти ДНК-копии используются впоследствии при сборке вирусных частиц. В некоторых случаях, например у бактериофага Т4, первые же новосинтезированные молекулы вирусной мРНК транслируются с образованием специальных белков, модифицирующих полимеразы клетки-хозяина таким образом, что те прекращают транскрипцию клеточных генов, не теряя при этом способности транскрибировать вирусные. В какой части клетки протекают процессы транскрипции и репликации вирусной ДНК: в ядре или в цитоплазме? Для бактериофагов такой дилеммы не существует; что же касается ДНК-содержащих вирусов животных, то, по-видимому, ни та, ни другая локализация не является для этой группы вирусов единственно возможной: у одних транскрипция и репликация происходят в ядре клетки-хозяина (например, у вируса герпеса), а у других — в цитоплазме (например, у поксвирусов).
Трансляция вирусной мРНК на рибосомах клетки- хозяина приводит к образованию вирусных белков. Некоторые из этих белков используются затем для построения капсидов, другие связываются с вирусной ДНК, вероятно, стабилизируя ее (у многих вирусов животных), а третьи, хотя и не войдут никогда в состав зрелых вирионов, участвуют в процессе их сборки в качестве ферментов (например, у бактериофага Т2).
Сборка вируса из его компонентов в клетке-хозяине может происходить спонтанно (и тогда она называется самосборкой), но может и зависеть от участия вспомогательных белков. Вирусная ДНК обычно покрывается слоем белка — капсидом. Капсид в свою очередь может заключаться в мембранную структуру, получаемую вирионом обычно от клетки-хозяина: покидая клетку путем отпочковывания от нее, вирусная частица оказывается окруженной плазматической мембраной.