МИКРОБИОЛОГИЯ Учебное пособие - 2012
ГЛАВА 12. НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ МИКРООРГАНИЗМОВ
Генетика — наука, изучающая закономерности и материальные основы наследственности и изменчивости организмов.
Наследственность — свойство организмов передавать своему потомству присущие им признаки.
Изменчивость — явление, заключающееся в возникновении различий между особями по ряду признаков или свойств. Другими словами — это свойство организмов приобретать новые признаки в процессе индивидуального развития.
Наиболее изучена генетика бактерий — организмов, характеризующихся малыми размерами и большой скоростью размножения, что позволяет проследить генетические изменения на большом числе популяций в течение небольшого промежутка времени. Бактериальная хромосома, содержащая основную генетическую информацию клетки, имеет длину около 1000 мкм, молекулярную массу 1,5—2,0 х 109 Д. Она спирализована, замкнута в кольцо, содержит от 3 до 5 тыс. генов. В цитоплазме бактерий располагаются также внехромосомные факторы наследственности: плазмиды, транспозоны и Is-элементы.
Плазмиды — нехромосомные молекулы ДНК небольшого размера, ответственные за какой-либо определенный признак. Так, фактор фертильности — F-фактор (от fertility — плодовитость) является плазмидой и содержит гены, контролирующие процесс конъюгации. R-фактор — фактор резистентности (resistence) несет гены, ответственные за устойчивость бактерий к лекарственным препаратам. Плазмиды могут определять вирулентность бактерий, например, у возбудителей чумы, столбняка, газовой гангрены и др.
Многие бактерии синтезируют белки, подавляющие рост родственных видов или видов, угнетающих их рост. Эти белки специфического действия называются бактериоцинами. Их синтез кодируется особыми плазмидами или бактериоциногенными факторами. Бактериоцины были выделены из клеток Escherichia coli (колицины), Pseudomonas aeruginosa (пиоцины), Lactobacillusacidophilus (лактоцины). Хромосома и плазмиды способны к автономному самокопированию — репликации, поэтому их называют репликонами.
Танспозоны — и Is-элементы — мигрирующие элементы, осуществляющие перенос генетической информации с одного репликона на другой. Транспозоны могут находиться в клетке в свободном состоянии, но не реплицируются самостоятельно, а только после включения в бактериальную хромосому. Is-элементы — это фрагменты ДНК длиной 1000 пар нуклеотидов. Они не обнаружены в свободном состоянии и представляют собой перемещаемые элементы, которые называют также «вставки последовательностей основании». В результате перемещения Is-элементов в различные участки ДНК происходит регуляция взаимодействия транспозонов, плазмид и умеренных фагов как между собой, так и с хромосомой бактерии; индуцирование мутаций при включении в хромосому.
Свойства микроорганизмов определяются генотипом, т. е. совокупностью всех генов, присущих данной особи. Фенотип представляет собой результат взаимодействия генотипа и окружающей среды, т. е. проявление генотипа в конкретных условиях обитания.
В основе изменчивости лежит либо изменение реакции генотипа на воздействие факторов внешней среды, либо изменение самого генотипа в результате мутации генов или их рекомбинации. Для прокариот, как и для эукариот, характерны два основных типа изменчивости — генотипическая (наследственная) и фенотипическая (модификационная):
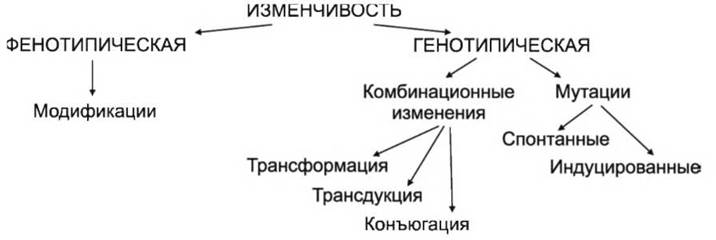
Фенотипическая изменчивость возникает как ответная реакция организма на изменение условий окружающей среды и проявляется у большинства особей в популяции. У ряда бактерий обнаружена уникальная адаптивная реакция в ответ на различные стрессовые воздействия. Стрессовое воздействие на бактериальную клетку вызывает ингибирование синтеза обычных белков, но индуцирует синтез небольшой группы белков, функция которых заключается в противодействии стрессу.
Модификация — изменение микроорганизмов под влиянием условий среды. Изменяются только фенотипические признаки. Модификации бактерий проявляются лишь временно, в период непосредственного воздействия фактора, и исчезают после его устранения. Роль фенотипической изменчивости сводится к обеспечению выживаемости микробной популяции в изменившихся условиях среды. Модификации проявляются в изменении морфологических, физиологических и других признаков с возвратом к прежнему фенотипу после устранения фактора, вызвавшего их.
Генотипическая изменчивость проявляется в виде мутаций и рекомбинации и осуществляется в результате изменении в первичной структуре генетического аппарата. Генотипические изменения возникают в природе как редкие события в жизни микробной популяции. Они не носят направленного характера и появляются спонтанно.
Мутация (от лат. mutatio — изменение) — стойкое наследственное изменение в структуре генетического аппарата. Механизм мутаций заключается в выпадении, вставке или замене одной пары нуклеотидов или группы нуклеотидов в молекуле ДНК, а также в изменении последовательности их расположений. По происхождению различают спонтанные и индуцированные мутации.
Спонтанные мутации возникают в популяции микробов без видимого внешнего воздействия и проявляются в самопроизвольном изменении генома клетки. Распространенными типами спонтанных мутаций являются устойчивость бактерий к фагам, ауксотрофность, резистентность к антибиотикам. Частота спонтанных мутаций составляет одну на 105— 106 клеток в популяции. В природе спонтанные мутации служат основным источником естественной изменчивости микроорганизмов, и именно они лежат в основе эволюционного процесса у про- и эукариот. Спонтанные мутации характеризуются изменением одного какого-нибудь признака и, как правило, стабильны.
Индуцированные мутации вызывают экспериментально воздействием какого-либо фактора — физического, химического или биологического.
К физическим относят различного рода излучения (рентгеновское, ультрафиолетовое, γ-излучение). Впервые возможность индуцированных мутаций показали в 1925 г. Г. А. Надсон и Г. С. Филиппов, воздействуя на дрожжевые клетки рентгеновскими лучами. Частота индуцированных мутаций значительно выше частоты спонтанных (одна на 103—104) и зависит от природы мутагенного агента и его дозы. Наиболее доступным мутагенным агентом является УФ-облучение с длиной волны около 260 нм. При этом механизм повреждения заключается в образовании димера тимина в молекуле ДНК, что приводит к блокированию нормального процесса репликации. При рентгеновском и у-облучении в ДНК бактерии возникают различные повреждения — разрывы цепей и химические изменения нуклеотидов. Эффект повреждения прямо пропорционален дозе облучения.
Химические мутагены обладают высоким мутагенным эффектом. По механизму действия на ДНК микробных клеток они существенно отличаются друг от друга. Действие одних приводит к изменению структуры ДНК. Так, аналог тимина — 5-бромурацил (5БУ), отличающийся от тимина только наличием атома брома на месте СН3-группы, спаривается не с аденином, как тимин, а с гуанином, в результате чего происходит замена пары АТ на 5БУГ. При воздействии азотистой кислоты, вызывающей дезаминирование азотистых оснований, цитозин превращается в урацил, а аденин — в гипоксантин. Урацил спаривается с аденином, а не с гуанином, как цитозин, в связи с чем происходит мутация — замена ГЦ на АТ. Акридиновые красители вызывают выпадения или вставки азотистых оснований в ДНК. Нитрозосоединения обладают множественным эффектом, вызывая высокую частоту мутаций, поэтому их называют супермутагенами. Наиболее сильными мутагенами являются нитрозометилмочевина, этиленимин, алкилирующие агенты (серный иприт нитрозогуанидин, этилметансульфат). Относительно безопасным мутагеном считается азотистая кислота (HN02).
Биологические факторы — это в первую очередь мигрирующие элементы: транспозоны, Is-элементы и ДНК вирусов. Перенос мигрирующих элементов (транспозиция) осуществляется с частотой от 10-4 до 10-6. В Is-элементах не закодировано никаких признаков, они содержат только информацию, необходимую для их переноса внутри клетки. У более сложно устроенных транспозонов имеются гены, ответственные за устойчивость клетки к антибиотикам, ионам тяжелых металлов и другим ингибиторам.
Умеренные фаги и определенные плазмиды служат переносчиками мигрирующих элементов между клетками. В результате встраивания мигрирующих элементов в бактериальную хромосому появляются мутантные клетки, у которых изменился порядок расположения нуклеотидов в триплете ДНК, вследствие чего нарушается процесс транскрипции. Доказано мутагенное действие вирусов и живых вирусных вакцин на млекопитающих.
По количеству мутировавших генов и характеру изменений в первичной структуре ДНК выделяют генные и хромосомные мутации.
Генные мутации затрагивают только один ген и чаще всего являются толковыми. Последние представляют собой выпадение, вставку или замену одной пары нуклеотидов.
Хромосомные мутации распространяются на несколько генов. Они носят характер крупных перестроек в отдельных фрагментах ДНК и проявляются в виде делеций (выпадения), инверсий (поворота на 180°), дупликаций (повторения какого-нибудь фрагмента ДНК). Один из механизмов образования хромосомных мутаций связан с перемещением Is-элементов и транпозонов из одного участка ДНК в другой или из репликона в репликон.
Комбинационная изменчивость. Ко второму типу наследственной изменчивости относятся изменения, возникающие у прокариот в результате рекомбинации генетического материала за счет частичного объединения геномов двух клеток. Известны три основных способа, приводящих к рекомбинации генетического материала прокариот: трансформация, трнсдукция и конъюгация, различающихся механизмами передачи хромосомной ДНК.
Трансформация (от лат. transformatio — преобразование, превращение) — непосредственная передача генетического материала из клетки донора в клетку реципиента путем проникновения фрагмента чужеродной ДНК. Явление трансформации у бактерий впервые доказал в 1928 г. английский микробиолог Ф. Гриффит.
Сущность эксперимента заключалась в следующем. Мышам одновременно были введены два штамма пневмококков: непатогенный, лишенный капсулы (R-штамм), и патогенный, имеющий капсулу (S-штамм), но убитый нагреванием. Большая часть мышей погибла от пневмонии. Из органов погибших мышей были выделены живые капсульные культуры пневмококков. Выяснилось, что культура убитого нагреванием капсульного пневмококка вызывала в организме животного трансформацию живых бескапсульных микробов, в результате чего у них появилась способность к образованию капсулы, что и обусловило патогенность. В 1944 г. группа ученых (О. Эйвери, К. Мак-Леод и М. МакКарти) поставили опыт Гриффитса in vitro: к культуре невирулентного бескапсульного штамма пневмококка добавили ДНК, полученную из вирулентного штамма пневмококка. Невирулентный штамм приобрел патогенные свойства. Таким образом ученые доказали, что трансформирующим агентом (в данном случае носителем вирулентных свойств пневмококка) является ДНК.
Трансформация бактерий происходит с частотой 10-2— 10-3 и зависит от вида микроорганизмов, свойств трансформирующей ДНК и состояния клетки реципиента. Способность ДНК проникать в клетку реципиента зависит как от природы самой ДНК, так и от физиологического состояния этой клетки. Клетки, способные воспринимать донорную ДНК, называются компетентными. Состояние компетентности непродолжительно. Оно возникает в определенный период роста бактериальной культуры, чаще всего в конце экспоненциальной фазы. В состоянии компетентности клеточная стенка бактерий становится проницаемой для высокополимерных фрагментов ДНК.
В клетку реципиента проникает очень малый фрагмент ДНК (соответствующий 0,3 % бактериальной хромосомы, или примерно 15 генам). Путем трансформации могут быть перенесены различные признаки: капсулообразование, способность к синтезу ферментов, устойчивость и чувствительность к антибиотикам.
При трансформации рекомбинация происходит лишь в том случае, если ДНК бактерий донора и реципиента родственны друг другу, но имеют разный генотип.
Трансдукция (от лат. transductio — перенос, перемещение) — передача генетического материала из клетки донора в клетку реципиента с участием умеренного бактериофага. Явление трансдукции открыли в 1952 г. Н. Циндер и Дж. Ледерберг на примере двух штаммов сальмонелл.
Механизм трансдукции заключается в следующем. В процессе репродукции некоторых умеренных фагов небольшой фрагмент бактериальной хромосомы, содержащий один ген или несколько сцепленных генов, включается в геном фага. Трандуцирующий фаг переносит фрагмент ДНК предыдущего хозяина в другую, чувствительную к фагу клетку.
Различают три типа трансдукции: общую, специфическую и абортивную. При общей трансдукции происходит перенос разных признаков одновременно; специфическая трансдукция предусматривает перенос только определенного признака; при абортивной трансдукции участок чужеродной ДНК, перенесенный бактериофагом в клетку реципиент, не включается в ее геном, поэтому рекомбинации не происходит и новый признак не наблюдается.
В естественных условиях перенос генетического материала с помощью фагов может быть самым распространенным механизмом рекомбинации у прокариот. В экспериментах по генной инженерии трансдукция открывает возможности не только межвидовой гибридизации бактерий, но и возможности получения гибридов среди таксономически отдаленных групп прокариот.
Конъюгация (от лат. conjugation — соединение) — процесс установления клеточного контакта между двумя различающимися в половом отношении бактериями. Это явление было обнаружено в 1946 г. Д. Ледербергом и Э. Татумом у мутантных штаммов Е. coli. При конъюгации у бактерий происходит направленный перенос генетического материала из клетки донора в клетку реципиента.
Способность бактерий к конъюгации связана с наличием у некоторых из них полового фактора (F-фактор, от англ. fertility — плодовитость). Такие клетки обозначают F+. F-фактор располагается в цитоплазме клетки в виде кольцевой ДНК, т. е. является плазмидой. Клетки, не имеющие F-фактора, являются реципиентами и обозначаются F-.
Процесс конъюгации начинается с прикрепления конца F-пили клетки донора к клетке реципиента. В течение нескольких минут обе клетки сближаются, а затем через конъюгационный мостик менее чем за 5 мин происходит передача полового F-фактора независимо от бактериальной хромосомы из клетки донора (F+) в клетку реципиента (F-). При этом клетка донор не теряет своей донорской способности, так как в ней остаются копии F-фактора. Если F-фактор включается в хромосому, то бактерии приобретают способность передавать фрагменты хромосомной ДНК и называются Hfr-клетками (от англ. high frequency of recombination — высокая частота рекомбинаций). При конъюгации клеток Hfr и клеток F- одна из нитей хромосомы разрывается и передается с определенного участка в клетку F-. Оставшаяся в клетке донора нить ДНК является матрицей для второй нити, а вторая переносимая ДНК донора взаимодействует с ДНК реципиента — происходит гомологичная рекомбинация. Прерывая процесс конъюгации бактерий, можно определять последовательность расположения генов в хромосоме. При конъюгации происходит только частичный перенос генетического материала, поэтому ее не следует отождествлять с половым процессом у других организмов.
12.1. ЭЛЕМЕНТЫ ГЕННОЙ ИНЖЕНЕРИИ
Общеизвестно, что скрещивание или гибридизация среди представителей царства эукариот осуществляется только между близкородственными организмами. У прокариот посредством передачи плазмид гибридизация не ограничивается рамками даже крупных систематических категорий. В связи с этим прокариоты открывают беспредельные возможности нового научного направления — генной инженерии, заключающегося в конструировании гибридов из материала совершенно разного происхождения.
Предпосылками к исследованиям в области генной инженерии послужили два открытия, сделанные в первой половине XX в.: во-первых, это явление трансдукции, а во-вторых — присутствие в резистентных к бактериофагу бактериях специальных ферментов, которые разрезают двойные спирали фаговых ДНК в строго определенных местах. Эти ферменты назвали рестриктазами. В настоящее время получено около 500 рестриктаз из различных бактерий, обладающих специфичностью к определенным участкам (сайтам) ДНК. Обнаружение плазмид в бактериях легло в основу технологии получения рекомбинантных ДНК.
Генная инженерия — это методы получения рекомбинантных ДНК с последующим их включением в реципиентную клетку. В основу генноинженерных методов заложена способность ферментов рестриктаз расщеплять ДНК на отдельные нуклеотидные последовательности, которые могут быть использованы для встраивания их в геномы бактериальных плазмид или фагов с целью получения гибридных или химерных форм, состоящих из собственной ДНК и дополнительных встроенных фрагментов несвойственной им ДНК.
Для получения рекомбинантных ДНК в первую очередь необходимо иметь хорошо отработанную систему хозяин-вектор. Под вектором понимается небольшая молекула ДНК, способная акцептировать чужеродные фрагменты ДНК и реплицироваться в определенном организме. В качестве вектора могут служить вирусы животных и растений, бактериофаги или плазмиды. Необходимо иметь также эффективный способ введения векторной и рекомбинантной молекулы в микроорганизм. Векторные молекулы должны обладать рядом свойств, позволяющих удобное введение чужеродной ДНК и ее последующую экспрессию. В векторных молекулах есть отдельные участки, которые рестриктазы узнают и расщепляют в них ДНК с образованием «липких концов». В расщепленные участки с помощью фермента ДНК-лигазы «вшиваются» нужные чужеродные гены, имеющие такие же липкие концы. Сконструированные таким способом рекомбинантные ДНК вводят с помощью трансформации в бактериальную клетку или другие клетки живых организмов, после чего автономная ДНК приобретает способность к автономному функционированию и при делении клетки реплицируется.
В качестве вектора применяют также космиды, представляющие собой гибрид плазмиды с фагом. Космиды используют для клонирования больших (до 45 тыс. пар нуклеотидов) фрагментов ДНК эукариот.
Генная инженерия имеет огромное прикладное значение. Получены не существующие в природе штаммы микроорганизмов — продуцентов ценных для человека белков и пептидов, в частности штаммы Е. coli, синтезирующие инсулин. Метод получения человеческого инсулина путем выращивания рекомбинантного штамма Е. coli решил проблему обеспечения людей, больных диабетом, этим препаратом. Для производства препаратов, получаемых из дефицитного или дорогостоящего сырья, также используют метод генной инженерии. Ведутся работы по совершенствованию штаммов дрожжей, используемых в пивоварении и виноделии. Этим организмам передаются гены, которые могут обеспечить усвоение пентоз, разрушение фенольных соединений, конкурентоспособность при росте в нестерильных условиях.