МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 5. МЕТАБОЛІЗМ МІКРООРГАНІЗМІВ
Катаболізм аеробних хемолітотрофних мікроорганізмів
Літотрофні мікроорганізми використовують як джерело (донор) електронів неорганічні сполуки у відновленому стані. Залежно від донора електронів серед аеробних хемолітотрофів розрізняють:
- водневі бактерії (окиснюють молекулярний водень);
- нітрифікуючі бактерії (окиснюють амоній і нітрит);
- безбарвні сіркобактерії (окиснюють сульфід чи сірку);
- тіонові бактерії (окиснюють сульфід);
- карбоксидобактерії (окиснюють монооксид вуглецю);
- залізобактерії (окиснюють залізо-ІІ).
Метаболізм аеробних хемолітотрофів характеризується чотирма особливостями:
1. АТФ утворюється в процесі окисного фосфорилювання, тобто в дихальному ланцюзі.
2. Реакції субстратного фосфорилювання відсутні (виняток - тіонові бактерії).
3. Електрони надходять на дихальний ланцюг на рівні цитохромів (виняток - водневі бактерії).
4. Транспорт електронів по дихальному ланцюгу відбувається у прямому та зворотному напрямках.
Оскільки джерелом електронів у хемолітотрофів є неорганічні сполуки, їх окиснення відбувається без використання гліколітичного, Ентнера - Дудорова, пентозо-фосфатного шляхів та циклу Кребса. Відповідно реакції субстратного фосфорилювання, що є характерними для хемоорганотрофів, у цих мікроорганізмів відсутні. Електрони з окиснених неорганічних сполук безпосередньо подаються на дихальний ланцюг. Місце їх надходження визначається значенням ОВП реакції окиснення донора. У табл. 5.4 наведено значення ОВП основних реакцій хемолітотрофного метаболізму.
Таблиця 5.4. Окисно-відновні потенціали (ОВП) основних реакцій хемолітотрофного метаболізму
Реакції |
ОВП, мВ |
Н2→ 2Н+ + 2е- |
-410 |
НАДН + Н+ → НАД+ + 2е- + 2Н+ (реакція окиснення НАДН2 на дихальному ланцюзі при хемоорганотрофному метаболізмі) |
-320 |
H2S → S + 2Н++ 2е- |
-250 |
S + 3Н2О → SO32- + 6Н++ 4е- |
+5 |
SO32-+ Н2О → SO42- + 2Н++ 2е- |
-280 |
NH4++ 2Н2О → NO2- + 8Н++ 6е- |
+440 |
NO2- + H2O → NO3- + 2Н++ 2е- |
+350 |
O2 + 4Н++ 4е- → 2H2O (реакція відновлення кисню на дихальному ланцюзі при аеробному диханні) |
+810 |
Відірвавшись від донора, електрон подається на той переносник дихального ланцюга, значення ОВП якого вище, ніж ОВП реакції окиснення донора. Таким чином, у всіх хемолітотрофів, крім водневих бактерій, електрони потрапляють на дихальний ланцюг не на початок (рівень флавопротеїдів), а на середину (рівень цитохромів) (рис. 5.14).
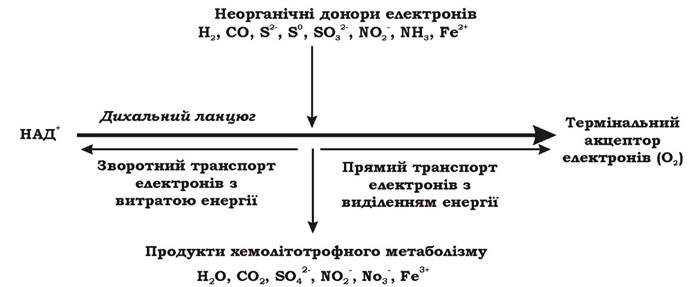
Рис. 5.14. Загальна схема хемолітотрофного метаболізму мікроорганізмів
У хемолітотрофних мікроорганізмів електрон може рухатися по дихальному ланцюгу у двох напрямках: прямому і зворотному. При прямому транспорті електрон прямує до термінального акцептора від переносників з нижчими значеннями ОВП до вищих. Таке перенесення електронів супроводжується утворенням градієнта протонів і, відповідно, синтезом АТФ. Оскільки шлях електронів у прямому напрямку в аеробних хемолітотрофів значно коротший, ніж у аеробних хемоорганотрофів, кількість пунктів фосфорилювання в них менше (як правило, один), а отже і вихід АТФ значно нижчий. При зворотному транспорті електрони від місця входження в дихальний ланцюг рухаються в напрямку флавопротеїдів (ФМН) і врешті відновлюють НАД+. Метою зворотного перенесення електронів є утворення відновлених еквівалентів, які необхідні клітині для біосинтезу і не можуть з'явитися іншим шляхом при хемолітотрофному метаболізмі. На відміну від хемоорганотрофів, в яких НАДН2 утворюється без витрат АТФ, у хемолітотрофів цей процес є енерговитратним. Енергія, необхідна для транспорту електронів по дихальному ланцюгу від переносників з вищими значеннями ОВП до нижчих при зворотному переносі, виділяється при гідролізі молекул АТФ, які попередньо синтезувалися при прямому перенесенні електронів. Так, Nitrobacter winogradskyi на синтез 1 моля НАДН2 витрачає 5 молів АТФ, а Thiobacillus - 2-3 молі АТФ.
Більшість хемолітотрофних бактерій є автотрофами. Асиміляція СО2 також потребує суттєвих енергетичних витрат. Так, для синтезу однієї молекули глюкози за циклом Кальвіна клітині необхідно використати 12 молекул відновлених еквівалентів та 18 молекул АТФ.
Таким чином, на відміну від хемоорганогетеротрофів, хемолітотрофи отримують АТФ лише за рахунок окисного фосфорилювання при прямому переносі електронів по короткій термінальній частині дихального ланцюга і додатково витрачають АТФ на автотрофний біосинтез та на відновлення НАД+ при зворотному переносі електронів.
Свої енергетичні проблеми хемолітотрофи вирішують таким чином:
1. Вони мають дуже розгалужену термінальну частину дихального ланцюга і високу концентрацію цитохромів. Це дозволяє їм передавати на термінальний акцептор велику кількість електронів. У зв'язку з цим більшість хемолітотрофів має розвинену систему внутрішніх цитоплазматичних мембран.
2. Окиснення неорганічних донорів електронів хемолітотрофами відбувається досить швидко. Вони переробляють велику кількість неорганічних сполук і виділяють у середовище багато кінцевих метаболітів, що є надзвичайно важливим для кругообігу речовин у природі.
Водневі (воднеокиснювальні) бактерії. Це аеробні хемолітоавтот- рофні прокаріоти, які здатні отримувати енергію та електрони шляхом окиснення молекулярного водню, а всі речовини клітини будувати з вуглецю СО2. Молекулярний водень використовується і в анаболізмі для отримання відновлених еквівалентів. Рівняння метаболізму водневих бактерій виглядають так:
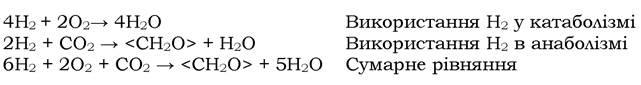
Водневі бактерії не є таксономічною групою. Це організми, яких об'єднують разом лише завдяки кільком спільним фізіологічним ознакам. До водневих бактерій належать представники 20 родів, які мають різну морфологію, різне забарвлення за Грамом, рухливі і нерухливі, спорові і неспорові форми. За винятком термофільних бактерій роду Hydrogenobacter, які характеризуються облігатною хемолітоавтотрофією, усі інші водневі бактерії - факультативні форми. Вони можуть використовувати як джерело вуглецю та енергії також різноманітні органічні сполуки, серед яких і одновуглецеві - оксид вуглецю, метанол, форміат.
Здатність окиснювати молекулярний водень пов'язана з наявністю гідрогеназ, що каталізують реакцію: Н2 → 2Н+ + 2е-. У водневих бактерій існує два типи цього ферменту (рис. 5.15):
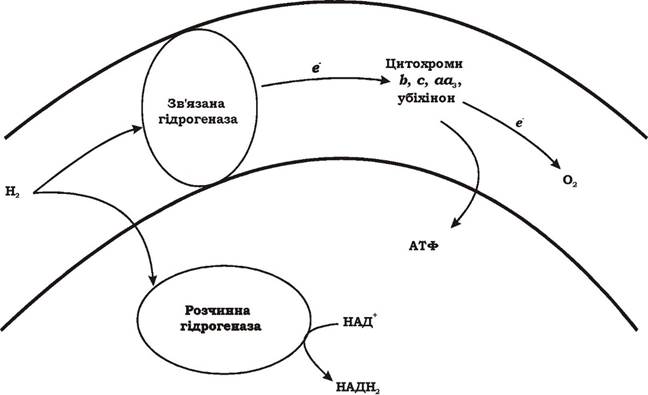
Рис. 5.15. Функції двох гідрогеназ у водневих бактерій
- розчинна гідрогеназа, яка міститься в цитоплазмі і каталізує реакцію відновлення НАД+. Продукт реакції НАДН2 у подальшому використовується для біосинтезу;
- гідрогеназа, яка зв'язана з субклітинними частками. Вона не здатна відновлювати НАД+ і безпосередньо передає електрони на дихальний ланцюг на рівень флавопротеінів, хінонів або цитохрома b. Тобто вона впливає лише на енергетичний метаболізм.
Більшість водневих бактерій містить лише одну форму гідрогенази - зв'язану з мембраною. Є види, що мають обидві форми або лише розчинну. Якщо водневі бактерії містять обидві форми гідрогенази, функції між ними чітко розмежовані. У разі відсутності розчинної гідрогенази виникає проблема отримання відновлених еквівалентів, яка вирішується шляхом зворотного перенесення електронів на НАД+. При функціонуванні лише розчинної гідрогенази вона виконує обидві функції: частина відновлених еквівалентів з НАДН2подається на дихальний ланцюг, інша використовується в конструктивному метаболізмі.
Таким чином, з усіх хемолітотрофних бактерій тільки водневі бактерії за допомогою певної форми гідрогенази можуть здійснювати пряме відновлення НАД+, окиснюючи неорганічний субстрат. З цим пов'язаний енергетичний вихід процесу: дихальний ланцюг має три або два пункти фосфорилювання.
Нітрифікуючі бактерії. Нітрифікація - це процес мікробного окиснення амонію до нітрату. Деякі хеоорганогетеротрофні мікроорганізми (FusaHum, Nocardia, Alcaligenes, Corynebacterium, Pseudomonas) можуть перетворювати амоній у нітрит за рахунок побічної діяльності ферментів. Таке окиснення не є джерелом енергії для клітини. У той же час існують дві вузькі фізіологічні групи аеробних бактерій, які здійснюють нітрифікацію з метою отримання енергії, тобто мають хемолітотрофний метаболізм. Саме такі бактерії називаються нітрифікуючими.
Процес нітрифікації має дві фази. Їх здійснюють різні фізіологічні групи бактерій за різними механізмами. Розрізняють першу і другу фази нітрифікації.
На першій фазі нітрифікації відбувається окиснення амонію (NH4+) до нітриту (NO2-), тобто N3- перетворюється в N3+. Цей процес проходить у два етапи:
1. Амоній → гідроксиламін (2NH4+ + O2 → 2NH4OH + 2H+)
Ця реакція енерговитратна (ΔG0' = +26,8 ккал), каталізується ферментом амоніймонооксигеназою (АМО) у присутності молекулярного кисню. Один атом молекулярного кисню відновлюється до води, а другий приєднується до амонію для утворення гідроксиламіну.
2. Гідроксиламін→ нітрит (NH2ОН + O2 → NO2 + H2О + Н+)
Гідроксиламін окиснюється до нітриту за допомогою гідроксиламі ноксидоредуктази. Ця реакція енергетично вигідна (ΔG0' = - 59,3 ккал), проходить через ряд нестабільних гіпотетичних продуктів (гіпонітрит, нітрогідроксиламін). Електрони, що вивільнюються при окисненні гідроксиламіну, подаються на дихальний ланцюг на рівень цитохрома с. Тобто істинним енергетичним субстратом першої фази нітрифікації є гідроксиламін.
Утворення гідроксиламіну пов'язане з високим значенням ОВП (+440 мВ) реакції NH4+ + 2H2O →NO2- + 8H+ + 6e-. У дихальному ланцюгу нітрифікаторів першої фази немає переносника, який міг би прийняти на себе електрон безпосередньо з амонію. Тому в бактерії виникає потреба спеціально, із витратою енергії, створювати проміжний продукт гідроксиламін, ОВП якого становить -40 мВ. Електрони, що утворюються при окисненні гідроксиламіну, приймає цитохром с, ОВП якого +245 мВ. На рис. 5.16 наведено схему окиснення амонію до нітриту в першій фазі нітрифікації.

Рис. 5.16. Схема окиснення амонію до нітриту на першій фазі нітрифікації: 1, 2 - етапи процесу; NH2OH -гідроксиламін; NOH - гіпотетичні проміжні продукти
Першу фазу нітрифікації здійснюють Nitrosomonas europea, Nitrosococcus oceanus, Nitrosococcus nitrosus, Nitrosolobus multiformis, Nitrosospira bnensis, Nitrosovibno tenuis.
На другій фазі нітрифікації відбувається окиснення нітриту (NO2-) до нітрату (NO3-), тобто N3+ перетворюється в N5+.
Цей процес проходить згідно з рівнянням: 2NO2- + O2 → 2NO3- і є енергетично вигідним (ΔG0' = - 36,4 ккал). Ключовий фермент нітри- фікаторів другої фази - нітритоксидоредуктаза. Вона зв'язана з мембраною. Електрони, що звільнилися при окисненні нітриту, подаються на дихальний ланцюг на рівень цитохрома аі. Другу фазу нітрифікації здійснюють Nitrobacter winogradskyi, Nitrospina gracilis, Nitrococcus mobilis, Nitrospira marina.
Для нітрифікуючих бактерій першої і другої фаз характерні незначний вихід АТФ, наявність зворотного переносу електронів, автотрофний анаболізм, потреба в кисні, вузький діапазон рН середовища. Оскільки нітрит є хімічно нестійкою сполукою і у природі не накопичується, нітрифікатори першої і другої фаз мешкають поруч.
Мікроорганізми, що окиснюють відновлені сполуки сірки. Бактерії здатні окиснювати такі сполуки сірки, як судьфіди (S2-,HS-), сульфіти (SO32-), тіосульфати (S2O32-), тритіонати (S3O62-), тетратіонати (S4O62-), елементарну сірку (S0), родоніди (CNS-), диметилсульфіди (CH3SCH3), диметилдисульфіди (CH3SSCH3), сульфідні руди. Їх можна поділити на три групи:
1. Бактерії, що мають хемолітотрофний метаболізм (тіонові бактерії).
2. Бактерії, що мають хемоорганотрофний метаболізм (безбарвні сіркобактерії).
3. Бактерії, що мають фототрофний метаболізм (пурпурні та зелені бактерії).
До групи тіонових бактерій належать роди Thiobacillus (T acidophilus, Т. albertis, T. aquaesulis, T. caldus, T. cuprinus, T. denitrifcans, T. ferrooxidans, T. halophilus, T. hydrotermalis, T. neapolitanus, T. novellus, T. plumbophilus, T. prosperus, T. tepidarius, T. thiooxidans, T. thioparus, T. thyasiris, T. versutus), Thiomicrospira (T. denitrificans, T. thermophila, T. pelophila, T. chilensis, T. frisia, T. crunogena, T. kuenenii, T. psychrophile, T. thyasirae), Paracoccus (стара назва Thiophaera) (P pantotropha), Thiodendron та ін. Це одноклітинні організми різної морфології, грамнегативні, спори не утворюють, аероби (Thiobacillus denitrificans може здійснювати нітратне дихання), мають облігатний чи факультативний хемолітотрофний тип метаболізму. Більшість тіонових бактерій - автотрофи (асимілюють СО2 за циклом Кальвіна), але серед них зустрічаються і гетеротрофи. У різних представників цієї групи, здатних рости хемоорганотрофно, виявлені ферменти гліколізу, пентозофосфатного шляху та шляху Ентнера - Дудорова. Описано функціонування цикла Кребса та гліоксилатного шунта. При хемолітотрофному метаболізмі тіонові бактерії отримують електрони при окисненні різноманітних сполук сірки (Thiobacillus ferrooxidans отримує енергію також і за рахунок окиснення заліза (ІІ)). Молекулярну сірку, як правило, вони не утворюють, але якщо вона все ж з'являється як проміжний продукт, то всередині клітини ніколи не накопичується.
Тіонові бактерії, на відміну від інших хемолітотрофів, при окисненні сполук сірки отримують АТФ в результаті як окисного, так і субстратного фосфорилювання.
При окисному фосфорилюванні вісім електронів, що поетапно відщеплюються від сульфіду, подаються на дихальний ланцюг на рівень цитохрома с. Термінальним акцептором електронів у більшості випадків є молекулярний кисень.
Субстратне фосфорилювання відбувається в процесі перетворення сульфіду в сульфат. Спочатку сульфід (S2-, HS-) окиснюється до сульфіту (SO32-). Можливо, це відбувається через стадію утворення елементарної сірки:
![]()
Потім, згідно зі схемою Пека (табл. 5.5), сульфіт активується АМФ і окиснюється до аденозин-5'-фосфосульфату (рис. 5.17) АФС- редуктазою. Електрони, що при цьому вивільнюються, спрямовуються на дихальний ланцюг.
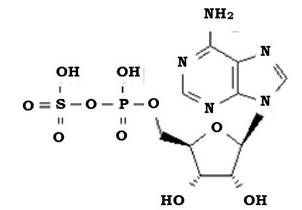
Рис. 5.17. Структурна формула аденозин-5'-фосфосульфату (АФС)
Таблиця 5.5. Схема субстратного фосфорилювання в тіобацил за Пеком
№ |
Рівняння |
Ферменти |
1. |
2SO32- + 2 АМФ → 2 АФС + 4е- |
АФС-редуктаза |
2. |
2 АФС + 2 Фн → 2 АДФ + 2SO42- |
АДФ-сульфурилаза |
3. |
2 АДФ → АМФ + АТФ |
Аденілаткіназа |
Позначення: АФС - аденозинфосфосульфат, Фн - фосфор неорганічний.
При подальшому фосфорилюванні АФС у середовище вивільнюється сульфат (SO42-), а перегрупування 2 АДФ призводить до синтезу АТФ. Оскільки ця молекула АТФ утворюється поза дихальним ланцюгом, її можна вважати результатом субстратного фосфорилювання.
НАДН2 тіонові бактерії отримують при зворотному перенесенні електронів на дихальному ланцюзі.
Безбарвні сіркобактерії дуже нагадують ціанобактерії, але не мають пігментів. Згідно з морфологією їх поділяють на дві групи: одноклітинні форми (роди Achromatium, Macromonas) та нитчасті форми (роди Thiothrix, Thioploca, Thiospinllopsis, Beggiatoa). Єдиною спільною ознакою цієї групи є те, що вони відкладають елементарну сірку в периплазматичному просторі клітин. Наразі достеменно не відомо, чи використовують ці бактерії відновлені сполуки сірки саме як джерело електронів у катаболізмі. Відомо, що вони мають низьку каталазну активність, а 80-90 % кисню, що поглинається при диханні, відновлюється лише до Н2О2. Для інактивації токсичного пероксиду водню цим організмам необхідна наявність у довкіллі невеликої кількості сульфідів. Відкладення елементарної сірки в периплазмі, таким чином, є результатом взаємодії Н2О2 та S2-. Здатність до автотрофного живлення в безколірних сіркобактерій також не доведено.
Пурпурні та зелені бактерії детально розглядатимуться нижче.
Карбоксидобактерії - це аероби, що здатні рости, використовуючи оксид вуглецю (СО) як єдине джерело електронів та вуглецю. Оскільки вони можуть також окиснювати молекулярний водень, деякі дослідники розглядають карбоксидобактерії як групу водневих бактерій. У той же час здатність використовувати як субстрат дихальну отруту вказує на здійснення карбоксидобактеріями нового типу хемолітотрофного метаболізму. На відміну від інших бактерій вони дуже стійкі до чадного газу і витримують його концентрацію до 20-95 % від об'єму.
Карбоксидобактерії окиснюють СО згідно з рівнянням:
![]()
Окиснення СО здійснюється за допомогою щонайменше одного специфічного ферменту - СО-оксидази, який міститься у клітині в розчинній та зв'язаній формах. Зв'язана СО-оксидаза локалізована з внутрішнього боку ЦПМ. При рості карбоксидобактерій на чадному газі СО-оксидаза виконує такі функції:
1. Окиснює СО до СО2.
2. Передає електрони на дихальний ланцюг.
3. Бере участь у синтезі НАДН2 шляхом зворотного переносу електронів.
Дихальний ланцюг карбоксидобактерій має один пункт фосфорилювання.
Вуглекислий газ, що утворився в результаті окиснення СО, служить джерелом вуглецю й асимілюється циклом Кальвіна. Тобто фактично джерелом вуглецю для карбоксидобактерій є не СО, а СО2. Сумарне рівняння використання СО є таким:
![]()
На рис. 5.18 наведено загальну схему асиміляції СО карбоксидобактеріями.

Рис. 5.18. Використання карбоксидобактеріями СО як джерела вуглецю та електронів
Однією з цікавих властивостей карбоксидобактерій є сам факт використання СО - інгібітора термінальних оксидаз, таких як цитохроми типу а. В електронотранспортних ланцюгах карбоксидобактерій не виявлено незвичайних цитохромів. Вважають, що можливі такі механізми їх резистентності до СО:
- швидка детоксикація СО за допомогою СО-оксидази;
- індукція синтезу СО-нечутливих термінальних оксидаз;
- підвищений синтез компонентів дихального ланцюга;
- просторове розмежування процесу окиснення СО та чутливих до нього цитохромоксидаз.
Карбоксидобактерії є факультативними хемолітоавтотрофами. Вони здатні рости і на органічних субстратах хемоорганогетеротрофно. Представники карбоксидобактерій: Pseudomonas carboxy do vorans, P. carboxydoflava, P. gazotropha, Seliberia carboxydohydrogena.
Фотосинтез (фотофосфорилювання) - це процес одержання організмами енергії за рахунок світла або це перетворення енергії світла в енергію хімічних зв'язків.
Сонячні промені, що досягають Землі, поділяються на три групи (рис. 5.19):
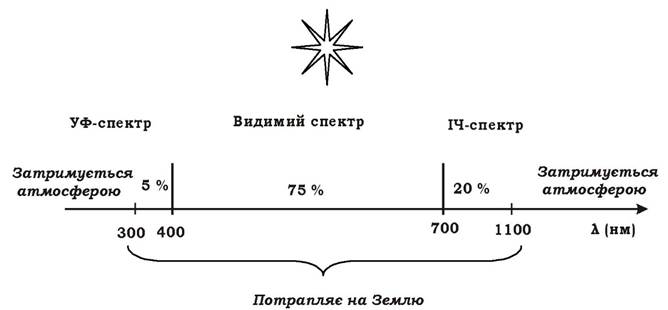
Рис. 5.19. Сонячні промені, що потрапляють на Землю
1. Ультрафіолетові (λ = 300-400 нм). На них припадає 5 % світла.
2. Видимого спектра (λ = 400-700 нм). Їхня частка становить 75 % світла.
3. Інфрачервоні (λ = 700-1100 нм). На них припадає 20 % світла.