Биологические мембраны - А. Н. Огурцов 2012
Структура и функции биомембран
Активный транспорт
Протонные насосы F-класса
Насосы F-класса, использующие ток протонов для синтеза АТФ, называются АТФ-синтазами или комплексами F0F1.
Комплекс F0F1 или АТФ-синтаза состоит из двух главных компонентов F0 и F1 (рисунок 67), которые ещё называют мембранным и цитозольным моторами.
Мембранный компонент F0 состоит из трёх типов интегральных мембранных белков, которые обозначаются а, b и С. В клетках бактерий и дрожжей часть F0 состоит из одной субъединицы а, двух субъединиц b и десяти субъединиц С, поэтому состав компонента F0 записывают как а1b2c10.
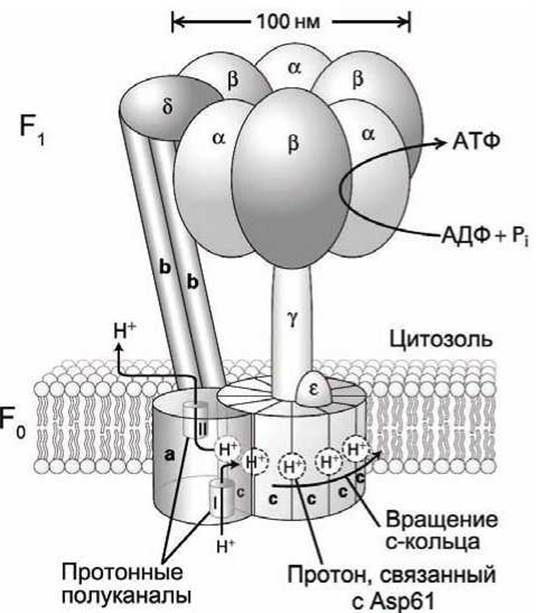
Рисунок 67 - Схема АТФ-ситнтазы в плазматической мембране бактерий
Компонент F0 митохондрий клеток животных содержит 12 субъединиц С, а хлоропластов растений — 14 субъединиц С. Субъединицы С образуют кольцо в плоскости мембраны. Субъединицы а и b прочно связаны между собой, но не связаны с кольцом субъединиц С.
Цитозольный компонент F1 является водорастворимым комплексом пяти различных полипептидов и имеет структуру а3β3уδє. В нижней части F1 находится субъединица є, которая прочно присоединена как к субъединице у, так и к нескольким субъединицам С компонента F0.
Субъединицы а и ß компонента F1 соединены друг с другом поочерёдно и формируют Колычевой гексамер aßaßaß или (aß)3, который расположен наверху субъединицы у. Субъединица δ прикрепляет этот гексамер к субъединицам b компонента F0. В результате а и b субъединицы компонента F0 и δ и (aß)3 гексамер компонента F1 образуют прочно связанную структуру, которая заякорена в мембране - своеобразный статор, который не участвует в механическом движении, и относительно которого вращается ротор, состоящий из субъединиц С мотора F0 и субъединиц у и є мотора F1.
Синтез АТФ. Протон через протонный полуканал I в субъединице а доходит до субъединицы С и связывается с аспартатом Asp61 этой субъединицы. После поворота с-кольца этот протон достигает полуканала II, который позволяет протону диссоциировать с аспартата и выйти в цитозоль.
Создаваемое таким потоком протонов вращение субъединицы у относительно неподвижных субъединиц a и ß циклически стимулирует конформационные переходы в каталитических субъединицах ß компонента F1 (рисунок 68).
Каждая каталитическая субъединица ß поочерёдно принимает три конформации.
1. О-конформация (open), которая слабо связывает АДФ и Рi, и не связывает АТФ;
2. L-конформация (Iow), которая сильнее связывает АДФ и Рi;
3. Т-конформация (tight), в которой АДФ и Рi связываются так сильно, что спонтанно формируют АТФ.
Дальнейшее вращение субъединицы у возвращает каталитическую субъединицу в исходную О-конформацию, в которой образованная молекула АТФ отсоединяется, и цикл начинается снова.
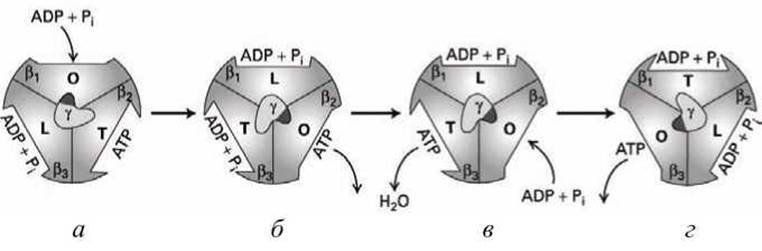
Рисунок 68 - Каталитический цикл АТФ-ситнтазы
Допустим АДФ и Рi связываются с субъединицей ß1 находящейся в О-конформации (рисунок 68(a)). Ток протонов проворачивает ротор АТФ-синтазы (и у-субъединицу) на 120° относительно неподвижных субъединиц ß (рисунок 68 (а)→(б)). В результате такого поворота способность субъединицы ß1 к связыванию АДФ и Рi возрастает с О до L, соответственно, у субъединицы ß3 возрастает с L до Т, а у субъединицы ß2 снижается с Т до О, что приводит к выходу образованной молекулы АТФ из субъединицы ß2.
На следующем этапе (рисунок 68 (б)→(в)) АДФ и Рi внутри субъединицы ß3, которая теперь имеет Т-конформацию, сливаются в молекулу АТФ, а к субъединице ß2 (в конформации О) присоединяются АДФ и Рi, В результате (рисунок 68(b)) образуется такой же комплекс F1 с метаболитами, как и вначале (рисунок 65(a)), но повернутый на 120°.
На следующем этапе (рисунок 68 (в)→(г)) происходит поворот ещё на 120° у-субъединицы, который снова стимулирует О→L→Т→О конформационные изменения субъединиц ß, описанные выше. Повторение этапов (а)→(б) и (б)→(в) приводит к образованию трёх молекул АТФ на каждый "полный оборот" у-субъединицы на 360°.
Молекулы АТФ или АДФ также связываются с регуляторными (или аллостерическими) центрами на трёх а-субъединицах. Такое связывание обеспечивает изменение скорости синтеза АТФ по мере изменения концентрации АТФ в матриксе, но не влияет непосредственно на синтез АТФ из АДФ и Рi.
Работа компонента F1 обратима. Вращение ротора так стимулирует каталитические субъединицы, что они катализируют синтез АТФ. В этом случае АТФ-синтаза работает как генератор АТФ. И наоборот, гидролиз АТФ стимулирует вращение ротора.
На рисунке 69 показаны конформационные изменения в компоненте F1 при вращении ротора. Изображена только одна каталитическая субъединица, а остальная часть мотора F1 показана контуром.
В верхних частях ß-субъединицы совместно формируют структуру, функционально подобную подшипнику, в которой закреплен конец ротора. Эти верхние части не изменяются при конформациях ß-субъединиц, сохраняя "подшипник". В нижней части рисунка 69 показаны два положения ротора. При вращении ротора изменяется конформация только нижней части каждой ß-субъединицы по отношению к верхней, неподвижной части. При этом стимулируется (прижимаются друг к другу) объединение АДФ и фосфата, и при дальнейшем повороте ротора готовая АТФ вытесняется из центра связывания.
Отметим, что мотор F1 не "запитывается" напрямую диссоциацией АТФ. Диссоциация АТФ обеспечивает необратимость процесса вращения на каждом этапе цикла АТФ-синтазы.
Когда АТФ-синтаза работает в качестве мотора, то молекуле АТФ легко связаться с каталитическими субъединицами, но сложно высвободиться, а молекуле АДФ, наоборот, сложно связаться, но легко высвободиться. АТФ, легко связавшись с АТФ-синтазой, гидролизуется и в виде АДФ легко покидает АТФ-синтазу.
При работе АТФ-синтазы в качестве генератора АТФ внешняя механическая энергия используется для реализации двух энергозатратных этапов: стимуляции связывания АДФ и стимуляции высвобождения АТФ.
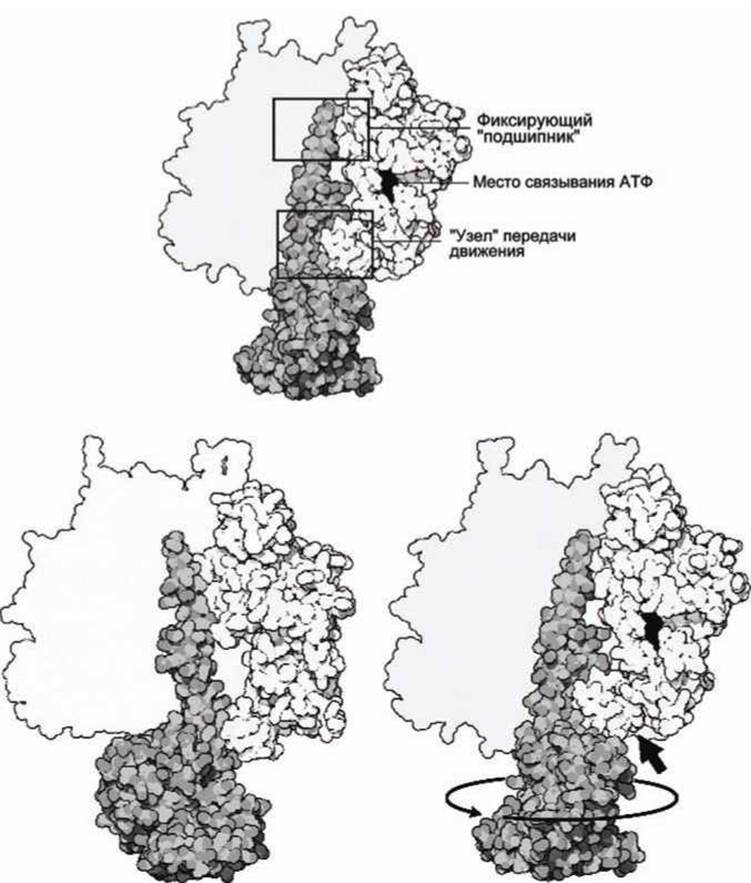
Рисунок 69 - Конформационные изменения в F1 моторе АТФ-синтазы.
Вращение ротора АТФ-синтазы. Принцип работы мотора F0, который в настоящее время считается наиболее обоснованным из множества выдвинутых гипотез и реализующий процесс вращательной диффузии, показан на рисунке 70.
Цилиндрическая трансмембранная часть ротора АТФ-синтазы, образованная субъединицами С, имеет отрицательно заряженные центры связывания протонов (радикалы аспарагиновой кислоты Asp61). Поскольку ротор погружен в мембрану, то он может вращаться, только если эти центры нейтрализованы протонами.
Протоны достигают ротора через один полуканал в статоре (на рисунке 70 схематически показан статорный домен, примыкающий справа к ротору) и, совершив круг вместе с ротором, выходят через другой полуканал в теле статора с противоположной стороны мембраны.
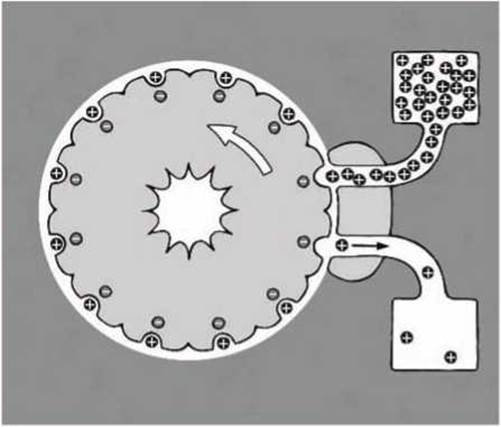
Рисунок 70 - Принцип работы мотора F0
В настоящее время подробно исследованы два варианта мотора F0, которые приводятся в движение потоками протонов или ионов натрия.
В принципе, цилиндрический ротор мотора F0 может совершать произвольные скачки в обоих направлениях, стимулированные тепловым движением. Для обеспечения однонаправленного вращения необходимо разрешать повороты в нужном направлении и блокировать повороты ротора в обратном направлении. Другими словами, мотор из произвольных случайных тепловых движений должен "отбирать" только "благоприятные" события или выполнять функцию "броуновского решета" (или "броуновского храповика", Brownian ratchet).
Отрицательно заряженные радикалы аспарагиновой кислоты в центрах связывания протонов ротора не позволяют ротору свободно вращаться в мембране.
Через полуканал в статоре (верхний полуканал на рисунке 70) протоны из области с повышенной концентрацией протонов проходят к ротору. В области, закрытой от мембраны статором, аспартаты ротора могут протонироваться, нейтрализуя тем самым свой отрицательный заряд.
Нейтрализованные домены ротора могут затем вращаться (против часовой стрелки, как показано на рисунке) внутри мембраны. После завершения оборота протонированный аспартат снова попадает в экранированную от мембраны статором область. Здесь находится второй полуканал, выходящий на противоположную сторону мембраны, где концентрация протонов ниже.
Протон диссоциирует с аспартата и выходит через полуканал, а оставшийся заряженный аспартат не позволяет ротору вращаться в обратном направлении (своеобразный молекулярный храповик, выполняющий функцию броуновского решета).
Весь этот процесс обратим.
Самопроизвольно мотор F0 будет захватывать протоны из полуканала, где их концентрация высока, и высвобождать их в полуканал, где концентрация протонов низка.
Однако если приложить к ротору постоянный вращательный момент в противоположном направлении, то ротор не сможет вращаться (теперь уже по часовой стрелке в направлении, противоположном указанному на рисунке 70 стрелкой) до тех пор, пока он не захватит протон из полуканала с низкой концентрацией протонов.
Завершив оборот, ротор высвободит протон в полуканал с высокой концентрацией протонов. В этом случае под действием внешней силы мотор будет работать как протонная помпа, перекачивающая протоны через мембрану.