МІКРОБІОЛОГІЯ - М.Г. Сергійчук - 2008
Розділ 9. РОЛЬ МІКРООРГАНІЗМІВ У КРУГООБІГУ РЕЧОВИН У ПРИРОДІ
Деструкція мікроорганізмами природних речовин
Річна продукція органічної речовини на Землі становить приблизно 3,2 х 1011 т. Основну масу цієї речовини становлять сполуки рослинного походження, хімічний склад яких досить різноманітний: білки, амінокислоти, жири, вуглецевмісні сполуки (клітковина, лігнін, пектинові речовини, геміцелюлози тощо). Домінують за масою целюлоза, геміцелюлози, пектинові речовини і лігнін.
Целюлоза - це основна складова будь-якого рослинного матеріалу, а її синтез за своїми масштабами перевершує синтез всіх інших природних сполук. Рослинний опад містить 34-59 % клітковини. Про запаси целюлози у природі можна судити з таких даних: загальний вміст вуглецю у складі живих організмів оцінюється у 700 млрд т, крім того, у складі органічної речовини ґрунту і торфу визначається ще близько 1 400 млрд т вуглецю. Якщо врахувати, що з цієї маси вуглецю майже 50 % припадає на частку целюлози, тоді стає очевидним, яке значення має трансформація цієї речовини у природі.
Целюлоза належить до числа стійких сполук і у природних умовах розкладається виключно мікроорганізмами.
За хімічною природою це полісахарид, нерозчинна сполука, яка у природних умовах має зв'язки з іншими речовинами - геміцелюлозою, лігніном, таніном та ін. Молекули целюлози являють собою міцели (мікрофібрили), на поверхні яких адсорбуються інші сполуки. Мік- рофібрили агрегуються в пучки, покриті загальною оболонкою, які містять віск і пектин.
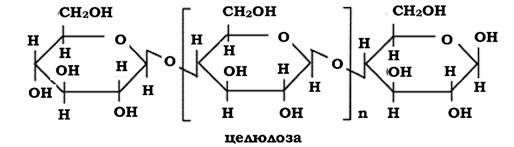
Елементарна одиниця лінійної полімерної молекули целюлози являє собою α-D-глюкозу із ступенем полімеризації ~ 14 000 і молекулярною масою ~ 500 000. Молекула не має певної довжини, а кількість глікозидних залишків може варіювати.
Лінійні молекули целюлози з'єднуються між собою водневими зв'язками, що визначає такі її властивості, як розчинність, гігроскопічність і реакційну здатність.
У зв'язку з великою кількістю целюлози, яка синтезується у природі, мікроорганізми, які її розкладають, відіграють особливо важливу роль у процесі її мінералізації та кругообігу вуглецю. Різноманітна мікрофлора забезпечує її трансформацію в різних умовах аерації, значеннях pH, температури та вологості. Целюлозу розкладають аеробні мікроорганізми (бактерії і гриби) і анаеробні мезо- та термофільні бактерії. Істинними целюлозоруйнівними організмами вважаються такі, які здатні впливати на нативну клітковину, а не на її похідні.
У добре аерованих ґрунтах целюлозу розкладають аеробні мікроорганізми (гриби, міксобактерії, еубактерії). Як і інші високомолекулярні сполуки, целюлоза не може асимілюватися в нативному стані - вона повинна попередньо поза клітиною гідролізуватися під впливом екзофер- ментів (деполімераз). У багатьох целюлозолітичних мікроорганізмів виявляються целюлази - ферменти, які розщеплюють целюлозу до глюкози або до дисахариду целобіози.
Важливе значення серед аеробних деструкторів целюлози займають представники роду Cytophaga (C. hutchinsonii), Sporocytophaga (S. mixococcoides), Sorangium (S. cellulosum), Archangium (A. gephyra) і Pseudomonas (P. fluorescens var. cellulosa). Але основна роль у розкладі целюлози в аеробних умовах належить грибам Fusarium і Chaetomium. Розкладають целюлозу також Aspergillus fumigatus, A. nidulans, Botrytis cinerea, Rhizoctonia solani, Trichoderma viride, Chetomium globosus і Myrothcium verrucaria.
Бактерії роду Cytophaga - це грамнегативні палички (0,3-0,8 х 1,515,0 мкм) із заокругленими або дещо звуженими кінцями, які здатні до ковзного руху. Клітинна маса зафарбована в жовтий, оранжевий або червоний колір за рахунок клітинних каротиноїдів або флексирубіноподібних пігментів. Облігатні аероби або факультативні анаероби. Хемоорганотрофи з дихальним або бродильним типом метаболізму. Усі представники здатні розкладати целюлозу, агар, хітин, пектин і крохмаль. Оптимальна температура росту 20-35° С. Оптимальне значення pH близько 7,0. Виявляються у ґрунтах, прісних та морських середовищах.
Гнучкі, грамнегативні із заокругленими кінцями палички (0,3-0,5 х 5,0-8,0 мкм), які здатні при старінні утворювати сферопласти і деформовані клітини та мікроцисти як форми спокою, віднесені до роду Sporocytophaga. Вони проявляють здатність до ковзного руху. Хемоорганотрофи з дихальним типом метаболізму. Облігатні аероби. Як джерело енергії і вуглецю використовують целюлозу, целобіозу, глюкозу. Агар і хітин не розкладають. Оптимальна температура росту коливається в межах 30 оС.
Представники роду Sorangium мають клітини паличкоподібної форми (0,9-1,2 х 2,5-6,0 мкм), здатні утворювати міксоспори - світлозаломлюючі циліндричні палички із заокругленими кінцями. Плодове тіло складається з дуже дрібних сферичних або багатогранних щільно упакованих спорангіолей, які утворюють пакети. Пакети збираються у великі скупчення, які утворюють суцільний щільний шар. Плодові тіла зафарбовані в жовтий, оранжевий або оранжево-коричневий колір. Усі штами розкладають целюлозу, а деякі - хітин.
В анаеробних умовах целюлоза трансформується переважно мезофільними і термофільними бактеріями роду Clostridium з утворенням різних кінцевих продуктів (табл. 9.3).
Таблиця 9.3. Продукти розкладу целюлози деякими целюлозолітичними клостридіями
Вид |
Продукти розкладу |
Мезофіли |
|
C. omelianskii |
Етиловий спирт, оцтова, молочна і мурашина кислоти, СО2, Н2 |
C. dissolvens |
Етиловий спирт, оцтова, молочна і масляна кислоти, СО2, Н2 |
C. cellobioparum |
Етиловий спирт, оцтова, мурашина і молочна кислоти, СО2, Н2 |
Термофіли |
|
C. thermocellum |
Етиловий спирт, оцтова, молочна і мурашина кислоти, СО2, Н2 |
Целюлозолітичні бактерії - це важливі складові мікробіоценозу рубця жуйних тварин. Основними джерелами вуглеводів для жуйних тварин є сіно, солома і трава. Вміст целюлози в сіні становить ~ 50 %. Целюлозні компоненти кормів були б недоступними для них, якби в процесі еволюції не створилися симбіотичні взаємовідносини з мікроорганізмами, які здатні гідролізувати целюлозу. Кількість целюлозолітичних бактерій в рубці становить 109-1010 клітин/мл.
У процесі ферментації в рубці жуйних близько 90 % целюлози трансформується в кислоти: оцтову (50-70 %), пропіонову (17-21 %), масляну (14-20 %). При цьому виділяється до 700 л за добу газів (СО2 і СН4). Молочна кислота в рубці не накопичується, вона під впливом інших мешканців трансформується в пропіонову та оцтову кислоти.
Бактерії рубця відіграють багатогранну роль у живленні жуйних: кислоти, які утворюються при розкладі полісахаридів, всмоктуються; самі бактерії при проходженні вмісту рубця в кишечник перетравлюються, а їхня клітинна речовина засвоюється твариною; бактерії рубця асимілюють і неорганічні джерела азоту, що суттєво збільшує кількість білка, який отримує організм тварини.
Розклад целюлози мікроорганізмами проходить у декілька етапів. Спочатку відбувається ферментативний гідроліз полімеру. Цей процес йде за участю целюлази, яка являє собою комплекс двох ферментів: ендо- і екзоглюканази (С1-фактор і Сх-фермент). С1-фактор (целюлобіогідролаза) розриває α-1,4-зв'язки, тобто гідролізує високоорганізовану кристалічну структуру целюлози до великих фрагментів з вільними кінцями. Це екстрацелюлярний фермент глікопротеїн із співвідношенням вуглеводів і білків 1 : 1. Сх-фермент (екзо-α-1,4-глюконаза) відщеплює на кінці ланцюга дисахарид целобіозу. Можливе відщеплення целотріоз, целотетроз. Сх-фермент - це група неоднорідних білків. Целобіоза під впливом α-глюкозидази гідролізується до глюкози, яка зазнає подальших метаболічних трансформацій:
![]()
Прості цукри, які містяться в рослинних опадах (моно- і дисахариди), а також низькомолекулярні полісахариди (крохмаль, інулін, та ін.) досить легко розкладаються різними групами мікроорганізмів.
Пектинові речовини - це високомолекулярні сполуки вуглеводної природи - полігалактуроніди (CHO(CHOH)4COOH), які складаються із залишків нерозгалужених a-D-галактуронових кислот, з'єднаних між собою α-1,4-глюкозидними зв'язками. Карбоксильні групи кислот повністю або частково етерифіковані метанолом.
Розрізняють три типи пектинових речовин: протопектин - водо- нерозчинна складова клітинної стінки; пектин - міжклітинний водорозчинний полімер галактуронової кислоти, який має метилефірні зв'язки; пектинова кислота - водорозчинний полімер галактуронової кислоти, в структурі якого метилефірні зв'язки не виявляються.
У рослині пектинові речовини виявляються у вигляді нерозчинного протопектину - комплексної сполуки пектину зі складними речовинами клітинної оболонки: целюлозою, геміцелюлозою, арабаном і ксиланом.
Розчинний пектин - це полісахарид, який складається із з'єднаних між собою залишків галактуронової кислоти, що входить до складу пектину у формі метилового ефіру. Залежно від походження молекулярна маса пектину коливається від 25 000 до 50 000. Протопектин і пектин містяться у клітинних оболонках стебла, плодів, кореня і насіння багатьох рослин, утворюючи в комплексі з іншими речовинами субстрат, який склеює клітини.
Мікроорганізми, які проявляють здатність до деструкції пектинових речовин, синтезують три групи екзоферментів: протопектиназу - фермент, який каталізує гідроліз протопектину до розчинного пектину; пектинестеразу, яка гідролізує метилефірний зв'язок пектину з утворенням пектинової кислоти і метилового спирту; пектиназу - фермент, який розриває зв'язки між одиницями галактуронової кислоти, пектину або пектинової кислоти з утворенням невеликих ланцюжків, і в кінцевому рахунку - D-галактуронової кислоти.
Здатність до розкладу пектинових речовин властива багатьом грибам і бактеріям, які розкладають ці сполуки в аеробних і анаеробних умовах. В 1 г ґрунту визначається ~105клітин мікроорганізмів, здатних до деструкції пектинових речовин. Фітопатогенність різних мікроорганізмів (Botrytis cinerea, Fusarium oxysporum) базується на виділенні ферментів, які руйнують пектини. Erwinia carotovora викликає розпад тканин (мокрі гнилі) у багатьох рослин (салат, морква, селера та ін.). Найактивнішими деструкторами пектинових речовин є Bacillusma.cerans і B. polymyxa. Таку здатність проявляють й інші бактерії: B. subtilis, Clostridium pectinovorum, C. felsineum, C. pectinolyticum, C. flavum.
Пектинолітичні ферменти синтезують гриби родів Cladosporium, Alternaria, Penicillium, Fusarium, Sclerotinia та ін.
Розпад пектинової кислоти можна подати таким чином:
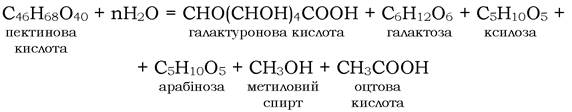
Сполуки, які утворилися в процесі розкладу пектинової кислоти, трансформуються в аеробних або анаеробних умовах. В анаеробних умовах в основному трансформується галактоза й арабіноза. Трансформація цих речовин відбувається за типом маслянокислого бродіння, яке викликається представниками роду Clostridium (C. pectinovorom, C. felsineum). Якщо трансформація забезпечується C. pectinovorom, то процес іде з утворенням масляної та оцтової кислот, а також з виділенням СО2 і Н2:
![]()
Бродіння, яке викликається C. felsineum, завершується утворенням, крім названих сполук, деякої кількості ацетону (СН3СОСН3) і бутанолу (СН3(СН2)2СН2ОН).
Продукти, які утворилися в процесі розкладу пектинових речовин, можуть окиснюватися до СО2 і Н2О. Найактивніше цей процес іде під впливом грибів (Cladosporium, Alternaria, Rhodotorula) та деяких бактерій (B. subtilis). Активніше окиснюється галактоза та оцтова кислота, дещо повільніше ксилоза, арабіноза і галактуронова кислота.
Розклад пектинових речовин має і позитивне значення, оскільки ці процеси відбуваються при розкладі рослинного опаду, освітленні фруктових соків при їх промисловому виробництві, мочінні льону і конопель.
Мета мочіння льону - відділення пучків целюлозних волокон від інших рослинних тканин. Найбільш давнім методом є росяне мочіння, за якого лубоволокнисті рослини розстеляють восени на траві. Такий метод дає непогані результати, але має й недоліки, а саме: процес триває 3-8 тижнів; може поширюватись і на целюлозу, тобто порушити цілісність (якість) волокна.
Останніми роками мочіння льону проводять на льонозаводах у спеціальних чанах при певній температурі, яка визначається культурою мікроорганізмів. Процес триває 3-5 діб. Непогано зарекомендував себе препарат пектолітин, який являє собою спори Clostddium felsineum.
Геміцелюлози - це гетерополісахариди (цукри й уронові кислоти), які визначаються в усіх рослинних тканинах. Вони входять до складу міжклітинної речовини. Солома і луб містять до 20 % геміцелюлози, макуха цукрового очерету 30 %, деревина хвойних дерев - 7-12 %, деревина листяних порід - 20-25 %.
За хімічною природою це полімер a-D-ксилози, з'єднаної 1,4-глікозидними зв'язками. Тобто ксилани відрізняються від молекул целюлози заміною груп СН2ОН на атоми водню. Деякі геміцелюлози містять арабінозу, глюкозу, галактозу і глюкуронову кислоту. Останніми роками замість терміна геміцелюлоза частіше вживається термін ксилан.
Ксилан розкладається швидше, ніж целюлоза. У його розкладі беруть участь різні групи мікроорганізмів, завдяки їхній здатності синтезувати ксиланазу. Деякі з них синтезують як ксиланазу, так і целюлазу. Найпоширенішими з них є бактерії роду Sporocytophaga, гриби родів Aspergillus, Rhizopus та Fomes. У лужних ґрунтах виявляються представники родів Vibdo, Bacillus, Clostridium, які теж здатні до деструкції ксилану.
У деяких бактерій (Clostridium) ксиланаза - конститутивний фермент, в інших вона утворюється в результаті індукції ксиланом.
Лігнін (лат. lignum - деревина) - найпоширеніша у природі полімерна циклічна сполука, яка міститься у вторинних шарах клітинної оболонки і як один із компонентів входить до складу міжклітинної речовини. Вміст цього полімеру збільшується при дозріванні рослини. Молоді трави містять ~ 6 % лігніну (за сухою масою), а у здерев'янілих тканинах рослин - від 18 до 30 %. Молекулярна маса лігніну коливається від 1 000 до 10 000. Ця сполука нерозчинна у воді і більшості органічних розчинників. Молекула лігніну містить лише три елементи - С, О і Н. Це тривимірний полімер фенольної природи, при окисненні якого утворюються альдегіди. Лігнін досить стійка сполука, він розкладається повільніше, ніж целюлоза і геміцелюлоза.
За хімічною структурою сполука неоднорідна. Лігнін, який міститься в рослинах різних видів, родів і родин рослинного царства хімічно теж відрізняється. Усі мономерні блоки в молекулі лігніну - це похідні фенілпропану, насамперед - коніфериловий спирт. Складність будови лігніну зумовлена різноманітністю зв'язків, за допомогою яких мономери з'єднуються між собою.
Лігнін хвойних порід складається переважно із залишків коніферилового спирту, лігнін листяних порід містить коніфериловий і синаповий спирти, а лігнін злакових, крім перелічених, - ще й кумаровий спирт. Відмінності спостерігаються, перш за все, у метоксильних групах, які визначаються у структурі лігніну.
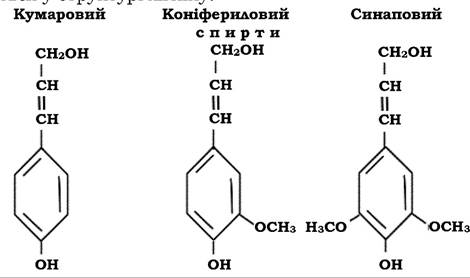
Характерною особливістю лігніну є те, що окремі його мономери зв'язані між собою не однією формою зв'язку, а різними - виявляються ефірні і С-С зв'язки, які надзвичайно стійкі до дії ферментів. В організмі рослин лігнін - це інертна сполука, яка виконує лише механічні функції. Здатність до деструкції цього полімеру проявляють лише мікроорганізми.
Більшість мікроорганізмів, діючи на лігнін, викликають незначні зміни, які проявляються у зменшенні кількості метоксильних груп. Але деякі сумчасті та недосконалі гриби можуть рости на субстратах, де лігнін є єдиним джерелом вуглецю (Fusarium lactis, F. nivale).
В аеробній деструкції лігніну беруть участь різні представники класу Basidiomycetes. Так, при помірній температурі лігнін розкладають виші гриби родів Clavaria, Armillariella, Fomes, Polistictuis, Polyporus, Ustilina. Активні щодо лігніну також гриби Trichoderma lignorum, Alternaria tenuis, Stremphylium botryosum. Бактерії роду Pseudomonas беруть участь у термофільній деструкції лігніну. В анаеробних умовах лігнін розкладається бактеріями роду Clostridium. Вважається, що первинна дія при деструкції лігніну здійснюється поліфенолоксидазами.
У ґрунті постійно відбуваються різні процеси розкладу природних сполук, які супроводжуються не лише утворенням кінцевих продуктів типу СО2, води, аміаку тощо, але й темно-коричневих продуктів, які отримали назву гумінових або перегнійних речовин. Ці сполуки містяться у ґрунті у великих кількостях і визначають його агрохімічні властивості.
Органічна речовина ґрунту існує у вигляді двох фракцій: великих частинок (неактивна фракція) і дрібних колоїдних частинок, до яких належить гумус. Гумус виконує такі функції:
- цементуючи грудочки ґрунту, він сприяє його структуризації;
- у гумусі накопичуються речовини, які після їх мінералізації використовуються рослинами;
- гумус має високу гідроємність, чим сприяє збереженню вологи у ґрунті і покращує його теплові властивості;
- у гумусі містяться біологічно активні речовини, які стимулюють ріст як рослин, так і мікроорганізмів;
- гумус покращує активність мікробіологічних процесів;
- відіграє важливу роль у циклі перетворення азоту та інших елементів. Гумус відрізняється за якістю та кількісним вмістом у ґрунтах різних
типів. Це визначається тим, що гумус, по-перше, утворюється з рослинних решток неоднакового хімічного складу і в різних кліматичних умовах, і, по-друге, в його утворенні беруть участь організми, специфічні для конкретної природної зони.
Хімічну природу гумусу остаточно не встановлено. Під цим терміном об'єднують комплекс високомолекулярних сполук, до яких входять гуміни, гумінові та гіматомеланові кислоти.
Сполуки гумусу відрізняються між собою деякими властивостями: кольором; вмістом С, Н, О та N; ступенем полімеризації; дисперсністю (табл. 9.4). Наприклад, гуміни і гумінові кислоти мають темно-буре забарвлення, фульвокислоти - жовте, а гіматомеланові - шоколадно- коричневе. Деякі гумусові сполуки розчинні в лугах (гумусові кислоти, фульвокислоти). Фульвокислоти розчинні також у воді, чим пояснюється їхня здатність мігрувати у глибокі шари ґрунту.
Процес утворення гумусу до кінця не вивчений. Але не викликає сумніву, що він утворюється у процесах синтетичних реакцій. На синтетичний процес утворення гумусу вказував ще в 1814 р. А. Трусов. В. Вільямс, а в подальшому С. Ваксман сформулювали теорію про лігно- протеїновий комплекс як ядро гумусу. У процесі гуміфікації лігнін розщеплюється, вивільнюються його структурні одиниці, які служать матеріалом для формування гумусу. До складу гумусу входять також білкові компоненти, жири, вуглеводи, тобто сполуки, які є продуктами життєдіяльності мікроорганізмів. Група мікроорганізмів, здатних розкладати гумус, була названа С. Виноградським автохтонною.
Таблиця 9.4. Склад та основні характеристики перегнійних сполук
Перегнійні сполуки |
Колір |
Розчинність |
Вміст, % |
|||
С |
Н |
О |
N |
|||
Гуміни |
Темно-бурий |
Нерозчинні в лугах |
52,0-63,0 |
3,5-5,6 |
30,2-38,4 |
3,5-5,9 |
Гумусові кислоти |
Темно-бурий |
Розчинні в лугах, кислотами осаджуються, нерозчинні у спиртах |
52,0-62,0 |
3,5-5,0 |
30,0-33,0 |
3,5-5,0 |
Фульвокис лоти |
Жовтий |
Розчинні у воді і лугах, не осаджуються кислотами |
44,0-49,0 |
3,5-5,0 |
44,0-49,0 |
2,0-4,0 |
Гіматоме ланові кислоти |
Шоколадно-коричневий |
Розчинні в лугах і спирті, осаджуються кислотами |
~60 |
~ 4,4 |
1,0-1,4 |
|
Гумус у різних ґрунтах накопичується і розкладається з різними швидкостями. Наприклад, у кислих, слабо окультурених дерново- підзолистих ґрунтах синтез гумусу домінує над його розкладом. У таких умовах рослинам не вистачає поживних речовин. Для прискорення процесів розкладу і встановлення рівноваги проводять вапнування ґрунту. У сіроземах, навпаки, гумус, що утворюється, швидко розкладається і не відбувається його накопичення. Для прискорення й нормалізації процесів гуміфікації рекомендується вводити в сівозміну бобові рослини. Вони мають довге коріння, яке здатне поглинати кальцій із глибоких шарів ґрунту й акумулювати його в кореневій шийці. При розкладі таких рослинних решток утворюється активний (діяльний) гумус з великим вмістом кальцію, який і надає ґрунту грудкуватої структури, склеюючи ґрунтові частинки, що має значення в його оструктуренні.
Структура ґрунту є важливою його характеристикою. Структурний ґрунт при оранці легко розпадається на грудочки. У такому ґрунті міститься багато некапілярних проміжків. Дощова вода легко поглинається таким ґрунтом і розсмоктується між капілярами. Середні й великі пори швидко звільнюються від води, в них формується запас ґрунтового повітря. Усе це забезпечує встановлення сприятливого співвідношення води і повітря у ґрунті. Структурні ґрунти легше обробляти, в них рідше виникають ерозійні процеси.
У безструктурних ґрунтах дрібні частинки залягають суцільною масою на всю глибину орного шару. Вони погано всмоктують дощові опади, велика кількість води стікає з полів. Волога і повітря в безструктурних ґрунтах - антагоністи. На поверхні ґрунту формується пробка, яка погіршує його аерацію.
Паралельно із процесами гуміфікації рослинних решток відбувається збагачення їх азотом, про що свідчить співвідношення C: N у рослинних рештках (40 : 1) і в гумусі (9 : 1).