Биологические мембраны - А. Н. Огурцов 2012
Электрогенез биомембран
Механизмы межклеточной сигнализации
Трансмиттеры
Для реагирования на изменение внешних условий или для согласования клеточной активности клеток в многоклеточном организме клеткам необходимо получать и передавать информацию. Для этого клетки постоянно обмениваются химическими сигналами, которые называются трансмиттерами.
Трансмиттеры классифицируют различным образом. Можно, например, различать трансмиттеры по времени их жизни в организме.
Минимальное время жизни у трансмиттеров, которые используются в щелях межклеточных синапсов шириной порядка 100 нм, составляет ~10-2-10-4с.
Гормоны представляют собой противоположный случай, когда трансмиттер существует в организме минуты и даже часы. Гормоны секретируются в кровь и разносятся по тканям организма током крови. Большинство гормонов вырабатываются специализированными железами внутренней секреции.
Паракринные трансмиттеры также имеют большое время жизни, но они выделяются не в кровь, а в специфические ткани и диффундируют в пределах данной ткани.
Другой способ классификации сигнализационных механизмов основан на местоположении и специфике рецепторов, принимающих химический сигнал. Различают три типа рецепторов для трансмиттеров:
1) ионотропные рецепторы наружной мембраны клетки;
2) метаботропные рецепторы наружной мембраны клетки;
3) внутриклеточные рецепторы.
Ионотропные рецепторы наружной мембраны клетки представляют собой ионные каналы, которые открываются в результате специфического связывания определённого трансмиттера с экзоплазматическим доменом такого канального белка. Ниже мы рассмотрим типичный пример - никотиновый ацетилхолиновый рецептор.
Механизм работы ионотропных рецепторов наружной мембраны клетки сходен с уже рассмотренными выше лиганд-управляемыми каналами: инозитол-трисфосфат-управляемым каналом (рисунок 140) и цАМФ-управляемым каналом (рисунок 145). Отличие же заключается в том, что эти два типа каналов открываются под действием цитозольных молекул, а ионотропные рецепторы наружной мембраны клетки управляются внеклеточными метаболитами.
Метаботропные рецепторы наружной мембраны клетки всегда сопряжены с клеточными ферментами. Такого типа рецепторы уже встречались выше: АДФ-рецептор активирует белок Gq, а затем фосфолипазу PLCß (рисунок 139); рецептор запаха (рисунок 145) и ß-адренергический рецептор сопряжены cG, и далее с аденилат циклазой, синтезирующей цАМФ. Тирозин-киназные рецепторы сами являются протеин-киназами, которые активируются связыванием со своими лигандами (как фактор PDFG на рисунке 147).
а -Адренергический рецептор (совершенно аналогично схеме для АДФ-рецептора (рисунок 139)) в ответ на связывание с норадреналином (рисунок 152) активирует белок Gq, а затем фосфолипазу PLCß, что приводит к росту концентрации инозитол-трисфосфата в цитозоле, открытию инозитол-трисфосфат-управляемых кальциевых каналов и росту концентрации кальция в цитозоле (рисунок 140).
Адренергические рецепторы а и ß являются подобными, но различными рецепторными белками. а-Адренергический рецептор связывается с норадреналином, а ß-адренергический рецептор - с адреналином.
Внутриклеточные рецепторы располагаются внутри клетки (в цитозоле или в нуклеоплазме) и связывают трансмиттеры, которые диффундировали через плазматическую мембрану. После связывания с трансмиттерами эти рецепторы активируют специфические ферменты. Примерами таких рецепторов являются рецептор оксида азота (NO) и рецептор стероидных гормонов.
NO является трансмиттером сигналов во многих тканях. Он не накапливается в организме, а синтезируется ферментом NO-синтаза из аргинина в том месте, где он нужен. Оксид азота легко диффундирует сквозь плазматические мембраны и связывается с цитозольными белковыми NO-рецепторами. Например, связывание с NO фермента гуанилат циклаза активизирует его, и он катализирует реакцию превращения ГТФ во внутриклеточный месенджер циклический гуанозин монофосфат, цГМФ.
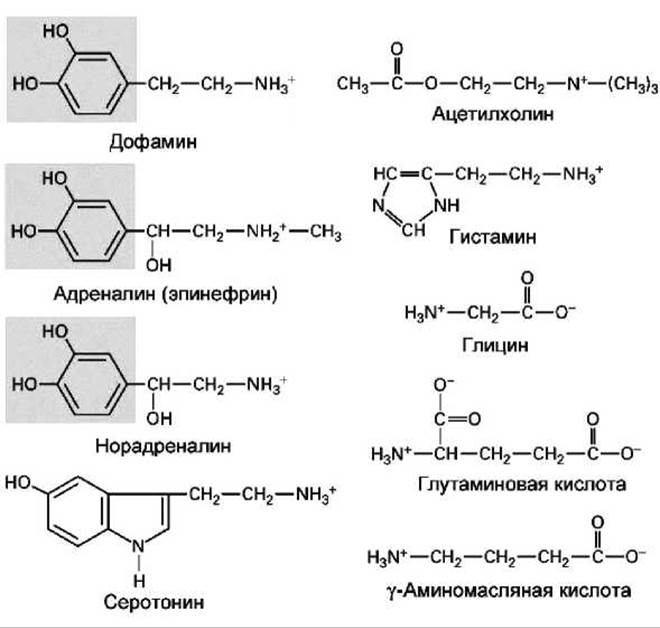
Рисунок 152 - Основные нейротрансмиттеры (нейромедиаторы)
Стероидные гормоны связываются с внутриклеточными рецепторами стероидных гормонов. Например, цитозольный рецептор глюкокортикоидных гормонов, связываясь с этими гормонами (такими, как гидрокортизон, кортикостерон и другие, синтезируемые клетками коры надпочечников), переходит в активную форму, после чего он может пройти через ядерную пору в ядро клетки, где он, связываясь с ДНК в специфических энхансерных сайтах HRE (hormone response element), стимулирует транскрипцию генов, обеспечивающих метаболизм углеводов и белков (см. [2] п. 3.5).
Нейротрансмиттеры синапсов. Передача сигналов между нейронами и от мотонейронов к клеткам мускулатуры производится в специализированных контактах, называемых синапсами, с помощью молекул-трансмиттеров, называемых нейротрансмиттерами (или нейромедиаторами).
Существует множество малых молекул, выполняющих функцию нейротрансмиттеров (рисунок 152).
За исключением ацетилхолина все остальные нейротрансмиттеры являются аминокислотами или производными аминокислот. Так дофамин, адреналин и норадреналин синтезируются из тирозина, серотонин - из триптофана, гистамин - из гистидина, у-аминомасляная кислота (y-amino-butyric acid, GABA) - из глутаминовой кислоты. Такие нуклеотиды, как АТФ, и соответствующие нуклеозиды с меньшим числом фосфатных групп, также используются в качестве нейротрансмиттеров. Каждый нейрон, как правило, синтезирует только один тип нейротрансмиттеров.
Все "классические" нейротрансмиттеры синтезируются в цитозоле нейрона, а затем импортируются в синаптические пузырьки в терминалях аксона, где они хранятся до момента использования (до прихода нервного импульса). Эти пузырьки диаметром 40-50 нм имеют повышенную кислотность люмена, которая создаётся протонными помпами V-типа, функционирующими в мембранах этих везикул.
Аналогично системам аккумулирования метаболитов в вакуолях растений (рисунок 105), созданный этими помпами градиент протонов (в люмене секреторной везикулы концентрация протонов выше, чем в цитозоле) обеспечивает импорт нейротрансмиттеров из цитозоля в люмен секреторных пузырьков с помощью Н+-сопряжённых антипортёров в мембране этих пузырьков.
Например, синаптические везикулы накапливают ацетилхолин из цитозоля с помощью Н+/ацетилхолинового антипортёра в мембране везикул. А сам ацетилхолин в цитозоле синтезируется из ацетил СоА и холина в реакции, катализируемой ацетилтрансферазой
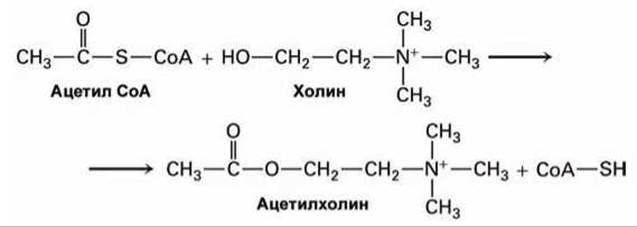
Примечательно, что ген, кодирующий белок-антипортёр, находится в первом нитроне гена, кодирующего ацетилтрансферазу. Этим самым в ходе эволюции сохранилась однозначная координация экспрессии этих двух белков. Для других нейротрансмиттеров используются другие Н+/нейротрансмиттерные антипортёры.