Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Механизмы генерации потенциала действия
Строение потенциалочувствительных ионных каналов
Исследование молекулярной структуры потенциалочувствительных ионных каналов показало, что они имеют чрезвычайно подобное строение. Большинство исследованных потенциалочувствительных калиевых каналов имеют терамерную структуру и состоят из четырёх идентичных субъединиц, объединённых в общую функциональную структуру так, чтобы образовать центральный канал.
Каждая субъединица потенциалочувствительного калиевого (К+) канала состоит из шести трансмембранных а-спиралей, обозначаемых S1-S6 и сегмента Р (рисунок 130).
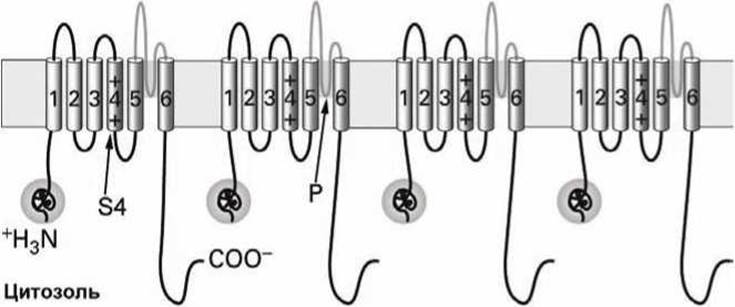
Рисунок 130 - Схема потенциалочувствительного калиевого канала
Фрагмент субъединицы, содержащий а-спирали S5, S6 и Р-сегмент структурно и функционально подобен субъединице нерегулируемого калиевого канала, обеспечивающего формирование и поддержание мембранного потенциала покоя (рисунок 91).
Трансмембранная а-спираль S4, которая содержит множество положительно заряженных лизинов и аргининов, выполняет функцию сенсора мембранного потенциала.
Глобулярный цитоплазматический домен, присоединённый к а-спирали S1 вблизи N-конца белковой цепи, выполняет функцию канал-инактивирующего (блокирующего) сегмента.
Потенциалочувствительные натриевые (Na+) и кальциевые (Са2+) каналы являются мономерными белками, состоящими из четырёх гомологичных доменов (рисунки 131, 132).
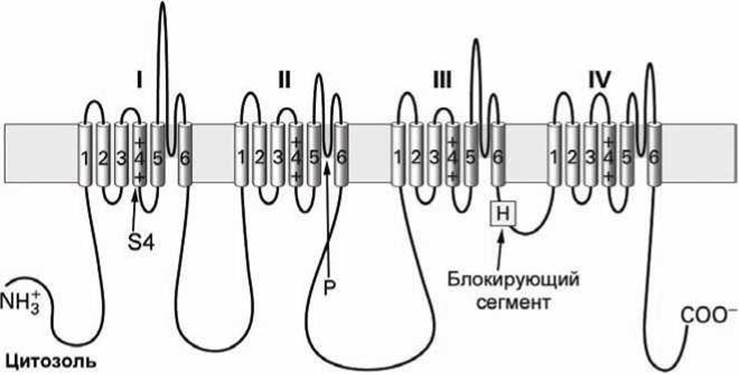
Рисунок 131 - Схема потенциалочувствительного Na канала
Каждый из этих доменов структурно и функционально подобен одной субъединице К+-канала (рисунок 130). Однако в отличие от потенциалочувствительных К+-каналов, у которых каждая из субъединиц имеет свой канал-блокирующий сегмент (а всего их четыре в функциональном К+-канале), мономерные потенциалочувствительные каналы имеют единственный такой блокирующий сегмент (Н).
Фильтр селективности канала образуется четырьмя парообразующими петлями (P-сегментами). На рисунке 132 показана схема взаимного расположения четырёх субъединиц Са2+-канала, каждая из которых состоит из шести трансмембранных а-спиралей; потенциалочувствительные а-спирали (сенсоры канала) отмечены штриховкой; порообразующие петли ориентированы внутрь поры в канале.
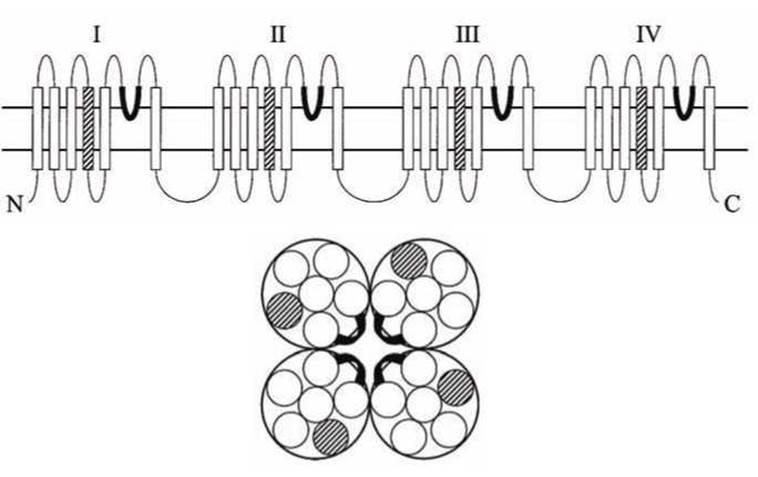
Рисунок 132 - Схема укладки элементов структуры потенциалочувствительного кальциевого канала
Таким образом, можно утверждать, что все ионные каналы и структурно, и функционально подобны, и, по-видимому, все они произошли от одного белка-предка, имевшего шесть трансмембранных а-спиралей.
Роль потенциалочувствительных сенсоров в потенциалочувствительных ионных каналах выполняют четыре положительно заряженные а-спирали S4, перемещение которых стимулирует конформационный переход белка-канала, при котором открывается или закрывается ионный канал (рисунок 128).
Собственно ворота ионного канала формируются N-концами спиралей S5 и С-концами спиралей S6 всех четырёх субъединиц.
Спонтанная инактивация ионных каналов через несколько миллисекунд после их открытия осуществляется с помощью специфических блокирующих сегментов, имеющих вид шарообразных положительно заряженных глобул на концах бесструктурных белковых нитей.
В состоянии покоя эти глобулы на N-концах четырёх субъединиц калиевых каналов свободно плавают в цитозоле.
Через несколько миллисекунд после открытия канала (после деполяризации мембраны) один из глобулярных шаров перемещается через боковое окно между двумя субъединицами канала и связывается в гидрофобном кармане центральной полости канала, блокируя поток ионов К+ (рисунок 133).
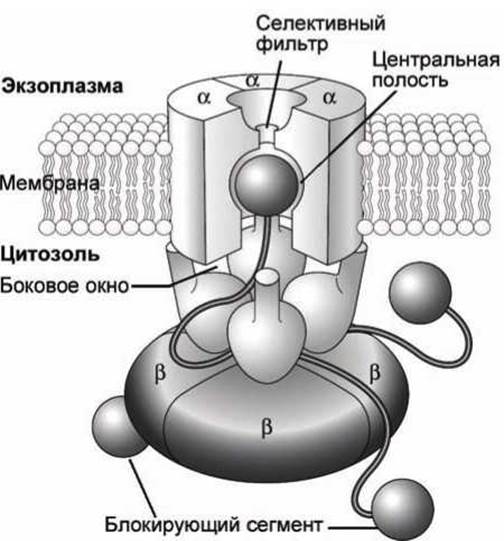
Рисунок 133 - Схема инактивации потенциалочувствительного калиевого канала
Через несколько миллисекунд после реполяризации мембраны блокирующий сегмент вытесняется из канала, и белок возвращается в исходное состояние.
Ионный канал образуется четырьмя мембранными доменами а, a цитозольные домены ß являются регуляторными. Глобулярные домены калиевого канала функционально эквивалентны блокирующему сегменту в натриевом канале.
Искусственная модификация белковых петель, соединяющих блокирующие глобулы с телом канала показали, что при укорочении соединяющей белковой нити канал инактивируется быстрее, а при удлинении этой нити инактивация канала замедляется.