МИКРОБИОЛОГИЯ Учебное пособие - 2012
ГЛАВА 3. МОРФОЛОГИЯ БАКТЕРИЙ
3.3. СТРОЕНИЕ ПРОКАРИОТИЧЕСКОЙ КЛЕТКИ
Тонкое строение клетки изучают с помощью электронного микроскопа и микротома, позволяющего приготовить ультратонкие срезы клеток. Основными структурами бактериальной клетки являются: клеточная стенка, цитоплазматическая мембрана, цитоплазма с включениями и нуклеоид. Некоторые бактерии имеют дополнительные структуры: капсулу, жгутики, пили. Бактерии определенных видов способны образовывать эндоспоры (рис. 3).
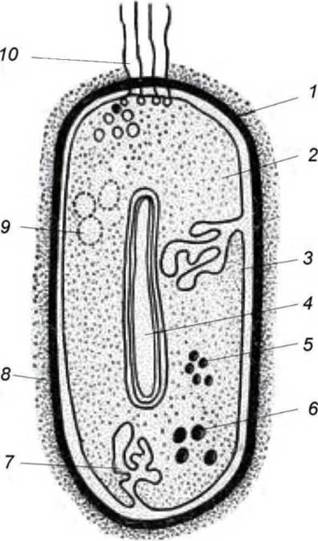
Рис. 3. Схема строения бактериальной клетки:
1 — клеточная стенка; 2 — цитоплазма с рибосомами; 3 — ЦПМ; 4 — нуклеоид; 5 — полифосфаты; 6 — липиды; 7 — мезосомы; 8 — капсула; 9 — включения серы; 10 — жгутики
Клеточная стенка — тонкая бесцветная структура, которая придает клетке определенную форму и вместе с прилежащей к ней цитоплазматической мембраной позволяет удерживать высокое внутриклеточное давление. Клеточная стенка прокариот выполняет разнообразные функции: обеспечивает поддержание постоянной формы клетки, участвует в процессе деления клетки, механически защищает клетку от воздействий окружающей среды, от избыточного проникновения в нее воды. На долю клеточной стенки приходится от 5 до 20 % сухих веществ клетки.
Основной каркас клеточной стенки бактерий составляет полимер, называемый пептидогликаном или муреином (от лат. murus — стенка). Способность синтезировать полимер такого рода свойственна только прокариотам (рис. 4).

Рис. 4. Схема строения фрагмента муреина
Муреин — гетерополимер, состоящий из параллельно расположенных молекул гликана, соединенных поперечной пептидной связью. Гликан образован чередующимися остатками N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью типа р (1 → 4). Остатки мурамовой кислоты через лактильные группы соединены с аминокислотами (рис. 5).

Рис. 5. Однослойная структура пептидогликана (Красильников, 1974)
В состав пептидного компонента муреина обычно входят L-аланин, Д-глутаминовая кислота, D-аланин и специфическая для прокариот диаминокислота — мезо-диаминопимелиновая. У некоторых бактерий она может быть замещена либо на L-лизин, либо на L-орнитин. Диаминокислоты играют большую роль в межмолекулярных сшивках гликановых слоев. Пептидными мостиками гетерополимерные цепи связаны между собой в муреиновый мешок. С муреином ковалентно связаны тейхоевые кислоты, представляющие собой полимеры рибит-фосфата или глицеринфосфата. Тейхоевые кислоты расположены по обе стороны пептидогликанового слоя, причем на наружной стороне их меньше, чем на стороне, прилегающей к цитоплазматической мембране.
Следует особо отметить, что в клеточной стенке бактерий содержатся вещества, которых нет у животных и растений, — муреин, тейхоевые кислоты и мезо-диаминопимелиновая кислота, не встречающаяся в составе белков.
Структура клеточной стенки была выяснена, в основном, в связи с изучением действия лизоцима и пенициллина на бактерии. Открытый
А. Флемингом лизоцим является бактерицидным ферментом, содержащимся в слезной жидкости, в носовой слизи, яичном белке и женском молоке. Лизоцим выделен также из бактерий и бактериофагов. При воздействии лизоцима на суспензию грамположительных бактерий наблюдают быстрое ее просветление. Лизоцим разрывает в муреине гликозидную связь между N-ацетилглюкозамином и N-ацетилмурамовой кислотой, являясь таким образом ферментом мурамидазой. Наряду с лизоцимом существует много других ферментов, лизирующих муреиновый каркас.
Антибиотик пенициллин действует, главным образом, на грамположительные бактерии. Он препятствует поперечной сшивке муреина, разрывая пептидную связь. Однако бактерицидное действие пенициллина наблюдается только на активно размножающихся клетках; старые клетки остаются жизнеспособными. При воздействии пенициллина на бактериальную клетку появляются особые формы бактерий — протопласты (клеточная стенка лизирована полностью) и сферопласты (клеточная стенка разрушена не полностью). Протопласты и сферопласты сохраняют обмен веществ, способность к росту и иногда к размножению. Такие бактерии принято называть L-формами. Морфологически они представляют собой шаровидные или неправильной формы клетки от мельчайших размеров до гигантских.
Окрашивание бактерий по Граму. По химическому составу и тонкому строению клеточной стенки прокариоты подразделяют на две большие группы — грамположительные и грамотрицательные. В 1884 г. датский ученый Христиан Грам предложил метод окраски тканей, который впоследствии был использован в практике микробиологии как один из важнейших морфологических признаков, используемых при определении вида бактерий, и получил название окраски по Граму. Сущность окраски состоит в следующем: клетки фиксируют на предметном стекле прогреванием, препарат окрашивают фиолетовым красителем (кристаллическим фиолетовым или генцианвиолетом), краситель смывают и обрабатывают сначала раствором йода, а затем органическим растворителем (спиртом или ацетоном), после чего дополнительно окрашивают контрастным красителем — фуксином.
Йод образует с фиолетовым красителем стойкий окрашенный комплекс. При обработке спиртом у грамположительных бактерий этот комплекс удерживается клеткой, и она остается окрашенной в фиолетовый цвет, а у грамотрицательных бактерий окрашенный комплекс вымывается из клетки, и она обесцвечивается. После окрашивания фуксином грамотрицательные бактерии приобретают красную окраску.
Клеточная стенка грамположительных бактерий достаточно массивна, толщина ее составляет 20—80 нм. Она имеет гомогенную трубчатую структуру, пронизанную порами, и плотно прилегает к ЦПМ. На долю муреина приходится 30—70 % сухой массы клеточной стенки. С муреином связаны тейхоевые кислоты. Они присутствуют только у грамположительных бактерий и представляют собой полимеры на основе рибита или глицерина, остатки которых соединены между собой фосфодиэфирным связями. Свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилгюкозамина и других соединений.
Обработка спиртом при окраске по Граму вызывает разбухание муреина и уменьшение диаметра пор клеточной стенки, что в целом приводит к снижению ее проницаемости, в результате чего окрашенный комплекс не вымывается из клетки.
Клеточная стенка грамотрицателъных бактерий многослойна и более разнообразна по химическому составу (рис. 6). Внутренний слой клеточной стенки представлен муреином, на долю которого приходится от 1 до 10 % ее сухой массы. Его толщина составляет лишь 2—3 нм. Муреиновый слой отделен от мембраны периплазматическим пространством, в котором содержатся ферменты. Со слоями муреина ковалентно связаны липопротеины. Их липофильные концы обращены наружу. Внешний слой клеточной стенки (наружная мембрана), имеющий толщину 8—10 нм, образован фосфолипидами, липополисахаридами и белками и связан с муреином посредством липопротеинов. Гидрофобные концы фосфолипидов и липополисахаридов обращены вовнутрь, а гидрофильные головы их — наружу. В двойной липидный слой наружной мембраны встроены белки — порины, пронизывающие этот слой насквозь. Порины пропускают через мембрану гидрофильные низкомолекулярные вещества (порядка 6000 Д). Липополисахарид (ЛПС) наружной мембраны состоит из трех компонентов: липида А, сердцевинной зоны (или ядра) и О-специфической боковой цепи полисахарида — О-антигена. Липид А придает токсичность липополисахариду, в связи с чем ЛПС относятся к наиболее действенным эндотоксинам бактерий.

Рис. 6. Модель строения клеточной стенки грамотрицательных бактерий (Шлегель, 1987):
Н — цитоплазматическая мембрана; ПП — периплазматическое пространство; М — муреин; НМ — наружная мембрана
Липополисахариды приобрели большое значение в бактериологической диагностике и в распознавании эпидемий. О-специфические боковые цепи ЛПС разных видов и штаммов бактерий отличаются друг от друга по составу. Они определяют серовар (разновидность бактерий, выявляемая серологическим методом с помощью иммунной сыворотки) определенного штамма бактерий.
Так как у грамотрицательных бактерии муреиновыи слои тонкий, он не играет существенной роли в проницаемости стенки. Кроме того, значительное содержание липидов в клеточной стенке способствует вымыванию окрашенного комплекса из клетки после обработки спиртом мазка при окраске по Граму.
В табл. 3 приведены некоторые представители грамположительных и грамотрицательных бактерий.
Таблица 3. Некоторые представители грамположительных и грамотрицательных бактерий
Грамположительные бактерии |
Грамотрицательные бактерии |
Микрококки, сарцины, стафилококки, молочнокислые, пропионовокислые, маслянокислые бактерии, бациллы, клостридии, бифидобактерии |
Кишечные палочки, сальмонеллы, возбудители дизентерии, холеры, бруцеллеза, брюшного тифа, псевдомонады, палочки протея, уксуснокислые бактерии |
Следует подчеркнуть, что окраска по Граму не всегда служит вполне надежным методом дифференциации этих двух групп бактерий. Некоторые грамположительные бактерии с возрастом быстро теряют способность удерживать окрашенный комплекс генцианвиолета с иодом. Поэтому для окрашивания по Граму нужно использовать молодые (18—24-часовые) культуры. Кроме того, отдельные грамположительные бактерии могут давать грамвариабельную реакцию.
Прокариоты, не имеющие клеточной стенки, встречаются в природе. Это самые мелкие прокариоты — сапрофиты или паразиты, способные размножаться самостоятельно в организме хозяина и вызывающие хронические инфекции. Их называют микоплазмами. В частности, Mycoplasma pneumoniae вызывает у человека заболевание по типу острой респираторной инфекции. Из-за отсутствия клеточной стенки эти бактерии не имеют определенной формы и весьма полиморфны.
Обнаружены прокариоты, у которых клеточная стенка по составу и структуре существенно отличается от вышеописанного пептидогликана. Эти микроорганизмы принадлежат к группе архебактерий. Так, у галофильных архебактерий рода Ualobacterium клеточная стенка состоит в основном из гликопротеинов, а у архебактерий рода Halococcus — из гетерополисахаридов. У метанобразующих бактерий обнаружены клеточные стенки трех типов: состоящие из псевдомуреина, построенные из белковых глобул и из гетерополисахаридов.
Уникальность химического состава клеточной стенки прокариот позволила создать целый ряд лекарственных препаратов, специфически действующих только на эту структуру бактерий. К таким препаратам относятся пенициллин, бацитрацин, новобиоцин и другие антибиотики.
Цитоплазматическая мембрана (ЦПМ) (от лат. membrane — кожица, оболочка) расположена между клеточной стенкой и цитоплазмой. Она является обязательным структурным элементом любой клетки и представляет собой белково-липидный комплекс, в состав которого входит от 50 до 75 % белков и от 15 до 45 % липидов. Толщина ЦПМ обычно составляет 4—7 нм.
В составе мембраны обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды (холестерин). Основную структурную роль в ЦПМ играют фосфолипиды. Они формируют двухслойную структуру, в которой гидрофильные головы фосфолипидов обращены наружу, а гидрофобные «хвосты» (это, как правило, две жирнокислотные цепи) — вовнутрь. Белковые глобулы плавают в фосфолипидном слое, будучи погружены в него полностью (интегральные белки), или находятся на поверхности (периферийные белки) (рис. 7).

Рис. 7. Модель строения плазматической мембраны:
1 — углеводные фрагменты гликопротеидов; 2 — двойной фосфолипидный слой; 3 — интегральный белок; 4 — головы фосфолипидов; 5 — периферический белок; 6 — холестерин; 7 — жирнокислотные хвосты фосфолипидов
Белковые глобулы формируют каналы, которые выполняют функции насосов — с помощью АТФ осуществляют движение веществ и ионов (натрия, калия, кальция и др.). Натрий-калиевые насосы перекачивают Na+ из клетки, а К+ — в клетку (антипорт) против их химических градиентов. Перенос ионов осуществляется основным компонентом насоса — натрий-калий-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия.
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют, как мобильную структуру. Мембрана играет важнейшую роль в обмене веществ клетки. Она служит осмотическим барьером клетки и контролирует как транспорт питательных веществ внутрь клетки, так и вывод метаболитов наружу. Мембрана ответственна также за биоэнергетические процессы, так как ферменты переноса электронов и окислительного фосфорилирования (цитохромы, железосерные белки и другие компоненты) локализуются внутри или на поверхности мембраны.
ЦПМ участвует в превращениях клеточной энергии. Бактерии получают энергию в процессах дыхания или фотосинтеза, а в ЦПМ расположены переносчики цепи электронного транспорта, которые генерируют электрохимическую энергию (ΔДμн+), используемую в клетке с различными целями, в том числе и для образования химической энергии — АТФ. В мембране расположены и ферментные комплексы, катализирующие превращение электрохимической энергии в АТФ (ΔДμн+→ АТФ).
Кроме того, на цитоплазматической мембране локализуется центр репликации бактериальной хромосомы.
Мезосомы. Темпы роста цитоплазматической мембраны обычно опережают темпы роста клеточной стенки, в результате чего мембрана часто образует многочисленные инвагинации (впячивания вовнутрь) различной формы, называемые мезосомами. Мезосомы бактерий довольно разнообразны по форме, размерам и локализации в клетке. Наиболее просто организованнные мезосомы имеют форму везикул (пузырьков), более сложно устроены ламиллярные (пластинчатые) и тубулярные (трубчатые) мезосомы. По расположению в клетке различают мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы), мезосомы, к которым прикрепляется нуклеоид перед делением клетки (ядерные), и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ (периферические).
Цитоплазма отделена от клеточной стенки цитоплазматической мембраной и представляет собой сложную коллоидную систему, состоящую на 80—85 % из воды и растворимых белков, ферментов, РНК, субстратов и продуктов метаболических реакций. В цитоплазме содержатся рибосомы, запасные питательные вещества: полисахариды, поли-β-оксимасляная кислота, полифосфаты, сера, железо и другие соединения.
Рибосомы — рибонуклеопротеидные частицы диаметром 15...20 нм, состоящие на 2/3 из РНК и на 1/3 из белков. Их основная функция — биосинтез белка. Количество рибосом в клетке колеблется от 5 до 90 тыс. Рибосомы прокариот имеют константу седиментации 70 единиц Сведберга (S); их называют 70S-рибосомами. Каждая рибосома состоит из двух субъединиц: 30S и 50S. Во время активного синтеза белка в клетке образуются правильные цепочки рибосом, напоминающие бусы; их называют полирибосомами или полисомами.
Нуклеоид прокариот представлен молекулой ДНК, которая формирует структуру, имеющую вид замкнутого кольца, компактно упакованную и занимающую определенную область в цитоплазме. Молекулярная масса нуклеоида (1—3) х 109 Д. Нуклеоид не отделен от цитоплазмы ядерной мембраной. В молекуле ДНК сосредоточена почти вся генетическая информация клетки, поэтому ее называют также бактериальной хромосомой. Обычно в бактериальной клетке содержится одна хромосома. У бактерий с крупными клетками (нитчатые цианобактерии) часто обнаруживают несколько нуклеоидов — до 8.
Внехромосомные генетические элементы. У многих бактерий обнаружены внехромосомные генетические элементы: плазмиды, умеренные фаги, транспозоны и IS-элементы.
Плазмиды — нехромосомные молекулы ДНК небольшого размера, ответственные за какой-либо определенный признак. Так, фактор фертильности — F-фактор (от fertility — плодовитость) является плазмидой и содержит гены, контролирующие процесс конъюгации. R-фактор — фактор резистентности (resisfence) — несет гены, ответственные за устойчивость бактерий к лекарственным препаратам. Плазмиды могут определять вирулентность бактерий, например, у возбудителей чумы, столбняка, газовой гангрены и др.
Многие бактерии синтезируют белки, подавляющие рост родственных видов или видов, угнетающих их рост. Эти белки специфического действия называются бактериоцинами. Их синтез кодируется особыми плазмидами или бактериоциногенными факторами. Бактериоцины были выделены из клеток Escherichia coli (колицины), Pseudomonas aeruginosa (пиоцины), Bacillusmegatherium (мегацины), Lactobacillus acidophilus (лактоцины).
Плазмиды, очевидно, контролируют также синтез особых белков — прионов — новых агентов инфекционных болезней. Они открыты в начале 70-х гг. XX в. нейробиологом Калифорнийского университета в Сан-Франциско Стэнли Прузинером. По предположению С. Прузинера, в зависимости от среды обитания белок подвергается генетической мутации, изменяется его стереоструктура. Он приобретает инфекционные свойства, вызывает гибель нейронов, на их месте образуются ячейки, губчатость, и как результат — нарушается нервная система, отсюда название заболевания: губчатая энцефалопатия или губчатый энцефалит. За изучение болезнетворного агента, вызывающего губчатую энцефалопатию или «коровье бешенство» у крупного рогатого скота, С. Прузинеру в 1997 г. была присуждена Нобелевская премия по физиологии и медицине.
Транспозоны и IS-элементы, называемые также мигрирующими элементами, представляют собой линейные молекулы двунитчатой ДНК, включающие от 200 до 6000 пар нуклеотидов. Эти элементы могут встраиваться в разные участки бактериальной хромосомы или мигрировать с бактериальной хромосомы на плазмиду. Характерно, что транспозоны и IS-элементы не способны к автономной репликации; их репликация происходит одновременно с соответствующей хромосомой или плазмидой. Перенос мигрирующих элементов (транспозиция) осуществляется с частотой от 10-4 до 10-6. В IS-элементах не закодировано никаких признаков, они содержат только информацию, необходимую для их переноса внутри клетки. У более сложно устроенных транспозонов имеются гены, ответственные за устойчивость клетки к антибиотикам, ионам тяжелых металлов и другим ингибиторам.
Умеренные фаги и определенные плазмиды служат переносчиками мигрирующих элементов между клетками. В результате встраивания мигрирующих элементов в бактериальную хромосому появляются мутантные клетки, у которых изменился порядок расположения нуклеотидов в триплете ДНК, вследствие чего нарушается процесс транскрипции.
Запасные вещества. У прокариот к резервным веществам относятся полисахариды, липиды, полифосфаты, соединения серы, углеводородные гранулы. Эти вещества накапливаются в клетке, если в питательной среде содержатся соответствующие соединения. При недостатке внешних источников энергии они включаются в метаболизм и могут продлить существование клетки.
Из полисахаридов в клетках бактерий обнаруживаются гликоген и крахмал. Анаэробные спорообразующие бактерии — клостридии способны накапливать крахмалоподобное вещество — гранулезу. Запасные полисахариды, в отличие от полисахаридов клеточной стенки, все образуются из α-D-глюкозы.
Липиды накапливаются в виде гранул и капелек жира, которые в световом микроскопе сильно преломляют свет. Липиды многих бактерий состоят из поли-β-гидроксимасляной кислоты. Поли-β-гидроксимасляную кислоту образуют многие аэробные бактерии, цианобактерии и факультативные фототрофные бактерии. Микобактерии в качестве резервных липидов накапливают воска (сложные эфиры высших жирных кислот и спиртов).
Полифосфаты. Многие бактерии способны накапливать фосфорную кислоту в виде гранул полифосфатов. Впервые такие гранулы были описаны у Spirillum volutans, поэтому их называют также гранулами волютина или метахроматиновыми зернами, так как при окрашивании препарата бактерий метиленовым синим цитоплазма окрашивается в голубой, а гранулы полифосфатов — в красно-фиолетовый цвет. Полифосфаты используются клетками как источник фосфора и энергии.
Сера накапливается серными бактериями в присутствии сероводорода в среде и окисляется до сульфата, когда весь сероводород оказывается исчерпанным. Для аэробных бесцветных серных бактерий сера служит источником энергии, а для анаэробных фототрофных пурпурных серобактерий — донором электронов.
Жгутики. На поверхности клеток многих бактерий имеются структуры, определяющие их подвижность в жидкой среде. Наличие, число, размеры и расположение жгутиков являются признаками, которые характерны для данного вида бактерий и имеют важное таксономическое значение. Однако эти признаки непостоянны и зависят от возраста и условий культивирования. Старые клетки, как правило, неподвижны, поэтому для определения подвижности используют молодые культуры. У палочковидных бактерий жгутики могут располагаться полярно (на полюсах клетки), или латерально (вдоль боковой поверхности клетки). В зависимости от числа жгутиков и их расположения на поверхности клетки различают по следующим типам:
✵ монополярный монотрих — один жгутик прикреплен к одному из полюсов клетки (рис. 8; 1);
✵ монополярный политрих или лофотрих — пучок жгутиков прикреплен на одном из полюсов клетки (рис. 8; 2 — а, б, в);
✵ биполярный монотрих — по одному жгутику на обоих полюсах клетки;
✵ биполярный политрих или амфитрих — по пучку жгутиков на каждом полюсе клетки (рис. 8; 3);
✵ перитрих — многочисленные жгутики расположены по всей поверхности клетки (рис. 8; 4).

Рис. 8. Основные типы жгутикования у бактерий (Шлегель, 1987)
У извитых форм жгутики обнаружены позже с помощью электронной микроскопии. Из этого следует, что извитые формы могут передвигаться не только за счет изгибов тела, но и с помощью жгутиков. Шаровидные формы бактерий, за исключением Sarcina urea, жгутиков не имеют.
Жгутики представляют собой спирально закрученные нити, состоящие из специфического белка флагеллина. Толщина жгутиков 12—20 нм, длина от 3 до 20 мкм. Жгутик состоит из трех частей: спиральной нити (фибриллы), «крюка» и базального тела. Нить жгутика крепится к крюку, представляющему собой изогнутый белковый цилиндр. Крюк, в свою очередь, соединен с базальным телом, закрепленным в плазматической мембране.
Жгутики осуществляют вращательное движение наподобие корабельного винта со скоростью 10—20 с-1. При прямолинейном движении клетки вперед жгутик вращается против часовой стрелки, а при перемещении назад — по ходу часовой стрелки. Если клетка несет много жгутиков, они при движении собираются в пучок.
Установлено, что подвижные бактерии способны реагировать на малейшие изменения внешней среды перемещением в определенном направлении. Такие перемещения называют таксисом, при этом различают хемотаксис, фототаксис, магнитотаксис, термотаксис, вискозитаксис. Наибольшее внимание привлекает хемотаксис — движение в определенную сторону относительно источника какого-либо химического вещества. Эти химические вещества относят либо к аттрактантам (привлекающим бактерии), либо к репеллентам (отпугивающим бактерии). Аттрактантами могут быть витамины, аминокислоты, сахара; репеллентами — спирты, щелочи, кислоты, фенолы. Молекулярный кислород для аэробных бактерий является аттрактантом, а для анаэробных — репеллентом. Аттрактантами часто могут быть пищевые субстраты.
Ворсинки (фимбрии, пили) — длинные неподвижные нитевидные структуры, находящиеся на поверхности некоторых бактерий. Фимбрии имеют цилиндрическую форму, длиной 0,2—2,0 мкм и внутренним диаметром 5—10 нм. Они состоят из белка, называемого пилином. По морфологии, антигенным свойствам и выполняемым функциям различают несколько типов фимбрий. Фимбрии первого типа выполняют функцию прикрепления клетки к поверхности субстрата или сцепления друг с другом. Фимбрии второго (общего) типа ответственны за питание и водно-солевой обмен клетки. Количество фимбрий общего типа достигает нескольких тысяч. Термин пили применяется для обозначения третьего типа фимбрий (F-пили) — половых, образуемых клетками-донорами. Их количество — 1—3 на клетку. F-пили обеспечивают контакт клетки-донора с клеткой-реципиентом, образуя конъюгационный канал, по которому происходит передача генетической информации между ними.
Капсула. Многие прокариоты синтезируют органические полимеры, которые откладываются с наружной стороны клеточной стенки в виде аморфного слоя, называемого капсулой или слизистым чехлом. В зависимости от толщины слизистого слоя принято различать: микрокапсуду, видимую лишь под электронным микроскопом, толщиной до 0,2 мкм, макрокапсулу — слизистый слой, толщиной более 0,2 мкм, и слизистый слой — слизистое вещество, окружающее клетку и по толщине часто превосходящее ее. У некоторых сапрофитных бактерий образуется общая капсула для многих особей. Скопления микроорганизмов, заключенных в общую капсулу, называют зооглеями.
Капсула и слизь выполняют защитную функцию — предохраняют клетку от высыхания, механических повреждений, препятствуют действию защитных факторов макроорганизма, проникновению бактериофага.
Капсула или слизь не являются обязательной структурой бактериальной клетки. При удалении слизистого слоя бактерии продолжают размножаться. У многих бактерий имеются капсульные и бескапсульные варианты. Обладание капсулой зависит от штамма микроорганизма, возраста культуры и условий выращивания. Некоторые бактерии, образующие капсулу, могут легко в результате мутации превращаться в бескапсульные формы. Этот процесс получил название S → R-трансформации. Колонии, состоящие из клеток, окруженных капсулой, имеют гладкую поверхность; их обозначают S-колонии (от англ. smooth — гладкий), а колонии, образованные бескапсульными бактериями, имеют шероховатую поверхность и называются R-колониями (от англ. rough — шероховатый).
Капсула в большинстве случаев состоит из полисахаридов гомо- или гетерополимерной природы. В первом случае они построены из сахарных остатков одного типа (глюканы, маннаны), во втором — из различных сахарных остатков. Капсулы некоторых бактерий рода Bacillus состоят из полипептидов — полимеров D- и L-форм глутаминовой кислоты. В образовании экзополисахаридов капсулы принимают участие клеточная стенка и ЦПМ. Однако существует и другой механизм их образования — при действии внеклеточных ферментов на дисахариды, в результате чего образуются декстраны и леваны. Обильное образование слизи наблюдается при размножении Leuconostoc subsp. dextranicum в среде, содержащей сахарозу. Этот представитель гетероферментативных молочнокислых бактерий быстро превращает сахарозу, содержащуюся в растворе, в декстран. Такое превращение происходит вне клетки и синтезируется внеклеточным ферментом — декстрансахаразой. Декстран — полисахарид, состоящий из остатков α-D-глюкозы, соединенных в положении 1,6, т. е. это 1,6-α-глюкан.
Стрептококки видов Streptococcus mutans и вызывающие кариес зубов, выделяют фермент, превращающий сахарозу в полифруктозу — леван. Этот полисахарид откладывается на поверхности зубов, и в нем накапливаются кислые продукты брожения, вызываемого стрептококками, главным образом, молочная кислота.
В настоящее время внеклеточные полимеры, синтезируемые определенными видами бактерий, находят практическое применение: их используют в качестве заменителя плазмы крови и для получения синтетических пленок.
Капсулу обнаруживают специальными методами окраски. К препарату добавляют тушь, конго красный или нигрозин, которые в капсулу не проникают, в связи с чем получается негативное контрастирование: светлая капсула выделяется на темном фоне.
Пигменты бактерий. Колонии многих бактерий бывают окрашены в разные цвета: желтый, оранжевый, розовый, красный, желто-зеленый, синий и др. Это обусловлено выделением окрашенного продукта во внешнюю среду или пигментацией самой клетки. Способность синтезировать пигменты заложена генетически и может быть использована в качестве характерного признака при определении вида бактерий. Пигменты обычно представлены разными классами веществ: каротиноидами, пирролами, азахинонами, антоцианами и др. Например, бактерия Serratia marcescens образует на питательной среде колонии ярко-красного цвета, напоминающие капли крови, за что она получила название «чудесной палочки». Бактерии этого вида синтезируют характерный красный клеточный пигмент продигиозин — производное трипиррола. Пигменты красного цвета могут синтезировать также актиномицеты и серные пурпурные бактерии, пигменты розового цвета — микрококки.
Бактерии вида Pseudomonas aeruginosa (синегнойная палочка) выделяют растворимый в воде голубой феназиновый пигмент пиоцианин, который диффундирует в среду. Палочка Pseudomonasfluorescens синтезирует водорастворимый пигмент зеленого цвета — флюоресцин, поэтому при росте на плотной среде она образует флюоресцирующие желтовато-зеленые колонии.