Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Структура липидных мембран
Распределение липидов между цитозольной и экзоплазматической сторонами мембраны
Всем типам мембран присуща асимметрия в распределении липидов в пределах мембраны. Хотя большинство липидов присутствуют в обоих монослоях бислоя, их концентрация, как правило, в одном из монослоёв намного выше, чем в другом. Например, в плазматических мембранах эритроцитов человека практически все сфингомиелины и фосфатидил- холины (и те, и другие образуют относительно менее жидкие бислои) обнаружены только в пределах монослоя экзоплазматической стороны мембраны.
И, наоборот, фосфатидилэтаноламин, фосфатидилсерин и фосфатидилинозитол (которые образуют более жидкие бислои) преимущественно локализованы в цитозольном монослое мембраны. Такое распределение липидов между монослоями бислоя может влиять на кривизну мембраны (рисунок 27).
Холестерол, в отличие от фосфолипидов, практически одинаково присутствует в обоих монослоях бислоя мембраны.
Относительное содержание определённых фосфолипидов может быть определено по результатам их гидролиза ферментами фосфолипазами, которые разрезают различные связи в гидрофильных головках фосфолипидов (рисунок 28). Каждый тип фосфолипаз разрезает определённую связь (показаны на рисунке 28 стрелками). Если фосфолипазы добавлять в межклеточное пространство, то они не могут гидролизовать фосфолипиды цитозольной стороны плазматической мембраны, поскольку эти фосфолипазы не способны преодолеть мембрану и попасть в цитозоль. Фосфолипаза С (PLC) (рисунок 28) является цитозольной, и она гидролизует фосфолипиды с цитозольной стороны плазматической мембраны, оставляя в мембране диацилглицерол (рисунок 15(6)).
В настоящее время пока ещё не ясно, каким образом возникает асимметричное распределение фосфолипидов в мембране. В чистых бислоях фосфолипиды не совершают самопроизвольный перескок (флип-флоп) из одного монослоя в другой, поскольку такой переход предполагает перемещение полярной головки через гидрофобный слой мембраны, а этот процесс чрезвычайно энергетически не выгоден.
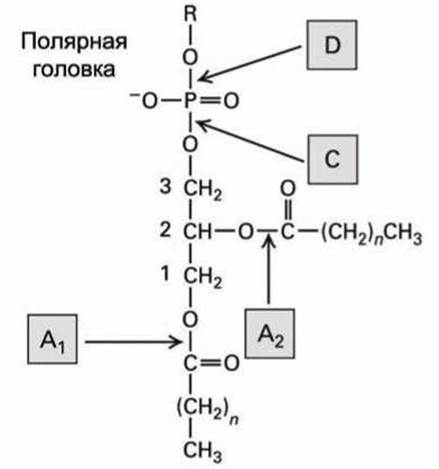
Рисунок 28 - Гидролиз фосфолипидов фосфолипазами А, С и D. Цифрами отмечены атомы углерода глицерола
Возможно, асимметрия в распределении фосфолипидов определяется векторным синтезом липидов в эндоплазматическом ретикулуме и аппарате Гольджи.
Сфингомиелин синтезируется на люминальной (экзоплазматической) стороне мембраны аппарата Гольджи, а фосфоглицериды, наоборот, синтезируются на цитозольной стороне мембраны эндоплазматического ретикулума, которая топологически эквивалентна цитозольной стороне плазматической мембраны.
Но такое предположение не объясняет преимущественное расположение фосфатидилхолина на экзоплазматической стороне мембраны.
Перемещение этого и других типов липидов из одного монослоя мембраны в другой катализируется особыми АТФ-зависимыми транспортными белками, которые называются флиппазами.
Избирательное расположение липидов на одной из сторон мембраны является физиологически необходимым для многих мембранных процессов. Например, полярные головки всех фосфорилированных форм фосфатидилинозитола (РІ) ориентированы в сторону цитозоля клетки. Некоторые из этих головок отделяются цитозольной фосфолипазой С, которая активизируется в результате действия на клетку многих гормонов.
Результатом такого действия фосфолипазы С является появление в цитозоле водорастворимого фосфоинозитола (IP), а в мембране липофильного диацшглицерола (diacyIglycerol, DAG) (рисунок 15(6)). Эти молекулы принимают участие в процессах внутриклеточной сигнализации.
Фосфатидилсерин тоже располагается преимущественно в цитозольном монослое плазматической мембраны. На первых стадиях стимуляции тромбоцитов сывороткой крови фосфатидилсерин на короткое время транслоцируется на экзоплазматическую сторону мембраны (по- видимому, флиппазой), где он активирует ферменты, обеспечивающие процесс сворачивания крови.