МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 6. ОСОБЛИВОСТІ ФІЗІОЛОГІЇ АНАЕРОБНИХ МІКРООРГАНІЗМІВ
Метаногенез
Організми, які утворюють метан, належать до особливої групи живих істот, що називаються археями. Розглянемо їхні особливості детальніше.
Наприкінці 70-х років XX ст. на основі р-РНК філогенії було показано, що археї (Archaea), які раніше називали архебактеріями, є особливою групою живих організмів, що існують окремо від Bacteria (Eubacteria) та Eucana.
Археї належать до двох філогенетичних груп: Crenarchaea та Euryarchaea (рис. 6.34). Усі Crenarchaea - термофіли, що використовують у своєму метаболізмі сполуки сірки, а Euryarchaea складається з кількох термофілів (утилізують сірку), метаногенів та екстремальних галофітів.
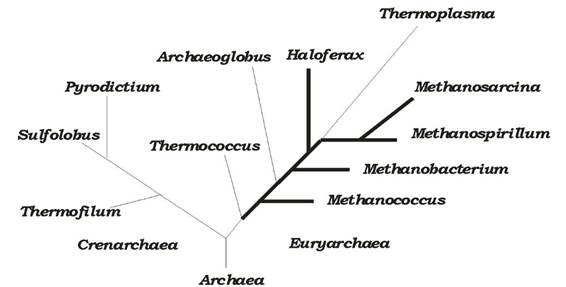
Рис. 6.34. Філогенія археїв (наведено деякі роди, жирними лініями показано галофіли та метаногени)
Археї демонструють величезне різноманіття способів існування і можливостей метаболізму. Багато видів адаптовані до екстремальних умов довкілля: високої солоності, температури, близької до точки кипіння води, екстремальних умов рН.
Фенотипово археї дуже нагадують бактерії. В основному це дрібні (0,5-5 мкм) палички, коки, спірили або нитчасті форми. Розмножуються вони поділом. Однак більшість археїв - термофіли, а багато й екстремальні термофіли. Вони часто мають автотрофний метаболізм чи дихають сіркою. Так само як і еукаріоти, археї мають велику кількість білків, що нагадують гістони, а також ДНК, складену у формі нуклеосом.
Метаногени (метаноархеї) - це група археїв, які у процесі своєї життєдіяльності утворюють метан (СН4). Метаногенез - єдиний шлях, за допомогою якого метаногенні археї можуть отримувати енергію для росту, і це єдині відомі організми, що здатні утворювати метан як продукт катаболізму.
Таксономічно всі метаногени належать до давнього царства Euryarchaeota. Вони поділяються на п'ять порядків: Methanobacteriales, Methanococcales, Methanomicrobiales, Methanosarcinales та Methano- pyrales. Серед них лише Methanosarcinales може зброджувати ацетат до СН4 і СО2 і рости на метанолі, метилтіолах та метиламінах як єдиних джерелах енергії. Гіпертермофільні види зустрічаються лише в порядках Methanobacteriales, Methanococcales і Methanopyrales.
Окрім особливостей у будові цитоплазматичної мембрани та клітинної стінки, притаманних усім археям, метаногени містять ще й унікальні коферменти, які беруть участь у перенесенні водню та С1-сполук при утворенні метану - кофермент F420, метанофуран, тетрагідрометаноптерин, кофермент М, фактор В, кофермент F430, метанофеназин.
Кофермент F420 - це похідна флавіну, що за структурою нагадує звичайний кофермент ФМН, але відрізняється будовою R-групи, не містить у середньому кільці атом азоту та на бензольному ядрі - метильної групи (рис. 6.35).
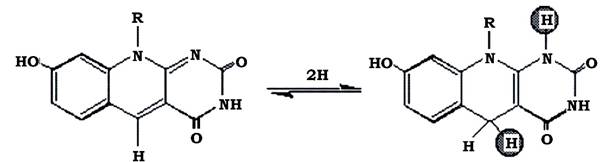
Рис. 6.35. Окиснена та відновлена форми фактора F420
Окисно-відновний потенціал F420 має досить низьке значення (Е0'= -380 мВ). Він взаємодіє з кількома різними ферментами метаногенів, у тому числі з гідрогеназою та НАДФ+-редуктазою. Окиснена форма F420 абсорбує світло при 420 нм і флуоресціює синьо-зеленим кольором, що дає змогу легко ідентифікувати метаногенів у флуоресцентному мікроскопі. Після відновлення F420 втрачає своє забарвлення. У клітині відновлений F420 виконує такі функції:
1. Переносить 2[Н] на НАДФ+ та фередоксин у реакціях біосинтезу, не пов'язаних з утворенням метану.
2. Переносить 2[Н] на сполуки вуглецю під час метаногенезу.
Метанофуран (MRF, MF) - низькомолекулярний кофермент, який є
першим акцептором СО2 при метаногенезі. Після першої реакції відновлення СО2 утворюється формілметанофуран. До складу метанофурану входять фенол, два залишки глутамінової кислоти, довгий ланцюг жирної кислоти і фуранове кільце (рис. 6.36).
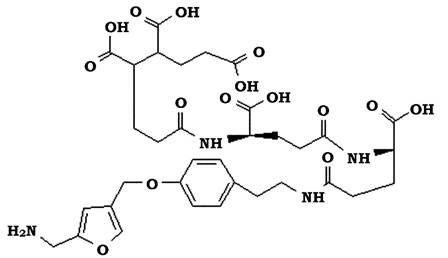
Рис. 6.36. Будова метанофурану
Тетрагідрометаноптерин (THMP, MP, H4MPT) - кофермент, що містить заміщене птеринове кільце, подібний до вітаміну фолієвої кислоти (рис. 6.37). Він є переносником С1-сполук протягом більшості етапів метаногенезу, починаючи з рівня формілу і до рівня метилу.
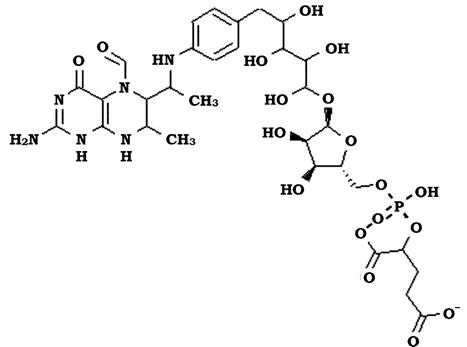
Рис. 6.37. Будова тетрагідрометаноптеріну
Кофермент М (2-меркаптоетансульфонова кислота, (H-S-KoM)) - переносник метильної групи. Він має дуже просту будову (рис. 6.38).
![]()
Рис. 6.38. Будова кофактора М
Кофермент М є дуже специфічним переносником у реакції відновлення метилу, яка відбувається через комплекс метилредуктаза- F30 на кінцевому етапі метаногенезу. Деякі мікроорганізми, зокрема Methanobrevibacter ruminantium, не можуть самі синтезувати цю сполуку, тому потребують її як фактор росту. Аналог коферменту М - брометаносульфонова кислота є сильним інгібітором метаногенезу і може використовуватися в експериментах для спрямованого пригнічення метаногенних археїв при вивченні анаеробних процесів. Це найменший з усіх відомих коферментів, знайдений лише в метаногенів.
Фактор В (кофермент В, H-S-KoB) - 7-меркаптогептаноїлтреонінфосфат (HTP-SH, HS-HTP) - це фосфорильована похідна амінокислоти треоніну, за будовою подібна до вітаміну - пантотенової кислоти (рис. 6.39).
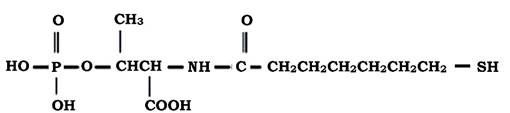
Рис. 6.39. Будова фактора В
Він так само, як і кофермент М, бере участь у заключних реакціях метаногенезу і також каталізує метилредуктазну систему, оскільки є для неї донором електронів. Фактор В вступає в реакцію з метил-КоМ і утворює дисульфід КоМ-S-S-HTP.
Кофермент F430 - розчинний тетрапірол, що містить нікель і має жовте забарвлення (рис. 6.40). Він абсорбує світло при 430 нм, але на відміну від F420 не флуоресціює. З наявністю F430 пов'язаний великий вміст нікелю в клітинах метаногенів. Цей кофермент має велике значення на заключному етапі метаногенезу, оскільки є частиною метил- редуктазної системи.
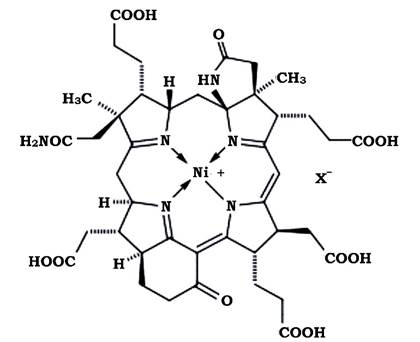
Рис. 6.40. Будова коферменту F430
Метанофеназин (МР) - це низькомолекулярний переносник електронів і водню, зв'язаний із мембраною (рис. 6.41). Він відновлюється F420-дегідрогеназою або гідрогеназою й, у свою чергу, відновлює гетеродисульфід на кінцевих етапах метаногенезу. Виявлений лише в метаногенів, що ростуть на метильованих сполуках.
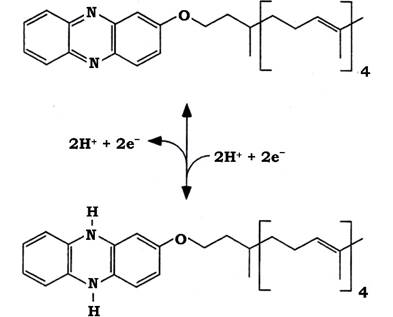
Рис. 6.41. Структура метанофеназину в окисненому та відновленому станах
Метаболізм метаногенів. Метаногени перетворюють у метан такі субстрати:
1. Речовини, які містять окиснений вуглець - вуглекислий газ СО2, форміат НСООН, монооксид вуглецю СО.
2. Речовини, що містять метильну групу - метанол СН3ОН, метиламін СН3NН3+, диметиламін (СН3)2NH2+, триметиламін (СН3)3NН+, метил- меркаптан СH3SH, диметилсульфід (СН3)2S.
3. Оцтову кислоту СН3СООН.
Ці субстрати перетворюються метаногенами з виділенням енергії згідно з рівняннями:
СО2 + 4Н2 → СН4 + 2Н2О ΔG0'= -134 кДж/реакція
4НСООН → СН4 + 3СО2 + 2Н2О ΔG0'= -281 кДж/реакція
4СО + 2Н2О → СН4 + 3СО2 ΔG0'= -209 кДж/реакція
4СН3ОН → 3СН4 + СО2 + 2Н2О ΔG0'= -323 кДж/реакція
СН3ОН + Н2 → СН4 + Н2О ΔG0'= -133 кДж/реакція
4СНзКНзОІ + 2НзО → зСН4 + СО2 + 4NH4Cl ΔG0'= -230 кДж/реакція
СН3СООН → СН4 + СО2 ΔG0'= -130 кДж/реакція
Метаногенез іноді розглядають як процес анаеробного дихання і називають його карбонатним диханням, оскільки при рості метаногенів на суміші СО2 та Н2 молекулярний водень є донором електронів, а вуглекислий газ - акцептором. Водночас у археїв відсутній традиційний
дихальний ланцюг з класичним набором переносників електронів. У літературі часто зустрічається термін "метанове бродіння". При рості метаногенів на метанолі одна молекула субстрату окиснюється, а друга - відновлюється, що нагадує процес збродження амінокислот. Однак відомо, що при метаногенезі АТФ синтезується за рахунок утворення градієнта протонів та іонів натрію, а це не є типовим для класичного бродіння. У метаногенів відсутнє субстратне фосфорилювання. Таким чином, метаногенез не можна розглядати ні як анаеробне дихання, ні як бродіння. Це унікальний специфічний процес отримання енергії, характерний лише для давніх форм живих організмів - археїв.
Утворення метану при рості метаногенів на суміші СО2 та Н (рис. 6.42). При утворення метану із суміші СО2 та Н2 відбувається вісім реакцій, наведених нижче:

Як видно з рівнянь, СО2 відновлюється поетапно, а С1-сполуки, що при цьому утворюються, постійно перебувають на одному з трьох переносників: метанофурані (MF), тетрагідрометаноптерині (Н4МРТ) та коферменті М (H-S-KoM).
Реакція 1. Першим акцептором СО2 є метанофуран (MF). Активований метанофураном вуглекислий газ відновлюється до рівня фор- мілу з утворенням формілметанофурану. Відновлення відбувається за рахунок водню, однак переносник водню в цій реакції досі чітко не визначений. Реакція утворення форміл-MF є енерговитратною.
Реакція 2. Формільна група переноситься з метанофурану на тетрагідрометаноптерін.
Реакції 3. Дегідратація формілтетрагідрометаноптеріну (CHO-H4МРТ) з утворенням метенілтетрагідрометаноптеріну (HC= Н4МРT).
Реакція 4. Відновлення метенілтетрагідрометаноптеріну (H2C=ЩМРT) до метилентетрагідрометаноптеріну (H2C=Н4МРT). Переносником водню в даній реакції є кофермент F420.
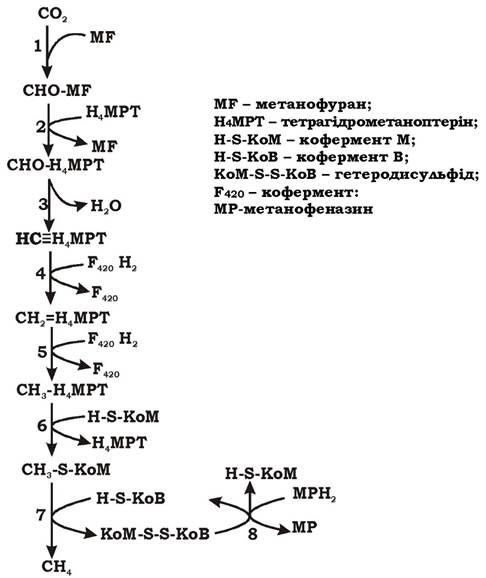
Рис. 6.42. Схема утворення метану при рості метаногенів на суміші Н2 та СО2
Реакція 5. Відновлення метилентетрагідрометаноптеріну (Н2С=Н4МРТ) до метилтетрагідрометаноптерину (Н3С-Н4МРТ). Переносником водню в даній реакції є кофермент F420.
Реакція 6. Метальна група переноситься на кофермент М. Процес супроводжується виділенням енергії (ΔG0' = -29,7 кДж/моль) та винесенням іонів Na+ у периплазму. Помпою для транспорту Na+ слугує фермент метил-Н4МРТ: КоМ-метилтрансфераза, що є частиною центрального шляху метаболізму метаногенів. Тому винесення Na+ у периплазму є обов'язковим при метаногенезі й приводить до утворення трансмембранного градієнта Na+. Метилтрансфераза містить кофактор (фактор ІІІ або Ко(ІІІ)), який бере участь у перетворенні метилу.
Кофактор Ко(ІП) у супервідновленій формі позначається як Ко(І). Механізм утворення градієнта іонів натрію (ΔμNa+) наведено на рис. 6.43.
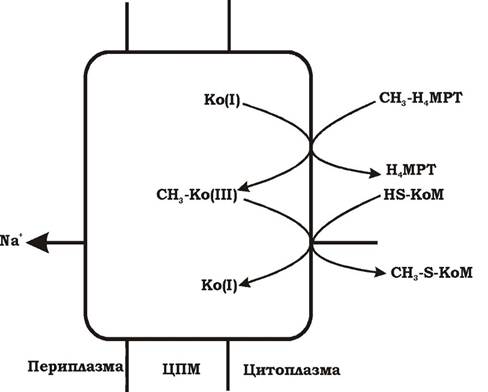
Рис. 6.43. Схема механізму транспорту іонів натрію за участю метил-Н4МРТ: КоМ-метилтрансферази
Перенесення метильної групи з Н4МРТ на КоМ відбувається у два етапи. На першому метильну групу приєднує Ко(І), відновлюючись при цьому до Ко(ІІІ) й утворюючи проміжний продукт СН3-Ко(ІII). На другому етапі СН3-Ко(ІІІ) передає групу СН3 на кофермент М, формується СН3-S-КоМ. Саме на другому етапі процесу відбувається транс- локація іонів натрію, пов'язана з реакцією деметилювання кофактора метилтрансферази.
Реакція 7. Метил-КоМ відновлюється до метану. Процес відновлення та деметилювання каталізується метил-КоМ-редуктазою і відбувається за участю електронів, які надходять із відновленого коферменту В, частиною метилредуктазної системи є кофактор F430. У результаті утворюється метан та гетеродисульфід КоМ та КоВ (КоМ-S-S-KoB).
![]()
Реакція 8. Гетеродисульфід КоМ-S-S-KoB відновлюється гетеродисульфідредуктазним комплексом ферментів. Електрони для його відновлення надходять із відновленого метанофеназину (МР) - зв'язаного з мембраною низькомолекулярного переносника.
![]()
Ця реакція необхідна для завершення циклу і є найбільш важливою для запасання енергії. Гетеросульфідредуктаза зв'язана з мембраною і функціонує як кінцевий фрагмент кількох електронотранспортних ланцюгів.
На рис. 6.44 показано зв'язок між реакціями 7 та 8.
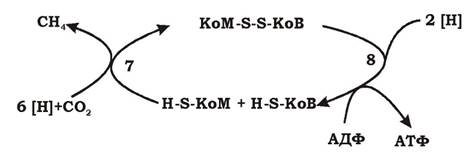
Рис. 6.44. Кінцеві реакції (7, 8) метаногенезу:
7 - реакція утворення метану; 8 - реакція відновлення гетеросульфіду КоМ-S-S-KoB
Кількість енергії, що виділяється в результаті відновлення метил- КоМ до СН4, є достатньою для винесення в периплазму протонів, тобто для створення градієнта протонів (ΔμΗ+).
Біосинтез органічних речовин у метаногенів, що ростуть на суміші СО2 та Н2, здійснюється за рахунок модифікованого шляху аце- тил-КоА (рис. 6.45).
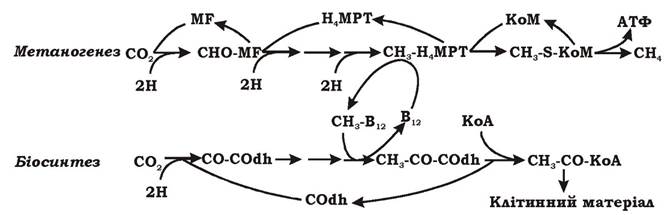
Рис. 6.45. Схема автотрофного метаболізму метаногенів:
MF - метанофуран; Н4МРТ - тетрагідрометаноптерін; KoM - кофермент М; COdh - СО-дегідрогеназа
При автотрофному рості на суміші газів СО2 та Н2 метаногени асимілюють СО2 модифікованим шляхом ацетил-КоА, кінцевим метаболітом якого є ацетил-КоА, що включається в біосинтез усіх необхідних клітинних компонентів. Оскільки метильна група ацетил-КоА утворюється при метаногенезі, цим мікроорганізмам не потрібно спеціально синтезувати її. Тому модифікований шлях ацетил-КоА має лише одну гілку, на якій СО2 відновлюється до карбонільної групи. Метильна ж група утворюється як інтермедіат при катаболізмі і після активації коферментом, що містить вітамін В12, включається у шлях ацетил-КоА.
Будова електронотранспортного ланцюга метаногенів. До ланцюга перенесення електронів у метаногенів можуть входити такі ферменти та переносники:
- Гідрогеназа - точка надходження в дихальний ланцюг електронів з молекулярного водню при використанні метаногенами Н2 як донора електронів.
- F420-дегідрогеназа - точка надходження в дихальний ланцюг електронів, що вивільнюються при окисненні F420H2 - універсального переносника водню в метаногенів.
- Гетеродисульфідредуктаза - фермент, що бере участь у відновленні гетеродисульфіду КоМ-S-S-KoB.
- Метанофеназин (МР) - низькомолекулярний переносник електронів і водню, зв'язаний з мембраною. Він відновлюється F42o-дегідро- геназою або гідрогеназою і, у свою чергу, відновлює гетеродисульфід КоМ-S-S-KoB.
- Цитохроми b- та с-типів.
За складом зв'язаних з мембраною переносників електронів і за механізмом утворення градієнта протонів метаногени можна поділити на дві групи:
1. Метилотрофні організми, наприклад Methanosarcina barken, M. mazei (містять різноманітні цитохроми b- та с-типів і переносник метанофеназин).
2. Метаногени, які споживають молекулярний водень, наприклад Methanobacterium thermoautotrophicum (не містять цитохромів, але містять поліфередоксини та аналоги флавопротеїнів).
Метаногени використовують різні механізми утворення градієнта протонів залежно від субстрату та складу переносників. На рис. 6.46 наведено можливі варіанти їхніх дихальних ланцюгів.
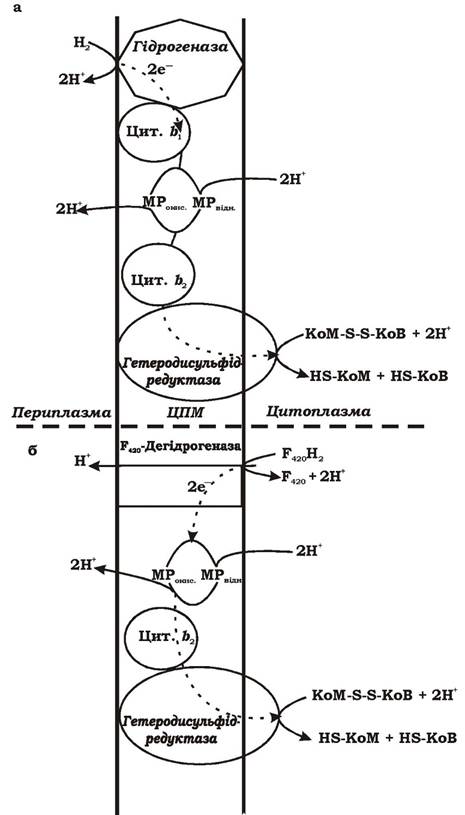
Рис. 6.46. Гіпотетична схема утворення градієнта протонів у метаногенів на етапі відновлення гетеродисульфіду КоМ-S-S-KoB при рості на різних субстратах: МР — Метанофеназин
Синтез АТФ у метаногенів. Метаногени - це організми, які створюють два градієнти іонів: протонів (ΔμΗ+) та натрію (ΔμNa+). Обидва ці градієнти беруть участь у синтезі АТФ. Протони повертаються у клітину через АТФ-синтетазу (Н+-АТФазу), що супроводжується синтезом АТФ. Щодо механізму синтезу АТФ завдяки градієнту іонів натрію існує дві гіпотези. Згідно з першою, у деяких метаногенів, наприклад у Methanobacterium thermoautotrophicum, крім Н+-АТФази є і Na+-AТФаза, яка забезпечує утворення АТФ при градієнті Na+ . Згідно з другою гіпотезою, іони натрію повертаються в клітину через Na+/H+ антипорт, що приводить до винесення протонів назовні. У цьому випадку синтез АТФ здійснюється лише за участю Н+-АТФази, а градієнт Na+ є додатковим джерелом енергії.
Утворення метану при рості метаногенів на метанолі. Метанол (СН3ОН) за ступенем окиснення вуглецю займає проміжне місце між СО2 та СН4, тому може як окиснюватися, так і відновлюватися. При окисненні метанолу до вуглекислого газу вивільнюються три пари електронів 3[2Н]. Анаеробне окиснення метильної групи - це термодинамічно невигідний процес, бо електрони від метильної групи (Е0'метанол/формальдегід = -180 мВ) мають надійти до переносника F420 (Е0'= -340 мВ), тобто при рості на метанолі метаногени мають додаткові витрати енергії. Для збалансування окисно-відновних процесів три пари електронів, що утворилися при окисненні метильної групи, мають відновити певний акцептор. Таким акцептором слугують інші молекули метанолу. На відновлення однієї молекули метанолу до метану витрачається одна пара електронів. Отже, в разі окиснення однієї молекули метанолу три молекули мають відновитися.
![]()
Реакція утворення метану при відновленні метильної групи пов'язана з утворенням градієнта протонів та синтезом АТФ. Схему метаболізму метаногенів при рості на метанолі наведено на рис. 6.47.
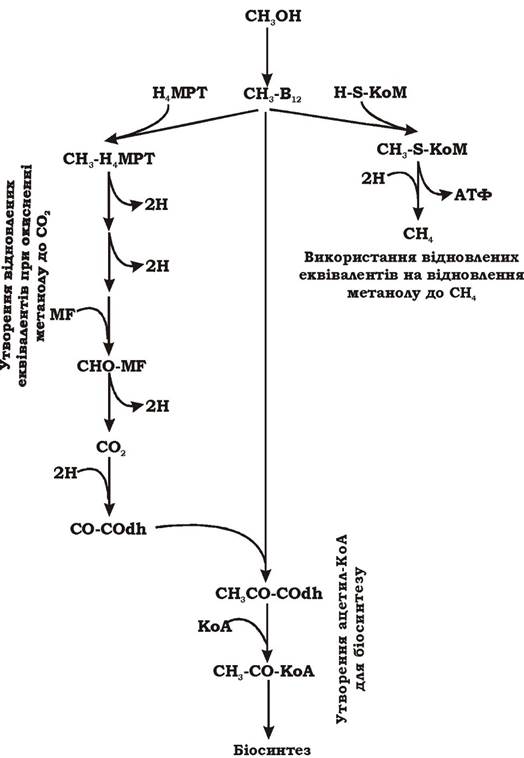
Рис. 6.47. Схема метаболізму метаногенів при рості на метанолі:
MF - метанофуран; Н4МРТ - тетрагідрометаноптерін; СОdh - СО-дегідрогеназа; H-S-KoM - кофермент М; В12 - кофермент з вітаміном В12
Як показано на рисунку, процес перетворення метанолу починається з активування метильної групи коферментом, що містить вітамін В12. Потім одна активована метильна група окиснюється до СО2. При цьому утворюється три пари відновлених еквівалентів і змінюється носій вуглецевої субстанції на Н4МРТ та MF (гілка зліва). Наступні кілька активованих метильних груп відновлюються до метану за участю відновлених еквівалентів, що утворилися в реакціях окиснен- ня метильної групи. Виділення метану супроводжується синтезом АТФ (гілка справа). Частково метильна група метанолу й СО2, що утворив-
ся в реакціях окиснення, за рахунок модифікованого шляху ацетил- КоА включаються в біосинтез.
Утворення метану при рості метаногенів на ацетаті. Ацетат розщеплюється метаногенами за допомогою великого комплексу ферментів на СО та метильну групу, зв'язану на різних етапах процесу з різними носіями. Потім СО окиснюється до СО2 за участю ферменту СО-дегідрогенази. Електрони, що при цьому вивільнюються, відновлюють метил-КоМ до метану з утворенням градієнта протонів і, врешті-решт, АТФ (рис. 6.48). Рівняння процесу має такий вигляд:
![]()
Оскільки продукти розщеплення ацетату є донорами (СО) й акцепторами (метил-КоМ) електронів, метаногенез на ацетаті нагадує бродіння. Однак АТФ у даному випадку утворюється не в результаті субстратного фосфорилювання, а за рахунок ΔμΗ+.
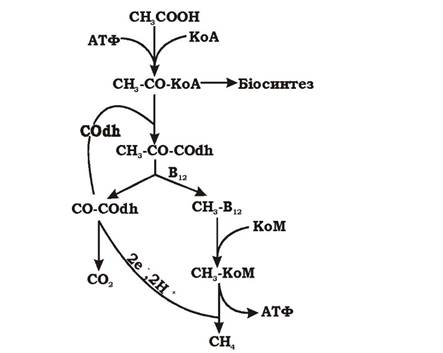
Рис. 6.48. Схема метаболізму метаногенів при рості на ацетаті: KoM - кофермент М; СОdh - СО-дегідрогеназа; В12 - кофермент з вітаміном В12