ОСНОВЫ БИОТЕХНОЛОГИИ МИКРОВОДОРОСЛЕЙ - Д. С. ДВОРЕЦКИЙ - 2015
1. ВОДОРОСЛИ. ОБЩИЕ СВЕДЕНИЯ
1.1. Основы классификации водорослей и филогенетические связи
Водоросли - очень многочисленная и крайне неоднородная в систематическом отношении группа организмов, рассматриваемая большинством систематиков как совокупность нескольких самостоятельных отделов (типов) [1, 2]. В прошлом водоросли считались примитивными растениями (без специализированных проводящих или сосудистых, тканей); их выделяли в подотдел водорослей (Algae), который вместе с подотделом грибов (Fungi) составлял отдел талломных (слоевцовых) или низших растений (Thallophyta), - один из четырёх отделов царства растений. В соответствии с системой, принятой альгологами, водоросли состоят из 11 отделов: Cyanophyta (сине-зелёные водоросли, цианеи), Cryptophyta (криптофиты), Rhodophyta (красные водоросли), Chrysophyta (золотистые водоросли), Bacillariophyta (диатомовые водоросли), Xanthophyta (жёлтозелёные), Dinophyta (перидинеи), Euglenophyta (эвгленовые водоросли), Phaeophyta (бурые водоросли), Chlorophyta (зелёные водоросли), Charophyta (харовые). За основу подразделения водорослей на отделы положены два общих критерия: морфологический (различия в уровне развития и степени дифференциации таллома) и физиологический (различия в пигментном составе).
Степень родства между отделами водорослей представлена на рис. 1 [2].
По типу клеточной организации водоросли делятся на прокариот (Cyanophyta), мезокариот (Dinophyta) и самая многочисленная группа эукариот (остальные отделы) [1, 2].
Эукариоты в свою очередь разделяются по цвету в зависимости от окраски хлоропластов на три ветви, соответственно включающие красные водоросли (Rhodophyta), зелёные водоросли (Euglenophyta, Chlorophyta, Charophyta) и группу отделов с преобладанием жёлто-бурых пигментов (Chrysophyta, Xanthophyta, Bacillariophyta, Phaeophyta).
Цвет - достаточно прочная, но не единственная основа для общей классификации этих организмов. Более существенны для выделения различных групп водорослей типы формирования их колоний, способы размножения, особенности хлоропластов, клеточной стенки, запасных веществ и т. п.
Следует обратить внимание на то, что все отделы водорослей, кроме Charophita и Phaeophyta, включают и одноклеточные формы, а Euglenophyta, Chrysophyta и Bacillariophytaпредставлены только этим уровнем организации таллома, причём даже у харовых и бурых водорослей в жизненном цикле сохранились одноклеточные стадии, представленные репродуктивными клетками.
Рис. 1. Происхождение и филогенетические связи между отделами водорослей
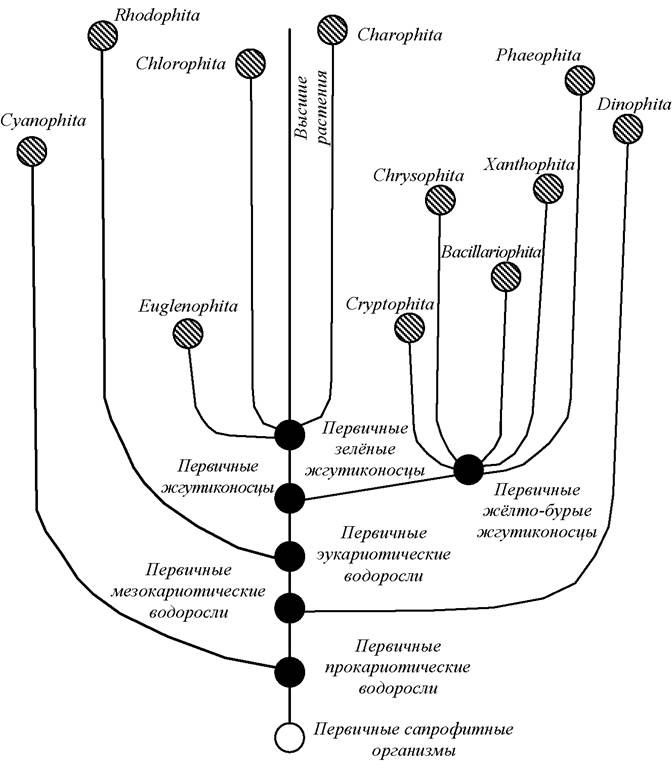
Эволюция различных отделов водорослей шла параллельно, что привело к появлению сходных ступеней морфологической дифференциации, но только какие-то предки Chlorophytaдали начало наземным хлорофиллоносным растениям. Близкой к Chlorophyta по происхождению, но рано отделившейся и специализировавшейся группой оказались эвгленовые (Euglenophyta), которые в своей эволюции не вышли за пределы монадной (одноклеточной) организации. Наоборот, харовые водоросли обособились от зелёных водорослей тогда, когда среди них появились формы, уже достаточно высокоморфологически дифференцированные.
Одноклеточные представители Chlorophyta составляют два хорошо выраженных класса - Volvocophyceae и Chlarococcophyceae. Вольвоксовые водоросли относятся к типичным монадам и лишь при наступлении неблагоприятных условий временно переходят в пальмеллевидное состояние, вызываемое сильным ослизнением клеток, хлорококковые водоросли представлены коккоидными формами, лишёнными жгутиков, а неподвижная стадия у них ограничивается коротким периодом размножения, когда образуются гаметы и зооспоры. Однако наиболее примитивные из числа хлорококковых иногда обнаруживают сходство с вольвоксовыми.
Многоклеточные растения без особого труда можно отличить от многоклеточных животных, но такого нельзя сказать относительно одноклеточных водорослей и животных. Это объясняется тем, что подвижные одноклеточные водоросли обладают признаками, которые свойственны не только растительным, но и животным организмам. Клетки всех эукариот имеют общий план строения и во многом сходный набор органелл (аппарат Гольджи, митохондрии, ядро, эндоплазматическую сеть и т.д.) с одинаковым типом организации. Растительная клетка характеризуется рядом отличительных черт [2]:
1) присутствие хлоропласта, который влияет на ход обменных процессов, приводит к выработке своеобразных в химическом отношении запасных питательных веществ полисахаридной природы и появлению нового набора ферментов;
2) возникновение наружного покрова в виде оболочек. У водорослей, наряду с целлюлозой, в их составе обнаружен пектин, фукоидан и другие полисахаридные соединения.
По этим признакам водоросли, без сомнения, должны быть причислены к растительному царству.
В то же время подвижные клетки некоторых водорослей роднит с одноклеточными животными отсутствие жёсткой оболочки и наличие таких структур, как жгутики, сократительные вакуоли, стрекательные структуры. Встречаются организмы, которые способны переходить от автотрофного к гетеротрофному типу питания, теряя при этом окраску. В состав водорослей входит некоторое число форм бесцветных жгутиконосцев, произошедших от окрашенных благодаря утрате хлорофилла (Astasia, Perantma).
Современная распространённая научная точка зрения рассматривает одноклеточные водоросли в качестве неорганизованных растительных организмов (Protophyta), а одноклеточных, соответственно, как наиболее примитивных представителей животного мира (Protozoa), не вкладывая в эти понятия конкретного таксономического смысла, а лишь подчёркивая сходный уровень в развитии биологических систем в двух разных царствах живой природы, рано разделив развивавшихся параллельно.
Зелёные водоросли обычно цвета зелёной травы (хотя окраска может варьировать от бледно-жёлтой до почти чёрной), а фотосинтетические пигменты у них такие же, как у обычных растений. Большинство зелёных водорослей - микроскопические пресноводные формы, многие виды растут на почве, образуя на её влажной поверхности напоминающие войлок налёты. Насчитывается несколько тысяч видов зелёных водорослей. Встречаются представители зелёных водорослей одно- и многоклеточные, могут образовывать нити, шаровидные колонии, листовидные структуры и т. д.
Клетки зелёных водорослей подвижные (с двумя жгутиками) или неподвижные. Половое размножение - разных уровней сложности в зависимости от вида. Клетки содержат ядро и несколько чётко оформленных хлоропластов.
Один из хорошо известных родов - плеврококк (Pleurococcus), одноклеточная водоросль, образующая зелёные налёты, часто наблюдаемые на коре деревьев. Широко распространён род Spirogyra - нитчатые водоросли, образующие длинные волокна тины в ручьях и холодных речках. Весной они плавают в виде липких желтовато-зелёных скоплений на поверхности прудов. Cladophora растёт в виде мягких, сильно ветвящихся «кустиков», прикрепляющихся к камням у берегов рек. Basiocladia образует зелёный налёт на спине пресноводных черепах. Состоящая из многих клеток водяная сеточка (Hydrodictyon), обитающая в стоячих водах, по строению, действительно, напоминает «авоську». У некоторых видов клетки соединены в нитчатые колонии. У свободноплавающей колониальной водоросли Scenedesmus серповидные или продолговатые клетки объединены в короткие цепочки. Этот род обычен в аквариумах, где его массовое размножение приводит к появлению в воде зелёного «тумана». Самая крупная зелёная водоросль - морской салат (Ulvа), макрофит листовидной формы.
Красные водоросли (багрянки) - морские листовидные, кустистые или корковые макрофиты, обитающие ниже линии отлива. Цвет багрянок преимущественно красный из-за присутствия пигмента фикоэритрина, но может быть пурпурным или синеватым. Некоторые багрянки встречаются в пресной воде, главным образом, в ручьях и прозрачных быстрых речках. Batrachospermum - студенистая на ощупь, ветвистая водоросль, состоящая из буроватых или красноватых, похожих на бусины клеток. Lemanea - щётковидная форма, часто растущая в быстро текущих речках и водопадах, где её талломы прикрепляются к камням. Audouinella - нитчатая водоросль, встречающаяся в мелких речках. Ирландский мох (Chondrus сripus) - обычный морской макрофит. Багрянки не образуют подвижных клеток. Их половой процесс очень сложен, и один жизненный цикл включает несколько фаз.
Бурые водоросли - обитатели моря. Лишь немногие виды микроскопические, а среди макрофитов встречаются самые крупные водоросли в мире. К последней группе относятся ламинарии, макроцистисы, фукусы, саргассумы и лессонии («морские пальмы»), наиболее обильные по побережьям холодных морей. Все бурые водоросли многоклеточные. Цвет их варьирует от зеленовато-жёлтого до тёмно-коричневого и обусловлен пигментом фукоксантином. Половое размножение связано с образованием подвижных гамет с двумя боковыми жгутиками. Экземпляры, образующие гаметы, часто совершенно не похожи на организмы того же вида, размножающиеся только спорами.
Диатомовые водоросли (диатомеи) объединяют в класс Bacillariophyceae, который в используемой здесь классификации входит вместе с золотистыми и жёлто-зелёными водорослями в отдел (тип) Chrysophyta. Диатомеи - весьма обширная группа одноклеточных морских и пресноводных видов. Окраска клеток диатомей варьируется от жёлтой до бурой, из-за присутствия пигмента фукоксантина. Протопласт диатомей защищён коробчатой кремнеземной (стеклянной) оболочкой - панцирем, состоящим из двух створок. Твёрдая поверхность створок часто покрыта характерным для вида сложным узором из штрихов, бугорков, ямок и гребней. Эти панцири - одни из самых красивых микроскопических объектов, а чёткость различения их узора используется иногда для проверки разрешающей силы микроскопа. Обычно створки пронизаны порами или имеют щель, называемую швом. В клетке находится ядро. Помимо деления клеток надвое известно и половое размножение. Многие диатомеи являются свободноплавающими формами, но некоторые прикреплены к подводным объектам слизистыми ножками. Иногда клетки объединяются в нити, цепочки или колонии. Различают два типа диатомей: перистые, с удлинёнными двустороннесимметричными клетками (они наиболее обильны в пресных водах) и центрические, клетки которых, если смотреть со створки, выглядят округлыми или многоугольными (их больше всего в морях). Панцири диатомей сохраняются после смерти клеток и оседают на дно водоёмов, с течением времени образуя пористую горную породу - диатомит [1, 2].
1.2. Основы микроводорослей. Особенности строения клеток микроводорослей
Клетки микроводорослей по строению похожи на клетки растений, однако у них имеются специфические особенности, обусловленные специфичностью среды обитания и обмена веществ [1 - 4]. Уникальными особенностями клеток микроводорослей обладают клеточные покровы, которые представляют собой многослойные, сложно организованные структуры [2, 5], и включают структуры: плазмалемму, перипласт, клеточную оболочку и др.
Плазмалемма. У водорослей содержимое клетки (протопласт) снаружи ограничивается тонким, порядка 60... 100 А белково-липидным слоем, обладающим специфическими свойствами, который сам по себе не способен фиксировать форму клетки и который получил название плазматической мембраны, или плазмалеммы.
На поперечных срезах плазмалемма представляется гладким образованием, обнаруживает трёхслойную организацию. На внутренней, обращённой к цитоплазме поверхности плазмалеммы, удалось выявить многочисленные глобулярные частицы и углубления, или складки, которые имеют белковую природу и выполняют транспортную функцию.
Перипласт у водорослей представляет собой плотное многослойное образование, состоящее преимущественно из белковых соединений, разрушающихся под действием пепсина и трипсина, причём один слой располагается над плазмалеммой, а другой под ней, и по толщине (порядка 75 А) они между собой не отличаются. Перипласт, толщина которого равна приблизительно 250 A, состоит из серии полос, направляющихся прямыми, продольными рядами от переднего к заднему концу клетки, где они обычно значительно сужаются. В областях контакта полос друг с другом образуются глубокие борозды. Чередование гребней и борозд придаёт клетке рельефный контур. В местах расположения гребней и борозд области перипласта отличаются по своей прочности, и более слабые из них обеспечивают клетке гибкость и позволяют в какой-то мере менять форму. Перипласт пронизан порами, через которые клетка сообщается с внешней средой.
Клеточная оболочка, состоящая из целлюлозы, наиболее удачно сочетает защитную и опорную функции с ростовыми процессами и проницаемостью, поэтому такого рода покров становится господствующим у большинства растений.
Толщина оболочки сильно варьирует в зависимости от видовой принадлежности организма, его возраста, функционального состояния и условий окружающей среды. Покоящиеся клетки имеют более толстые оболочки по сравнению с репродуктивными и молодыми вегетативными клетками. При дефиците влаги в среде обитания оболочки значительно утолщаются.
У подавляющего большинства водорослей оболочки сплошные, но встречаются и составные - из двух или более частей, как у некоторых диатомовых, жёлто-зелёных и зелёных.
На долю клеточной оболочки в среднем приходится 13.15% сухого веса всей клетки, 7,2% от этого количества составляют жиры, а остальное - белки и углеводы. В клеточной оболочке у Chlorella жиров содержится около 9%, а у Anacystis nidulans до 36%, однако основой клеточной оболочки служит белково-углеводный комплекс.
Клеточная оболочка представляет собой многослойную структуру. Слои отличаются друг от друга по толщине, плотности, химическому составу, - внутренние слои обычно целлюлозные, а наружный - пектиновый. У некоторых водорослей, таких, например, как Scenedesmus, мощно развитый пектиновый слой, помимо защитной функции, способствующий объединению клеток в колонии.
У подавляющего большинства водорослей оболочка оказалась двухкомпонентной, состоящей из аморфного основного вещества (матрикса) и погружённых в него фибрилл целлюлозной природы с сечением 200...300 А, которые выполняют роль опорного элемента и повышают прочность всей конструкции.
Иногда клеточные оболочки интенсивно пропитываются (инкрустируются) солями железа (у вольвоксовых), кальция (у харовых) или кремния (у диатомовых), которые укрепляют матрикс и нередко создают структуру в виде панциря. Оболочки могут пропитываться не только минеральными солями, но и органическими соединениями, из которых самыми распространёнными оказались лигнин и кутин. Последний, помимо опорной, выполняет ещё и защитную функцию, поскольку он задерживает ультрафиолетовые лучи.
Оболочки некоторых видов водорослей способны выделять и накапливать вещества на поверхности клетки, образуя дополнительный твёрдый покров - кутикулу. Этот процесс называется адкрустацией. Подобного рода структуру имеют вегетативные клетки Porphyra, Oedogonium, эпидермальные клетки бурых водорослей. У красных водорослей на поверхности оболочек адкрустируются содержащие сульфатные группы полисахариды. Их пополнение идёт непрерывно, поэтому они скапливаются в таком большом количестве, что образуют вокруг клетки мощную капсулу в виде слоистого чехла, состоящего из аморфного вещества. Водоросли, вырабатывающие подобного рода полисахариды, являются ценным сырьём для получения агар-агара.
У многих водорослей, особенно из числа одноклеточных, на поверхности оболочки появляются различного рода дополнительные структуры - щетинки, шипики, чешуйки. Роль этих структур заключается в выполнении защитной функции или поддержании клетки в состоянии парения.
Поскольку микроводоросли представляют собой эукариотические организмы, строение и структура органелл клеток является сложной. Как и большинству клеток животных, растений, клетки микроводорослей содержат структуры, общие для этих царств - ядро, митохондрии, аппарат Гольджи, эндоплазматическая сеть и др. [2].
Ядро и аналоги. У прокариотических сине-зелёных водорослей данная структура представлена нуклеоидом, у мезокариотических динофлагеллат - слабо дифференцированными хромосомами, у эукариотических водорослей - структурно оформленными хромосомами [2, 5].
Установлено, что в клетках сине-зелёных водорослей отсутствует морфологически выраженное ядро, с помощью электронного микроскопа выявлен хорошо дифференцированный пучок микрофибрилл, соответствующий одной кольцевой молекуле ДНК.
В клетках динофлагеллат уже присутствует морфологически оформленное ядро с оболочкой, ядрышком и хромосомами. Однако хромосомы, подобно нуклеоиду, лишены гистонов и состоят из микрофибрилл толщиной приблизительно 50 А, содержащих чистую ДНК. Хромосомы динофлагеллат слабо дифференцированы и постоянно находятся в конденсированном состоянии.
У всех остальных водорослей хромосомы, состоящие из ДНК, связанной с белком, имеют размеры порядка 125 А. Ядра всех эукариотических водорослей в интерфазе окружены чётко выраженной оболочкой и содержат нуклеоплазму, ядрышко, хроматин.
Большинство водорослей характеризуется крайне мелкими размерами ядер, размер которых, как правило, коррелирует с размером клетки. Форма ядер у водорослей очень разнообразна, но чаще встречается шаровидная или чечевицеобразная. Этот признак весьма изменчив и находится в зависимости от таких факторов, как фаза жизненного цикла организма или местоположение в клетке (центральное или периферическое).
Ядерная оболочка (нуклеолемма) представляет двухмембранную систему, мембраны которой отстоят друг от друга на расстоянии порядка 100...300 А. Местами оно может значительно расширяться и частично переходить в систему каналов эндоплазматической сети, причём наружная мембрана оболочки несёт на поверхности, обращённой к цитоплазме, многочисленные рибосомы. Оболочка ядра обычно пронизывается порами диаметром от 300 до 850 А.
Всё пространство ядра заполняется нуклеоплазмой, в которой располагается от одного до нескольких ядрышек. Они лишены пограничной мембраны и имеют вид плотных образований, чаще всего, круглой формы. Число, форма и размеры ядрышек могут меняться на протяжении жизненного цикла клетки. Местоположение ядрышка в ядре строго не фиксировано, поэтому оно различно даже на разных стадиях развития одного и того же организма.
Во время деления ядра ядрышко обычно претерпевает сильные изменения, при этом затрагиваются не только его размеры и форма, но и внутренняя организация. Преобразования в основном приурочены к двум периодам - профазе (профазный цикл перестроек) и телофазе (телофазный цикл перестроек).
Помимо ядрышек, в нуклеоплазме располагаются также хроматиновые образования в виде скрученных нитей и глыбок, называемых иногда хромоцентрами, или кариосомами.Основная масса хроматина обычно скапливается вдоль оболочки, поэтому остальная часть ядра выглядит почти гомогенной.
Еще в конце прошлого столетия в цитоплазме как животных, так и растительных клеток были обнаружены множественные, очень своеобразные органеллы, получившие название диктиосом. Вся их совокупность в клетке стала обозначаться как аппарат, или комплекс Гольджи. Число и размер диктиосом у разных видов водорослей сильно колеблется, чаще всего, они располагаются вокруг ядра.
Диктиосомы в клетках водорослей представляют собой систему собранных в стопки, коротких, уплощенных мешочков - цистерн с отделяющимися по бокам многочисленными мелкими пузырьками, которые по мере удаления от органеллы увеличиваются в размерах.
Аппарат Гольджи принимает участие в накоплении, транспорте и секреции полисахаридов для клеточных оболочек, в образовании некоторых структур клеточных оболочек. Установлена связь аппарата Гольджи с секреторной деятельностью клетки и образованием лизосом. В отпочковывающихся от диктиосом пузырьках происходит наряду с концентрацией и уплотнение продуктов секреции, которые затем транспортируются в разные участки клетки.
Митохондрии - сравнительно крупные, часто овальной формы тельца. Они отсутствуют лишь у прокариот. Число и размер митохондрий может значительно варьировать в зависимости от возраста и функционального состояния клетки. Расположение митохондрий в клетках строго не фиксировано. Обычно они располагаются более или менее равномерно по всей клетке, митохондрии являются источниками энергии в клетке, её дыхательными центрами [2, 5, 6].
Ультраструктура митохондрий у всех живых организмов однотипна. Митохондрия окружена двухмембранной оболочкой: наружная мембрана гладкая, сплошная, внутренняя - образует выросты, или кристы, обращённые в гомогенный матрикс, содержащий отличные от ядерных ДНК и рибосомы. Нередко интенсивное развитие крист компенсирует небольшие размеры митохондрий или их малочисленность в клетке.
Во время ядерного деления происходит увеличение числа митохондрий в клетке. Выделено, по крайней мере, два механизма обеспечения дочерних клеток митохондриями: синтез и деление материнской митохондрии.
Цитоплазма клетки в большей или меньшей степени пронизана системой канальцев - эндоплазматической сетью, которые местами расширяются, образуя то цистерны, то пузырьки. Мембраны эндоплазматической сети представлены двумя формами - гладкой и шероховатой, несущей рибосомы.
Эндоплазматическая сеть участвует в перемещении и распределении различных веществ. Как правило, скопление её каналов происходит в тех областях, где интенсивно идут формообразовательные процессы (в местах роста плазмалеммы и заложения клеточных оболочек, в зоне расположения делящихся органелл и т.д.) и куда усиливается приток веществ. Кроме того, в каналах происходит концентрация, а возможно, и синтез, главным образом, белковых соединений, в отличие от цистерн диктиосом аппарата Гольджи, в которых происходит только скопление углеводов. Эндоплазматическая сеть наряду с аппаратом Гольджи принимает участие в образовании клеточных оболочек и других поверхностных структур. Ещё одной функцией эндоплазматической сети является объединение всех компонентов клетки в единое целое [2, 5, 6].
При формировании вакуолярной системы клеток микроводорослей активное участие принимает не только аппарат Гольджи, но и эндоплазматическая сеть, причём с возрастом вакуолярная система становится интенсивно развитой: вакуоли увеличиваются как в числе, так и в размере. Последнее происходит за счёт слияния мелких пузырьков в более крупные. В старых клетках нередко можно наблюдать одну крупную, центральную вакуоль [2, 5, 6].
В растительных клетках, в том числе микроводорослей, помимо эндоплазматической сети обнаруживаются прямолинейные, гладкоконтурные трубчатые элементы, или микротрубочки, которые особенно чётко видны, когда собираются в параллельные пучки. В сечении микротрубочки имеют вид цилиндров с диаметром просвета 200...250 А. Микротрубочки, в свою очередь, состоят из упорядоченной системы более мелких трубчатых элементов сечением порядка 60 А в числе от 8 до 14.
Микротрубочки могут то появляться в клетке, то исчезать, перемещаться из одной её части в другую, увеличиваться или уменьшаться в числе. Например, у Chlorella микротрубочки возникают перед самым делением ядра и исчезают сразу же после завершения цитокинеза [2].
Функционально микротрубочки, наряду с каналами эндоплазматической сети, участвуют в транспортировке веществ в клетке в места, где протекают интенсивные формообразовательные процессы, например, в зонах заложения клеточных перегородок, они принимают участие в образовании веретена деления.
В цитоплазме встречаются различной формы и размеров цитоплазматические тела, окружённые одиночной мембраной и заполненные содержимым, отличающимся по своей электронной плотности.
Среди цитоплазматических частиц выделяется особая группа структур - лизосом. Лизосомы отличаются высоким содержанием ферментов- гидролаз, способных расщеплять важнейшие классы химических соединений, включая белки, углеводы и нуклеиновые кислоты. Содержащиеся в лизосомах ферменты разобщены мембранным барьером с клеточным субстратом, поэтому они расщепляют только те вещества, которые в них поступают через систему транспортных пузырьков. Если целостность лизосомных мембран нарушается, то ферменты высвобождаются непосредственно в клетку и вызывают растворение внутриклеточных мембран, разрушение органелл и даже целых участков протопласта.
Пероксисомы характеризуются высоким содержанием целого ряда других ферментов пероксидазного комплекса. Наличие окислительных ферментов говорит об участии пероксисом в процессах фотодыхания. Образование пероксисом самым тесным образом связано с деятельностью эндоплазматической сети. Пероксисомы в клетках водорослей имеют палочковидную, дисковидную или линзовидную форму, достигают размеров 0,4...1,8 мкм и состоят из мелкозернистого материала.
Сферосомы - сравнительно крупные (0,5...1,0 мкм), сильно преломляющие свет частицы, встречаются только в цитоплазме растительных клеток и часто используются ботаниками для изучения её токов. В составе сферосом обнаружены скопления гидролитических ферментов, липидов, ароматических аминокислот типа тирозина. Основная функция сферосом состоит в синтезировании липидов. В образовании сферосом принимает участие эндоплазматическая сеть.
Клетки микроводорослей, как и любые живые клетки, содержат включения немембранной природы, которые представляют собой запасные питательные вещества.
Скопления полисахаридов запасаются в виде крахмальных зёрен, которые у Chlorophyta и Charophyta располагаются, как и у высших растений, исключительно в хлоропластах, а у водорослей из других отделов - в цитоплазме. У Euglenophyta вырабатывается особый тип полисахарида - парамилон и концентрируется преимущественно вокруг пиреноида, у Phaeophyta выявлен водорастворимый полисахарид - ламинарин, у Rhodophyta - багрянковый крахмал, а у Chrysophyta - капли жидкого лейкозина.
Запасные вещества в виде капель липидов обнаружены в клетках многих водорослей, преимущественно в тот период, когда они находятся в покоящемся состоянии, но особенно богаты липидами клетки динофлагеллат и диатомовых водорослей. Липиды скапливаются в виде разного размера плотных глобул.
У многих водорослей в цитоплазме присутствуют сферические плотные структуры, содержащие полифосфаты.
Микроводорослям как фотосинтетическим организмам присущи некоторые специфические растительные клеточные образования. В первую очередь, следует отметить фотосинтетический аппарат. Его возникновение обусловило переход организмов на автотрофный способ питания, вызвало изменения в обменных процессах, привело к появлению сложных ферментативных систем и особых типов запасных веществ. Основная функция фотосинтетического аппарата - поглощение энергии солнечного света и превращение её в энергию химических связей, что возможно только в присутствии хлорофилла. В фотосинтетическом аппарате, помимо хлорофилла, сосредоточиваются в большом количестве и другие пигменты, от количественного соотношения которых зависит окраска клеток [2, 3, 5].
Водоросли, очевидно, были именно той группой растительного царства, в пределах которой впервые возник и закрепился аппарат, выполняющий функции фотосинтеза.
У эукариотических водорослей фотосинтетический аппарат представлен структурно оформленной органеллой - хлоропластом.
Хлоропласты водорослей отличаются крайним разнообразием своей морфологии. Здесь встречаются организмы, в клетках которых присутствует всего один очень крупный хлоропласт, и наряду с этим известно много случаев, когда содержится два и больше хлоропластов. Очень разнообразна форма хлоропластов. По положению в клетке различают постенные и осевые хлоропласты.
Структурно хлоропласт окружён оболочкой, состоящей из двух одиночных мембран. Хлоропласт - гетерогенная система, за счёт того, что в его строме располагаются более плотные зоны ДНК, диски, микротрубочки, рибосомы, фотосинтетические продукты (крахмал или другие полисахаридные соединения), метаболические гранулы.
Фотосинтетический аппарат водорослей крайне неоднороден по пигментному составу. Зелёная окраска обусловлена присутствием хлорофилла, представленного несколькими формами - a, b, c и d. Значительная доля приходится на каротиноиды и фикобилины (фикоциан и фикоэритрин) [2, 5, 6]. Они нередко маскируют основную зелёную окраску, вместо которой появляется сине-зелёная, бурая, красная и т. д.
Помимо дисков, в хлоропластах присутствуют также микротрубочки, которые обычно собираются в пучки и располагаются, как правило, вдоль оболочки органеллы, эти структуры играют роль скелетных образований.
В строме хлоропластов водорослей всех отделов обнаружены рибосомы; у водорослей из отделов Chlorophyta и Charophyta в хлоропластах иногда скапливается значительное количество крахмальных зёрен.
Хлоропласт относится к числу преемственных органелл и обладает способностью к самовоспроизведению. Хлоропласты делятся перетяжкой или почкованием.
Пиреноид (от греч. пирен - косточка, эйдос - вид) - специфическая структура водорослевых хлоропластов. Пиреноид представляет собой плотное образование белковой природы, где помимо ферментов, обнаружены следы ДНК, РНК и хлорофилла. Снаружи пиреноид обычно окружён сильно преломляющей свет обкладкой, которая преимущественно состоит из крахмала. У бурых водорослей в её образовании принимает участие водорастворимый полисахарид - ламинарин. У красных водорослей вырабатывается особая форма крахмала - багрянкового; у эвгленовых водорослей отмечено скопление парамилона; у хризомонад откладываются липидные капли [2].
У подавляющего большинства водорослей пиреноиды находятся внутри хлоропластов, поэтому их нередко называют погружёнными.
Пиреноиды способны к самовоспроизведению, которое возможно только по достижении ими определённых размеров и осуществляется несколькими путями: делением надвое и фрагментацией. Деление начинается сразу после завершения митоза и по времени совпадает с делением пластиды и клетки. Фрагментация начинается с распадения пиреноида на многочисленные, часто неравные сегменты, рассеивания хлоропласта, достройки и вступления в фазу роста. В спорах и гаметах пиреноиды способны возникать заново.
Основная функция пиреноида заключается в синтезе различного рода соединений и, в первую очередь, крахмала.
Стигма (в переводе с греческого - метка, пятно) присуща, как правило, подвижным клеткам водорослей. Она функционально взаимосвязана не только с хлоропластом, но ещё и со жгутиковым аппаратом клетки. В прижизненном состоянии стигма представляет собой скопление красного или оранжевого пигмента и имеет сферическую, палочковидную, линзовидную или трапециевидную форму, а после фиксации приобретает вид плотного, интенсивно окрашивающегося гомогенного тела, размеры которого даже в пределах видов одного рода могут заметно колебаться. Стигма является фоторецепторным органом [2].
Основу стигмы составляют пигментонесущие, тесно прилегающие друг к другу плотные глобулы разного диаметра. Линзовидное тело занимает почти 2/3 стигмы и состоит из группы строго упорядоченных уплощённых пузырьков, из которых наружные - пустые, а внутренние - заполнены зернистым содержимым. В основании линзы размещается ретиноид-чашеподобное образование, содержащее зоны определённым образом ориентированных фибрилл и зернистого вещества, а также расположенные на самом дне в один ряд глобулы диаметром около 3000 А. Такая организация стигмы способствует тому, что свет, попадая на линзу, фокусируется и затем концентрированным пучком посылается через ретиноид на глобулы, т. е. приходит туда в преобразованном виде.
Почти во всех отделах существуют водоросли (исключение составляют сине-зелёные и красные), в цикле развития которых могут образовываться подвижные клетки, характерные для животных организмов. В ходе филогенеза это свойство постепенно утрачивается на пути развития к наземным зелёным растениям [2, 5].
В пределах Chlorophyta у низкоорганизованных одноклеточных представителей из числа Volvocophyceae организмы сохраняют подвижность в течение всей своей жизни (монадные формы) и лишь при наступлении неблагоприятных условий отмечается переход к пальмеллевидному состоянию. У ряда классов водорослей подвижные клетки образуются в определённый период онтогенеза, как правило, это гаметы и зооспоры, обладающие специфичностью. У таких водорослей, как Chlorella и подобных ей представителей автоспоровой группы хлорококковых водорослей подвижная стадия в жизненном цикле полностью выпадает.
У многоклеточных зелёных водорослей подвижная стадия обычно ограничивается коротким периодом существования репродуктивных клеток и длится не более часа.
Подвижные клетки отличаются от неподвижных прежде всего появлением у них особого двигательного аппарата. Двигательный аппарат клетки - это сложная система, состоящая из трёх различающихся по организации, но тесно связанных друг с другом как структурно, так и функционально частей: жгутиков, базальных тел и ризопластов.
Движение клеток осуществляется при участии особых структур, представляющих либо временные (ризоподии), либо постоянные (реснички, псевдоцилии, жгутики) выросты клетки. Ресничками обычно называют многочисленные (от нескольких десятков до нескольких сотен и даже тысяч) короткие (порядка 5... 10 мкм) образования. Псевдоцилии иногда называют ложными ресничками, хотя они не столь многочисленны и по виду больше напоминают длинные, тонкие, чрезвычайно нежные волоски. Под жгутиками подразумевают длинные (порядка нескольких десятков микронов), как и реснички, толстые (около 0,2 мкм), но немногочисленные выросты. Жгутики отличаются от ресничек и меньшей частотой биений.
У некоторых водорослей из отдела Chrysophyta, помимо жгутиков, клетки несут ещё и неподвижный, вперёд направленный вырост - гаптонему, с помощью которого они нередко прикрепляются к субстрату. Гаптонема обычно плотнее, толще и короче жгутиков.
Среди водорослей чаще всего встречаются формы одножгутиковые, двухжгутиковые, реже трёхжгутиковые (Trichloris), четырёхжгутиковые (Carteria, зооспоры Urospora, Ulothrix, Stigeoclonium), даже восьмижгутиковые (Schizomeris), а иногда и с несколькими десятками жгутиков (в зооспорах у Oedogonium, Derbesia, Vaucheria). По соотношению длины жгутиков в пределах одной клетки водоросли разделяют на две группы: равножгутиковые и разножгутиковые.
Обычно жгутики направляются вперёд по ходу движения клетки и у равножгутиковых форм работают одинаково энергично. У разножгутиковых нередко наблюдается разница в частоте и интенсивности биений (гетеродинамизм), а также положении по отношению к клетке: как правило, длинный жгутик направлен вперёд и работает энергичнее, чем короткий, который обращён в сторону или назад по ходу движения организма. Считается, что длинный жгутик обеспечивает поступательное движение клетки и поэтому назван двигательным, а короткий - корректирует её направление и считается рулевым.
Тело жгутика цилиндрическое почти на всём своём протяжении, суживается в большей или меньшей степени лишь на самых концах. Оно окружено снаружи тонкой, порядка 90 А, мембраной, представляющей продолжение плазмалеммы, и заполнено аналогичным цитоплазме веществом, в которое погружена система из 11 пар правильно ориентированных микротрубочек с диаметром 200... 360 А каждая. Ближе к вершине жгутика число микротрубочек в нём нередко редуцируется и сохраняется только центральная. Проксимальный конец жгутика заканчивается перегородкой, к которой прикрепляется центральная пара.
В основании каждого жгутика лежит базальное тело. Оно располагается в цитоплазме вблизи поверхности клетки. Базальное тело представляет собой короткий (до 2 мкм) цилиндр, в периферической части которого располагается 9 триплетов микротрубочек, составляющих с микротрубочками жгутика единую систему, а центральная пара здесь отсутствует.
Очень сходную с базальными телами организацию имеют центриоли и среди фотосинтезирующих организмов встречаются только у водорослей. Они представляют собой парные цилиндрические тела длиной до 175 нм и сечением около 200 нм, расположенные под прямым углом друг к другу и содержащие систему из 9 триплетных периферических микротрубочек, чем очень напоминают внутреннее устройство базальных тел жгутиков. Центриоли, как правило, располагаются около ядра (внеядерные центриоли), и лишь в очень редких случаях их обнаруживают в ядре (внутриядерные центриоли). Во время митоза наблюдается расхождение центриолей в противоположные стороны. Здесь они образуют своеобразные полюса ядра, к которым обычно сходятся нити веретена деления, к которым крепятся центромеры хромосом.
Репликация центриолей приурочена к поздней телофазе или даже к интерфазе ядерного деления.
1.3. Закономерности роста и развития микроводорослей
1.3.1. Способы клеточного деления у водорослей
Одним из основных свойств организма следует признать его способность к росту и размножению путём деления. Клеточное деление (цитокинез) сопровождается сложными преобразованиями всех систем клетки. В результате её деления возникают либо только две дочерних клетки, либо их число достигает нескольких десятков и даже сотен, причём в одних случаях они оказываются обособленными друг от друга, давая начало одноклеточным организмам, а в других - объединяются в более или менее прочные, долго существующие многоклеточные комплексы (колонии, многоклеточные талломы) [2, 4].
Цитокинез самым тесным образом связан с митозом. В зависимости от степени их согласованности во времени различают последовательное, или сукцессивное деление, при котором за делением ядра сразу же следует образование клеточной перегородки (рис. 2, а), и одновременное, или симультанное деление, характеризующееся тем, что образование клеточных перегородок наступает одновременно и лишь после того, как завершится вся серия ядерных делений в клетке (рис. 2, б).
Рис. 2. Способы клеточного деления у водорослей: а - последовательное (сукцессивное) деление; б - одновременное (симультантное) деление
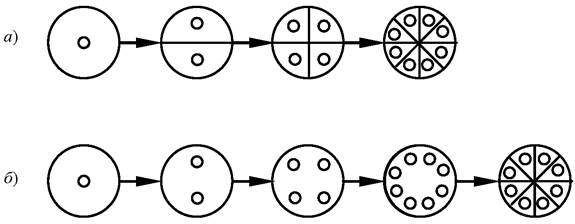
Цитокинез относится к числу наиболее продолжительных клеточных процессов и иногда растягивается на несколько часов. На скорость цитокинеза влияют такие факторы, как продолжительность и интенсивность освещения, температура, состав питательной среды и др. При выращивании водорослей в лабораторных условиях задержку клеточного деления можно вызвать за счёт прекращения обновления питательной среды, причём в данном случае влияет не столько недостаток минеральных солей, сколько накопление продуктов выделения. Проблемы клеточного деления очень актуальны в связи с решением задачи повышения продуктивности водорослей при искусственном разведении, поэтому они имеют важное практическое значение.
Деление клеток осуществляется несколькими разными способами. Многие одноклеточные как подвижные, так и неподвижные водоросли, а также вегетативные клетки в талломах многоклеточных водорослей размножаются путём деления клетки надвое, которое может повторяться, почти не прерываясь периодом роста (сукцессивное деление клеток). Наряду с этим для водорослей характерен множественный, одновременный распад содержимого клетки на отдельные участки, которые впоследствии превращаются в дочерние клетки, как это часто происходит при гамето- и спорообразовании (симультанное деление клеток).
Деление находящихся в дифференцированном состоянии клеток перетяжкой надвое у монадных форм водорослей относится к самому простому типу. У вольвоксовых, эвгленовых, хризомонад, криптомонад и динофлагеллат с гетерополярной формой клеток деление надвое происходит только в одной плоскости - в направлении длинной оси. В этот период одни клетки сохраняют подвижность, а другие - временно останавливаются.
Деление перетяжкой начинается с врастания плазмалеммы внутрь клетки и происходит либо на обоих концах одновременно, но чаще вначале затрагивает только передний конец, где располагаются жгутики. Обычно впячивающиеся (инвагинирующие) участки плазмалеммы в виде кольцевой борозды направляются навстречу друг другу и, смыкаясь, рассекают клетку на две части. Иногда деление до конца не завершается, и тогда возникает колония простейшего типа - плазмодий.
У клеток сферической формы, имеющих неопределённо большое число осей симметрии, деление на две половины происходит во всевозможных направлениях [2, 4, 5].
1.3.2. Закономерности роста и развития микроорганизмов
Рост и развитие микроводорослей полностью подчиняется основным закономерностям роста и развития всех живых организмов, поэтому при культивировании микроводорослей можно наблюдать те же фазы, что и при культивировании других микроорганизмов. Состояние процесса культивирования во времени характеризуется следующими основными параметрами: концентрация биомассы микроорганизмов X; концентрация питательной среды - субстрата (или его основного компонента) S; концентрация продукта Р.
Концентрация биомассы в процессе периодического культивирования изменяется по кривой, представленной на рис. 3 [7].
Рис. 3. Фазы культивирования микроорганизмов в полулогарифмической системе координат
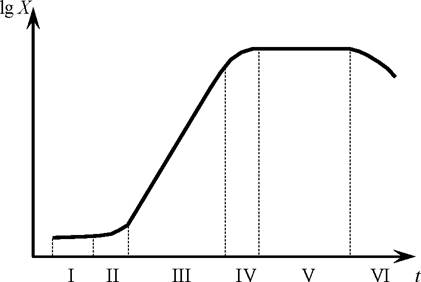
В начале культивирования (фаза I) некоторое время микроорганизмы как бы приспосабливаются к новой среде, их концентрация не меняется. Этот период называется лаг-фаза (II). Далее начинается рост - это фаза ускорения роста (III). Третья фаза - фаза наиболее интенсивного роста, происходит наибольший относительный прирост биомассы. Это фаза экспоненциального роста (IV). Затем скорость роста (относительная) начинает уменьшаться - это фаза замедления роста (V). Достигнув некоторой максимальной величины, концентрация биомассы далее перестаёт возрастать. В этой фазе - стационарной - в среде истощаются питательные вещества и накапливаются продукты обмена, тормозящие рост. Биомасса растёт и одновременно происходит гибель части клеток (автолиз), так что общая концентрация сохраняется постоянной. И наконец, в фазе отмирания автолиз начинает преобладать над ростом, и концентрация биомассы микроорганизмов снижается.
Процесс периодического культивирования характеризуется следующими кинетическими показателями:
- кинетические показатели роста биомассы (общая и удельная скорости роста);
- кинетические показатели потребления субстрата (общая и удельная скорости потребления субстрата);
- кинетические показатели биосинтеза продуктов метаболизма общая и удельная скорость биосинтеза продукта).
Важным показателем роста биомассы при периодическом культивировании является скорость роста биомассы. Для описания скорости роста используется такая характеристика, как общая скорость роста или продуктивность по биомассе Qx (г/л • ч):
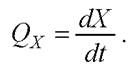
Этот показатель не вполне отражает физиологическое состояние биомассы в процессе его роста, поскольку в зависимости от концентрации биомассы в среде при одинаковом абсолютном приросте относительный прирост биомассы может отличаться в разы. В связи с этим больший интерес для характеристики интенсивности роста представляет не величина Qx, а удельная скорость роста в пересчёте на единицу биомассы μ, ч-1:
![]()
В экспоненциальной фазе, когда рост ничем не лимитирован, величина μ постоянна, а рост биомассы описывается уравнением
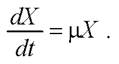
По аналогии со скоростью роста биомассы можно ввести кинетическую характеристику - общую скорость потребления субстрата QS (г/л • ч):
![]()
Знак «минус» обозначает, что скорость потребления положительна, когда концентрация субстрата в среде падает.
Аналогично определяется удельная скорость потребления субстрата, (г/г • ч):
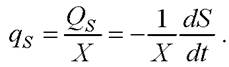
В некоторых процессах наряду с ростом биомассы происходит накопление в среде продукта метаболизма (его текущая концентрация Р).
Общая скорость биосинтеза продукта метаболизма QP в периодическом процессе, (г/г • ч):
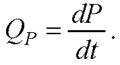
Удельная скорость биосинтеза продукта из единицы биомассы обозначается qР равна
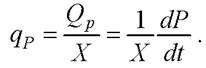
Наряду с кинетическими характеристиками, при математическом описании процессов ферментации часто пользуются и так называемыми макростехиометрическимихарактеристиками, которые выражают взаимосвязь между приростом биомассы, продукта и расходованием субстрата.
Простейшей такой характеристикой процесса ферментации является выход биомассы по субстрату, или экономический коэффициент. Его определяют, сравнивая количество выросшей за весь цикл ферментации биомассы Хк к количеству загруженного субстрата S0
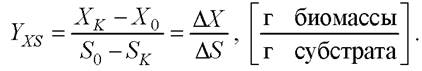
Выход продукта метаболизма по субстрату может быть вычислен аналогично:
![]()
1.4. Практическое значение микроводорослей в деятельности человека
Микроводоросли - самые быстрорастущие и высокоэнергетические растения. В настоящее время их применение чрезвычайно широко и разнообразно [3, 8 - 10].
1. Пища для человека. Во многих странах, особенно на Востоке, люди используют в пищу несколько видов водорослей, в том числе, микроводоросли. В Японии, например, сухая хлорелла применяется при изготовлении кисломолочного напитка «Yakult», хлеба, кондитерских изделий. Сухой порошок хлореллы и сценедесмуса применяют в качестве биологически активной добавки к пище [9].
2. Пища для водных животных. Водоросли лежат в основе пищевой пирамиды, поскольку синтезируют органические вещества из неорганических. Водные животные потребляют эту органику непосредственно или косвенно по пищевой цепи. Применение биомассы протококковых водорослей в качестве корма для рыбы при её промышленном производстве имеет большое значение для прудового рыбоводства. Некоторые виды рыб, например, толстолобик, питаются преимущественно водорослями. Кроме того, внесение микроводорослей в рыбоводные пруды способствует увеличению количества кормовых водных организмов, улучшает гидрохимическое состояние прудов, способствует насыщению кислородом, что приводит к увеличению продуктивности рыбных прудов до 25...35% [3].
3. Удобрение. Водоросли - ценное удобрение. Содержание почвенных микроводорослей во многом определяет плодородие участка. Микроводоросли обогащают почву органическими веществами, улучшают её структуру, стимулируют рост полезных почвенных микроорганизмов. Они являются источником физиологически активных веществ, играющих важную роль в почвенных процессах, влияют на рН почвенной среды, нейтрализуя её. Под влиянием водорослей водоудерживающая способность почв повышается на 40% и более. Водоросли обогащают почву макро- и микроэлементами, продуцируют антибиотически активные вещества, способствующие самоочищению почв, благоприятно влияют на рост и развитие растений и санитарно-эпидемиологическую обстановку. В почвах содержится много видов азотфиксирующих микроорганизмов, обогащающих почву азотом. Альголизация почв протококковыми микроводорослями может проводиться несколькими способами: замочкой семян в суспензии микроводорослей, внесением микроводорослей при орошении, внесением биомассы микроводорослей под зиму [3].
4. Кормовая добавка к пище сельскохозяйственных животных. Микроводоросли способны за короткий срок накапливать большое количество белка, липидов, витаминов, поэтому их биомасса по питательности приравнивается к кормам животного происхождения. Биомасса протококковых микроводорослей давно и успешно применяется в качестве добавки к пище различных сельскохозяйственных животных в сушёном виде, в виде паст и суспензий (свиноводство, овцеводство, кролиководство, разведение крупного рогатого скота и птицы). Такая кормовая добавка позволяет у различных видов животных увеличить привес живой массы, сократить падёж, увеличить перевариваемость грубых кормов, повысить удои и яйценоскость. Существуют примеры применения протококковых микроводорослей для подкормки тутового шелкопряда и пчёл. Из-за высокой биологической активности биомассы микроводорослей насекомые охотно потребляют водорослевый прикорм, особенно в весенне-зимний период, как следствие, повышается их жизнестойкость и продуктивность. Хороший результат обеспечивает введение биомассы протококковых микроводорослей в качестве кормовой добавки в рацион американской норки. Такая подкормка значительно повышает белковый обмен, стимулирует рост и развитие, повышает качество и размер шкурок [3].
5. Диатомит. Диатомит применяется в составе абразивных порошков и фильтров, а также служит теплоизоляционным материалом, заменяющим асбест.
6. Водорослевые культуры. Биологи уже давно выращивают водоросли в лабораториях. Сначала их выращивали в маленьких прозрачных чашках с прудовой водой на солнечном свету, а в последнее время применяют для этого особые культуральные среды с определённым количеством минеральных солей и специальных ростовых веществ, а также регулируемые источники искусственного света. Обнаружено, что для оптимального развития некоторых водорослей необходимы весьма специфические условия [3].
7. Применение микроводорослей в качестве сырья для биоэнергетики. В последнее десятилетие наблюдается снижение разведанных запасов высококачественной «лёгкой» нефти, истощение старых месторождений, а освоение новых сопряжено с возрастающими трудностями. Проблема усугубляется необходимостью снижения выброса парниковых газов в атмосферу. В качестве важнейшего компонента возобновляемых источников энергии рассматривают растительную биомассу. Использование биомассы микроводорослей связывают с появлением биотоплива «третьего» поколения. Считается, что биомасса микроводорослей как энергетическое сырьё по своим характеристикам превосходит другие сырьевые биоресурсы благодаря высокой продуктивности, большой скорости роста, нетребовательности к питательной среде и условиям культивирования [10, 11].
8. Комплексная технология переработки биомассы микроводорослей. Химический состав биомассы никоторых видов микроводорослей может широко варьироваться в зависимости от условий культивирования. Как и высшие растения, микроводоросли содержат нейтральные и полярные липиды. Нейтральные липиды состоят в основном из сложных эфиров глицерина. При благоприятных условиях микроводоросли производят в основном полярные липиды (например, фосфолипиды) [12]. Полярные липиды являются структурными компонентами всех живых клеток, входят в состав цитоплазматической, митохондриальной и других мембран, играют существенную роль в мембранной проницаемости, ответственны за расположение ферментов дыхательной цепи и перенос электронов. Поэтому фосфолипиды широко применяются в качестве пищевых добавок [10].
При неблагоприятных или ограниченных условиях роста микроводорослей (стрессовые условия культивирования) накапливаются нейтральные липиды в виде липидных капель в цитоплазме [13] и являются основными запасными компонентами клетки. Неполярные липиды находят широкое применение при производстве биотоплива, биополимеров и других продуктов технической биотехнологии.
Изменяя условия культивирования, можно получить биомассу микроводорослей с различным химическим составом. После извлечения из клеток целевых компонентов, остатки биомассы всё ещё будут содержать большое количество ценных биологически активных веществ. Анализируя все вышеперечисленные направления использования микроводорослей, можно сделать вывод о целесообразности разработки комплексной технологии переработки биомассы микроводорослей.
На рисунке 4 представлена схема комплексной переработки биомассы микроводоросли Chlorella vulgaris.
Рис. 4. Комплексное использование биомассы микроводорослей Chlorella vulgaris
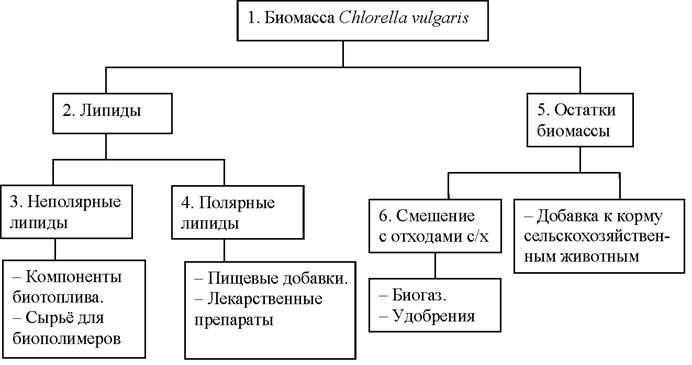
Технология комплексного использования биомассы микроводоросли Chlorella vulgaris заключается в следующем [10]. Штамм культивируется в течение 8 дней на стандартной питательной среде до достижения стационарной стадии роста, затем для клеток штамма создаются стрессовые условия путём пересадки на питательную среду с дефицитом азота для стимулирования накопления внутриклеточных липидов (выращивание в течение 3 дней). Затем клетки биомассы (1) отделяются от суспензии. На следующем этапе стенки клеток разрушаются для более полного извлечения липидов. Извлечение липидов осуществляется смесью полярного и неполярного растворителей, при этом фосфолипиды (4) будут содержаться во фракции полярного растворителя, а триглицериды (3) - неполярного.
Триглицериды после проведения реакции с метанолом (метанолиз) в присутствии катализатора образуют метиловые эфиры жирных кислот (компонент смесевого биодизельного топлива) и технический глицерин, который применятся во многих отраслях промышленности (производство моющих и косметических средств, сельское хозяйство, текстильная, бумажная и кожевенная отрасли промышленности, производство пластмасс, лакокрасочная промышленность, электротехника и радиотехника).
Извлечённые фосфолипиды подвергаются очистке и используются в качестве пищевой или биологически активной добавки к пище [14].
Остатки биомассы (5) (белки, углеводы) подвергаются очистке и используются как добавка для корма сельскохозяйственных животных, либо смешиваются с отходами сельского хозяйства и используются для получения биогаза, жидких и твёрдых удобрений.