ФАРМАЦЕВТИЧЕСКАЯ МИКРОБИОЛОГИЯ - В. А. Галынкин - 2015
ЧАСТЬ I. БИОЛОГИЯ МИКРООРГАНИЗМОВ
ГЛАВА 2. ГРИБЫ
2.1 Положение грибов в системе живой природы
Грибы — многочисленная широко распространенная в природе группа организмов, включающая в себя около 100 тыс. видов. Они обитают в почве, воде, на растительных и животных остатках [9].
Грибы составляют особое царство Fungi (Mycota), они обладают признаками, характерными как для растений, так и для животных. Грибы (Fungi) (рис. 19) — бесхлорофильные низшие эукариотические организмы, использующие для питания только органические вещества. Вегетативное тело грибов — грибница, или мицелий, состоит из ветвящихся нитей, называемых гифами. У низших одноклеточных грибов гифы не разделены поперечными перегородками (септами) на отдельные участки. У высших грибов мицелий септирован. Мицелий может развиваться на поверхности субстрата, в толще субстрата, проникая внутрь. Толщина (поперечник) гиф составляет 5.. .50 мкм и более. Своеобразие грибов проявляется не только в сочетании признаков, присущих растениям и животным, но и в наличии специфических черт и свойств, характерных только для членов царства Mycota: мицелиальной структуры вегетативного тела; сложных ядерных циклов и плеоморфизма; многоядерности и гетерокариоза (разнокачественность ядер в одной клетке); дикариоза (длительное существование в одной клетке двух ядер, одновременно делящихся и имитирующих диплоидное ядро).
Рис. 19. Морфология грибов. Вегетативные структуры: столоны (1) и ризоиды (2) Mucor nigricans; оидии (3) и хламидоспоры (4) Mucor chibinensis; псевдомицелий (5) и бластоконидии (6) Candida spp.; почкующиеся клетки (7); Saccharomyces cerevisiae. Анаформы: спорангий со спорами (8), спорангиеносец (9), конидиеносец (10), стеригмы (11), конидии (12); Aspergillus spp. (А) и Penicillium spp. (B). Телеоформы: сумки c аскоспорами (13) S. Cerevisiae (Endomycetes), перитеций, содержащий сумки с аскоспорами (14) Sordaria sp. (Ascomycetes), зигоспора (15) Mucor spp. (Zygomycetes).
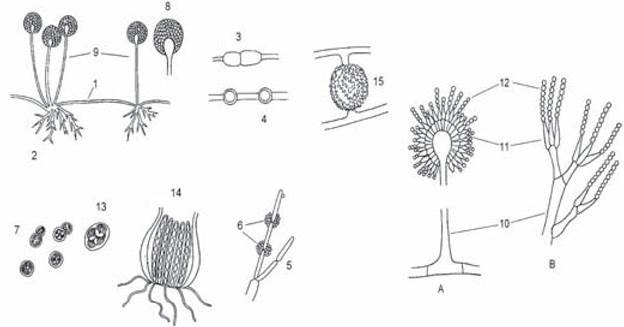
Признаки растений: голофитный способ питания (поглощение питательных веществ через клеточную стенку), способность к синтезу витаминов, наличие ригидной клеточной стенки, вакуолей, поперечных перегородок в мицелии, полярность клетки, способность к неограниченному росту, размножение спорами. Признаки животных: отсутствие хлорофилла, гетеротрофный тип питания, образование мочевины в ходе азотистого обмена и гликогена — в процессе углеводного обмена, наличие хитина в клеточной стенке, формирование лизосом в цитоплазме, особенности первичной структуры цитохромов и тРНК. Классификация грибов, построенная на основании структурно-химических и морфологических признаков представлена в табл. 3. Истинные грибы разделяют на шесть классов: хитридиомицеты, оомицеты, зигомицеты, аскомицеты, или сумчатые, базидиомицеты, дейтеромицеты, или несовершенные грибы.
Хитридиомицеты. Примитивные низшие одноклеточные организмы. Мицелий отсутствует или в зачаточном состоянии. Клеточная оболочка содержит хитин и не имеет целлюлозы. Размножение бесполое и половое. Преимущественно обитают в водоемах. Некоторые виды вызывают болезни сельскохозяйственных растений.
Оомицеты. Одноклеточные (низшие) мицелиальные грибы. Функцию скелетного вещества оболочки выполняют целлюлоза и глюкан. Размножение бесполое. Обитают в водоемах; наземные формы — паразиты высших растений.
Зигомицеты. Мицелий хорошо развит, многоядерный, несептированный (низшие грибы). Клеточная оболочка содержит хитин, иногда глюкан. Размножаются спорангиоспорами, реже конидиями или половым путем. Широко распространены в верхнем слое почвы, развиваются на органических остатках растений. Используют в микробиологической промышленности с целью получения соевого сыра, спирта из картофеля, антибиотика рамицина и др.
Таблица 3. Классы грибов [10].
(Г + Ц) % |
Клеточная |
стенка |
Ядерная фаза |
Ана-морфа |
Телео-морфа |
|||
Порядок |
Класс |
Биосинтез триптофана |
Хитин, % |
Маннан, % |
||||
Chytridiomiycota |
Chytridiomycetes |
44 — 46 |
Ia |
>5 |
<5 |
Гаплоидная или гаплодиплоидная |
Зооспорангий с зооспорами |
Покоящаяся спора, первичный спорангий |
Zygomycota |
Zygomycetes |
28 — 54 |
Ib |
>5 |
<5 |
Гаплоидная |
Спорангий со спорангиоспорами |
Зигоспора, первичный спорангий |
Ascomycota |
Endomycetes |
26 — 54 |
II |
<5 |
>5 |
Гаплоидная, диплоидная или гаплодиплоидная |
Почкующиеся клетки, конидии |
Сумка с эндогенными аскоспорами |
Ascomycetes |
46 — 60 |
Ia |
>5 |
<5 |
Гаплоидная или гаплоидно- дикариотическая |
Конидии |
||
Basidiomycota |
Ustomycetes |
48 — 70 |
Ib |
>5 |
<5 |
Гаплоидно-дикариотическая |
Почкующиеся клетки |
Промицелий с экзогенными споридиями |
Basidiomycetes |
44 — 65 |
la, Ib, Ic |
>5 |
>5 |
Конидии, почкующиеся клетки |
Базидии с экзогенными базидиоспорами |
||
Типичными представителями этого класса являются грибы рода мукор (головчатая плесень). У мукора (рис. 19) от одноклеточного мицелия отходят бесцветные спорангиеносцы, на верхушке которых развивается по одному спорангию. При наличии влаги оболочка зрелого спорангия легко растворяется, освобождая несколько тысяч спорангиоспор, которые затем прорастают. У животных и человека могут вызывать мукоромикозы.
Аскомицеты, или сумчатые грибы. Высшие грибы с разветвленным многоклеточным мицелием. Размножение вегетативное, бесполое (при помощи конидий) и половое (сумчатая стадия). В результате полового процесса возникают аски, или сумки, внутри которых после слияния ядер половых клеток (гамет) образуются аскопоры — обычно восемь в одном аске. Аскомицеты широко распространены в природе, известно около 30 тыс. видов. Обитают в почве, органических субстратах, кормах, пищевых продуктах, вызывая их порчу. Паразитируют на растениях, животных, разрушают целлюлозу. Токсические виды способны вызвать микотоксикозы. Используют как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), ферментов. К аскомицетам относятся некоторые съедобные грибы — сморчок, трюфель.
Базидиомицеты. Высшие грибы с многоклеточным мицелием. Специальным органом плодоношения служит базидия. Базидии образуют наружные споры на концах гиф в результате полового процесса. Сапрофиты и факультативные паразиты хлебных злаков (головня, ржавчина). К базидиомицетам относятся также съедобные и ядовитые шляпочные грибы.
Дейтеромицеты, или несовершенные грибы. Высшие грибы с многоклеточным, сильно разветвленным мицелием. Весь жизненный цикл их проходит в гаплоидной стадии, без смен ядерных фаз. Размножаются вегетативным и бесполым путем при помощи конидий. Конидии различаются по форме, окраске и образуются на специализированных ветвях мицелия — конидиеносцах или пикнидах (плотных органах конидиального спороношения).
Это самый многочисленный класс, включающий в себя грибы родов аспергилл, пеницилл, стахиботрис, фузариум и др. (см. рис. 19). У аспергилл, или леечной плесени, мицелий септирован, конидиеносцы одноклеточные; на их вершине формируется расширение в виде головки, от которой отходят ответвления — стеригмы — с отшнуровывающимися от них конидиями. Конидии могут быть окрашены в различные цвета, чаще черный, располагаются радиально и напоминают струйки воды, вытекающие из лейки.
У грибов рода пеницилл (кистевик) мицелий и конидиеносцы многоклеточные. В верхней части конидиеносцы разветвлены в виде кисти руки, их последние сегменты — стеригмы — заканчиваются конидиями. Образуют зеленый, белый и другие пигменты. Обитают в почве, сырых помещениях, кормах, пищевых продуктах.
К несовершенным грибам относят и дерматофиты — возбудители микоспории, трихофитии, фавуса (парши) животных, а также дрожжеподобные грибы родов Candida и Cryptococcus,вызывающие кандидоз и криптококкоз.
Дрожжи. Безмицелиальные, не образующие хлорофилла одноклеточные грибы (15). Филогенетически гетерогенная группа организмов, часть из которых типичные аскомицеты, другие — базидиомицеты, третьи — дейтеромицеты.
Это крупные сферические или палочковидные клетки размером 3.. .7 мкм; удлиненные формы могут быть более 20 мкм.
В специальной литературе часто встречается собирательный термин «плесени» («плесневые грибы»). Это нитчатые, микроскопические грибы разных классов, способные образовывать субстратный и воздушный мицелий, например, мукор, аспергиллы, пенициллы и др.
Клетки микроскопических грибов разнообразны по форме, размерам, но имеют общие структурные элементы. Клетка всех грибов состоит из клеточной стенки, цитоплазмы с цитоплазматической мембраной и эндоплазматической сетью, митохондриями, рибосомами, включениями, вакуолями, ядром или несколькими ядрами (рис. 2).
2.2 Морфология и ультраструктура грибов
В ходе жизненного цикла грибов происходит образование вегетативных и репродуктивных структур. Вегетативная структура — это мицелий и его видоизменения. Любая часть мицелия в благоприятных условиях может дать начало новой особи.
Репродуктивными называют структуры, специально предназначенные для размножения.
У грибов встречаются три типа размножения: вегетативное, бесполое и половое. У многих видов они последовательно сменяют друг друга в цикле развития.
Вегетативное размножение обычно осуществляется неспециализированными частями мицелия, которые дают начало новому мицелию. Мицелий большинства грибов обладает высокой способностью к регенерации, что и лежит в основе этого способа размножения и широко используется, например, при приготовлении грибницы для искусственного выращивания съедобных грибов, таких как шампиньон двуспоровый, вешенка обыкновенная и др., а также при получении биомассы грибов в пищевых и кормовых целях.
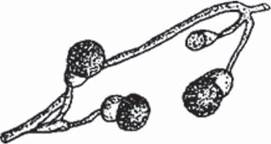
К специализированным структурам вегетативного размножения относятся оидии, тонкостенные клетки, и хламидоспоры, толстостенные клетки, на которые распадается мицелий и которые дают начало новому мицелию. Хламидоспоры выполняют и функцию перенесения неблагоприятных условий (рис. 19, 20). У большинства дрожжей вегетативное размножение происходит путем почкования клеток.
Рис. 20. Хламидоспоры грибов в чистой культуре: хламидоспоры Mycogone perniciosa — возбудителя мягкой гнили шампиньона [9].
Бесполое размножение осуществляется при помощи разнообразных специализированных клеток или многоклеточных структур, спор. У грибов известны
эндогенные и экзогенные споры бесполого размножения. Эндогенные подвижные споры, зооспоры, развиваются в зооспорангиях разной формы. Это голые клетки, снабженные жгутиками, число, расположение и строение которых различно в разных систематических группах грибов. Зооспоры разного строения характерны для представителей отделов оомикота, гифохитридиомикота и хитридиомикота, т. е. в основном для водных и реже наземных грибов. Для осуществления размножения с помощью зооспор нужна вода, хотя бы в виде отдельных капель на поверхности почвы или растений, в которой зооспоры могут передвигаться с помощью жгутиков. Эндогенные неподвижные споры, спорангиоспоры, одеты оболочкой и образуются внутри спорангиев, развивающихся на специализированных гифах, спорангиеносцах, обычно поднимающихся над субстратом. Спорангиоспорами осуществляется бесполое размножение у зигомикот.
Экзогенные споры бесполого размножения грибов, конидии, неподвижны, образуются на специализированных, обычно морфологически отличных от вегетативного мицелия, дифференцированных спороносцах, конидиеносцах. Типичные конидии характерны для сумчатых, базидиальных и анаморфных грибов. Как и у зигомикоты, это в основном наземные грибы, и распространение неподвижных спор бесполого размножения, спорангиоспор и конидий у таких грибов осуществляется, в основном пассивно токами воздуха или воды. Иногда распространение спор может осуществляться с помощью животных, например, при поедании плодовых тел шляпочных грибов. Специализированные структуры, связанные с вегетативным и бесполым размножением у грибов, называются анаморфами.
2.3 Организация клеточного цикла дрожжей
Наиболее исследован клеточный цикл у дрожжей Saccharomyces cerevisiae., хотя многие положения являются общими для всех эукариотических клеток (рис. 21).
Клеточный цикл эукариотов разделен на четыре фазы: G1, S, G2, М. Синтез ядерной ДНК занимает лишь некоторую часть интерфазы — период, называемый S-фазой клеточного цикла. Между концом М-фазы и началом синтеза ДНК имеется интервал, известный как фаза G1(от англ. gap — промежуток). Другой интервал, называемый фазой G2, отделяет конец синтеза ДНК от начала М-фазы. Таким образом, интерфаза состоит из последовательности фаз: G1, S, G2 и обычно занимает не меньше 90% всего времени клеточного цикла. В клеточном цикле выделяют три особых критических точки: в фазе G1, в фазе S и в фазах G2+M (G2/M -точка), которые контролируются внутриклеточными процессами. Продолжительность фаз клеточного цикла неодинакова; в среднем фаза G1 составляет 30-40% от всей продолжительности клеточного цикла, фаза S — 30-50%, фаза G2 — 10-20% и фаза М 5-10%. Эти параметры могут достаточно сильно варьироваться в зависимости от биологического вида, но достаточно неизменны для конкретного типа клеток. Существует особая критическая точка в фазе G1 названная точкой старта (Start). При прохождении этой точки (для высших эукариотов — точка рестрикции) в клетке происходят изменения, после которых она должна уже пройти все последующие этапы клеточного цикла. Такое детерминирование клеточного цикла предполагает внутриклеточный контроль завершения каждой из фаз клеточного цикла (Hartwell L.H., 1974). Иными словами, после точки старта клетка детерминирована в завершении цикла и выходы из него невозможны (но не остановки) в любой другой фазе (S, G2, М). Такое детерминирование клеточного цикла предполагает внутриклеточный контроль завершения каждой из фаз клеточного цикла. События, происходящие, например, в фазе G2, не могут разворачиваться при прохождении фазы S, и наоборот.
Если после окончания клеточного цикла не имеется условий для вхождения в новый цикл, клетки переходят в состояние GO. По своему определению это состояние просто характеризует клетки, не включенные в клеточный цикл. В этом состоянии могут находиться клетки, которые подготавливаются к входу в клеточный цикл (например, увеличивают свой размер до критического, необходимого для прохождения точки Старта), в этом случае тестируется (как бы) временное увеличение фазы G1. Иногда, такими добавочными условиями для вхождения в клеточный цикл, является синтез не конституционных ферментных систем, необходимых для специфического субстрата (например, для роста дрожжей на н-алканах необходима индукция системы Р-450 (Галынкин В. А. и др., 1990)). С другой стороны, в состоянии GO находятся и клетки, осуществляющие синтез целевого продукта (например, биосинтез антибиотиков), в этом случае мы имеем дело с физиологической дифференцировкой. В состоянии GO находятся клетки, осуществляющие морфологическую дифференцировку (например, образование спор). В некоторых случаях морфологическая и физиологическая дифференцировка совпадают (синтез некоторых антибиотиков и переход к споруляции совпадает по времени). Наконец, морфологически дифференцированые клетки могут войти в клеточный цикл в этом статусе. В этом случае наблюдается рост не отдельных клеток, а в виде мицелия, что также приводит к среднему увеличению времени деления клеток. По существу, из GOсостояния клетки либо возвращаются в клеточный цикл (иногда в новом морфологическом статусе), либо переходят к программированной клеточной гибели (автолизная программа).
Для дрожжей Saccharomyces cerevisiae (рис. 21) один из генов, контролирующих прохождение точки старта, был выявлен и назван cdc28 (аналог cdc2). Белок CDC28, кодируемый этим геном, представляет собой протеинкиназу, но для функционирования этого белка требуется наличие другого белка, названного циклином. Именно циклин и является тем белком, который периодически синтезируется в ходе клеточного цикла и исчезает к его окончанию (Э. У. Мюрей, 1991).
Дальнейшие исследования показали, что уместно говорить не об одном циклине, а о целом семействе циклинов. Были найдены три группы циклинов: циклины фазы Gi (GI cyclins), контролирующие вхождение в цикл (точка старта); циклины фазы S (S-phase cyclins), контролирующие прохождение фазы S; циклины фазы М (M-phase cyclins), контролирующие завершение цикла. Причем циклины, ответственые за каждую из фаз цикла, синтезируются в своей фазе, а затем ферментативно деградируют (распадаются) (Dirick L. 1995, Nasmyth К., 1996). Было показано, что фазы цикла инициируются растворимыми цитоплазматическими факторами. Активатор М-фазы был назван М-стимулирующим фактором (М-phasepromoting factor, MPF), а активатор S-фазы назван S-стимулирующим фактором (S-phase-promoting factor). MPF имеет универсальное значение для эукариотических клеток и в эволюции высококонсервативен.
Рис. 21. Показаны четыре фазы: G1, S, G2, M и выход из клеточного цикла состояние Go Показаны соответствующие фазо-стимулирующие факторы (phase-promoting factor),представляющие комлекс циклина (cyclin) — кружок и циклин зависимой протеинкиназы (cydindependent kinases — CDK) — квадрат. После прохождения фазы цикла показана деградация циклинов. На рисунке также представлен стимулирующий комплекс анафазы (anaphase-promoting complex — АРС), необходимый для прохождения одной из стадий митоза — анафазы (расхождение хромосом).
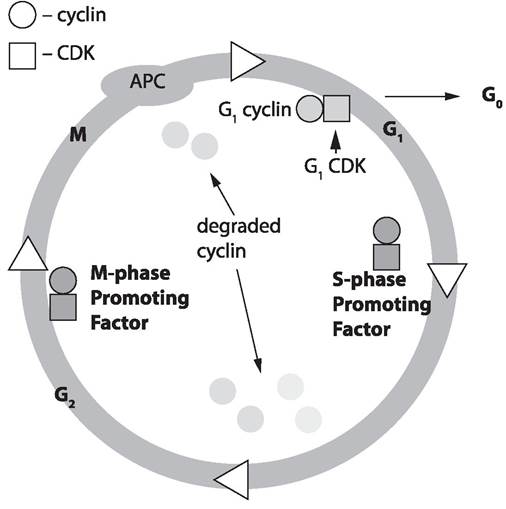
Механизм регуляции цикла построенный на активации циклинзависимых протеинкиназ (cyclindependent kinases — CDK), выполняется для всех эукариотов, но его детали сильно различаются для конкретного типа эукариотической клетки.
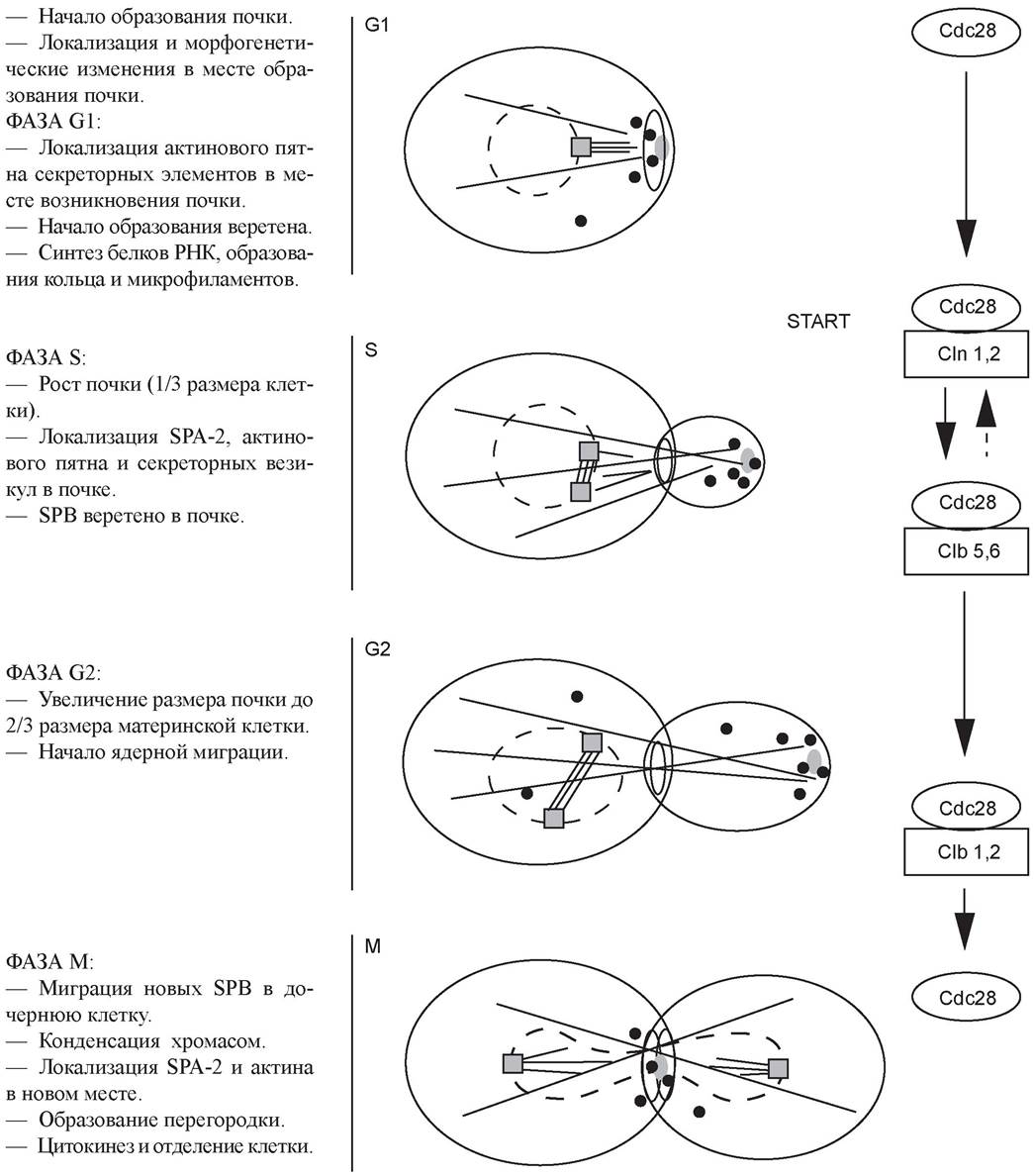
На рисунке 22 показано бесполое размножение дрожжей Saccharomyces cerevisiae с одновременным прохождением биохимических процессов и изменением генетического статуса клетки.
Рис. 22. Бесполое размножение дрожжей
Половое размножение у грибов, связанный с ним процесс смены ядерных фаз, строение половых органов — всё это отличается у разных групп грибов. Существеннейшие моменты полового процесса у грибов: плазмогамия, кариогамия и мейоз. Соответственно, гриб может находиться в гаплоидной или диплоидной стадиях. Плазмогамия и кариогамия у сумчатых и базидиальных грибов не совпадают во времени, в результате чего после плазмогамии у них возникает особая стадия дикариотичного мицелия, когда гаплоидные ядра попарно ассоциированы, сближены, но не слились и образуют дикарион. Ядра дикариона обычно синхронно делятся с параллельным расположением осей веретен деления. В определенный момент цикла развития они сливаются, образуя диплоидное ядро, которое затем делится редукционно. Смена ядерных фаз будет подробно рассмотрена при описании отделов и классов грибов.
В результате полового процесса образуются гаплоидные, неоднородные в генетическом отношении споры, что принципиально отличает их от спор бесполого размножения грибов. Эти гаплоидные споры располагаются или на мицелии, или чаще, на поверхности или внутри плодовых тел различного строения, которые называются телеоморфами. Таким образом, размножение с помощью спор, возникших половым путем, даёт начало формам с новой комбинацией генетического материала, что является основой дальнейшей эволюции форм, а размножение спорами, возникшими бесполым путем, способствует распространению и сохранению данной формы.
Типы полового процесса у грибов. Половое размножение у грибов известно у всех групп, за исключением анаморфных, или несовершенных, грибов. Половой процесс у грибов разнообразен и его особенности лежат в основе выделения классов. У грибов известно три основных типа полового процесса: гаметогамия, гаметангиогамия и соматогамия (рис. 23).
Рис. 23. Типы полового процесса у грибов [9].
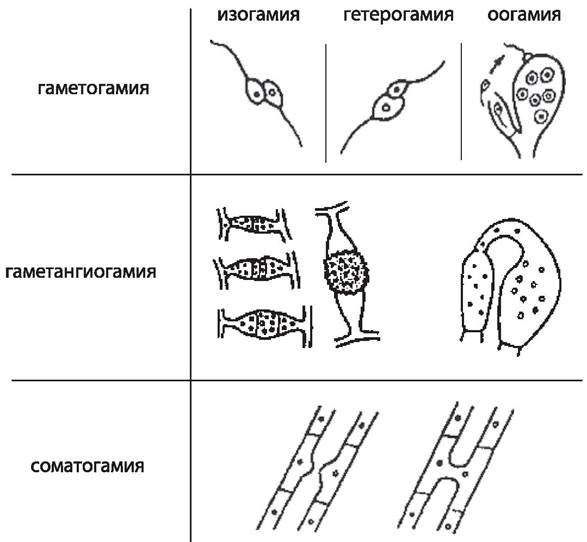
Гаметогамия — слияние гамет, образующихся в гаметангиях. Различают изогамию — слияние подвижных морфологически сходных гамет и гетерогамию — слияние подвижных, отличающихся по размерам и часто по степени подвижности гамет. Эти два типа гаметогамии характерны для хитридиевых и гифохитридиевых грибов. При оогамии крупные, неподвижные яйцеклетки, формирующиеся в специальных оогониях, оплодотворяются мелкими, подвижными сперматозоидами, развивающимися в специализированных антеридиях. У многих грибов с этим типом полового процесса сперматозоиды не образуются, а яйцеклетка оплодотворяется недифференцированным на сперматозоиды содержимым выростов многоядерного антеридия. Такой тип оогамии характерен для всех представителей отдела оомикота.
Второй тип полового процесса, гаметангиогамия, состоит в слиянии двух обычно многоядерных специализированных структур, содержимое которых не дифференцировано на гаметы. Гаметангиогамия характерна для зиго- и аскомикот. Гаметангиогамия у зигомикот носит название зигогамии. Она заключается в слиянии в основном многоядерных клеток, гаметангиев, хорошо отличимых от вегетативного мицелия, на котором они формируются, но недифференцированных морфологически по половому знаку на мужской и женский. Из образовавшейся в результате их слияния зиготы формируется одетая толстостенной окрашенной оболочкой зигоспора, которая прорастает после периода покоя в особый зародышевый спорангий (рис. 23, 24).
Рис. 24 (1). Жизненный цикл и зигота Rhizopus nigricans: а — спорангий, б — спорангиеносец, в — ризоиды, г — зигоспора, д — подвески суспензоры, е — столон.
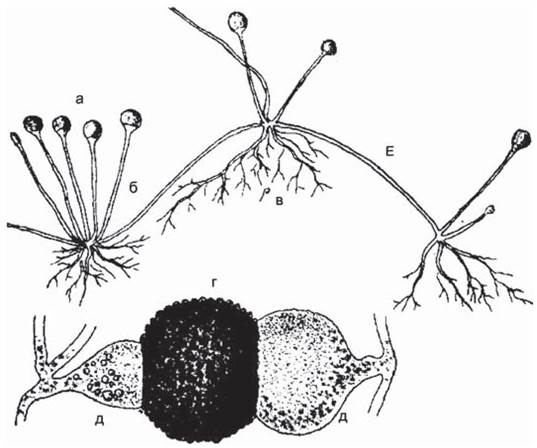
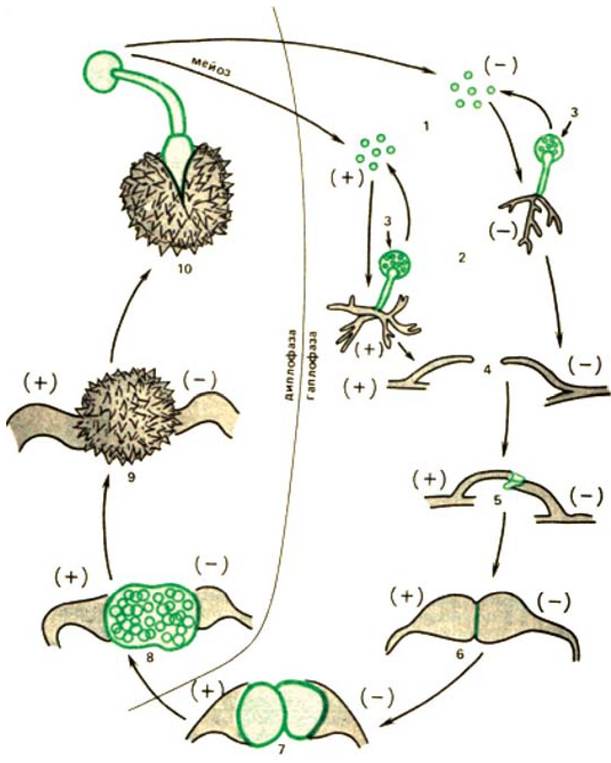
Рис. 24 (2). Жизненный цикл Мucor mucedo. 1 — гаплоидные спорангиоспоры; 2 — вегетативный мицелий; 3 — спорангии, образующиеся на вегетативном однодомным мицелии; 4 — образование зигофор в результате взаимодействия (+) и (-) нитей мицелия; 5 — слияние зигофор; 6 — формирование парагаметангия; 7 — образование ограниченного гаметангия с суспензорами; 8 — молодая зигоспора; 9 — зрелая зигоспора; 10 — проростание зигоспоры [11].
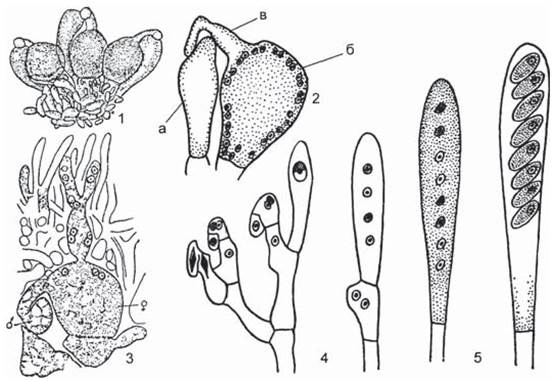
У аскомикот при гаметангиогамии также сливаются два многоядерных гаметангия, но у них, в отличие от зигомикот, половые органы дифференцированы на женский — аскогон и мужской — антеридий. Аскогон состоит из двух клеток: крупной многоядерной, или собственно аскогона, и тонкой нитевидной — трихогины, помещающейся на его вершине, через которую в аскогон переливается содержимое многоядерного антеридия. При этом происходит только плазмогамия, а ядра ассоциируются в пары, образуя дикарион. Из оплодотворенного аскогона без периода покоя вырастают аскогенные дикариотичные гифы. В их клетках происходит слияние ядер дикариона и образование диплоидного ядра, которое в дальнейшем делится мейотически. В результате этого процесса на аскогенных гифах достаточно сложным путем формируются особые образования — сумки, или аски, внутри которых после митотического деления пост- мейотических ядер формируются восемь эндогенных гаплоидных аскоспор (рис. 25).
Рис. 25. Половой процесс и развитие сумок у Pyronema omphaloides: 1 — группа половых органов, 2 а — антеридий, б, в — аскогон с трихогиной, 3 — развитие двуядерных (дикариотичных) аскогенных гиф из оплодотворенного акскогона, 4 — схема образования сумок из аскогеных гиф с помощью крючка, 5 — молодая и зрелая сумка с аскоспорами [9].
Третий тип, соматогамия, — половой процесс, при котором сливаются обычные соматические, или вегетативные клетки мицелия. Половые органы и гаметы отсутствуют. Соматогамия характерна для некоторых представителей отделов хитридиомикота и гифохитридиомикота, имеющих одноклеточный таллом. В этом случае целиком сливаются две одноклеточные особи. Такой тип соматогамии называется хологамией. Соматогамия у базидиомикот заключается в слиянии двух вегетативных клеток гаплоидного мицелия. При этом, как и у аскомикот, сначала имеет место только плазмогамия, в результате чего формируются дикарионы и образуется дикариотический (состоящий из двуядерных клеток) мицелий. Это наиболее длительная стадия в цикле развития базидиальных грибов. Затем на этом дикариотическом мицелии формируются особые клетки, базидии, в которых происходит слияние ядер дикариона и мейотическое деление диплоидного ядра, после чего на базидии формируются экзогенные гаплоидные базидиоспоры (рис. 26).
Рис. 26. Типы базидий. 1 — холобазидия; 2-4 — гетеробазидии; 5 — телио- (фрагмо-) базидия [9].
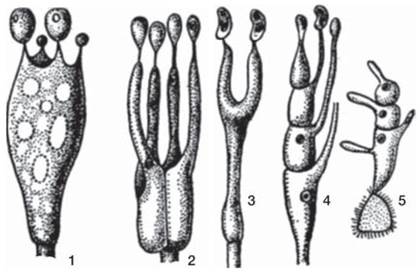
Эндогенные аскоспоры сумчатых грибов и экзогенные базидиоспоры базидиальных грибов образуются в результате полового процесса, т. е. их появление связано с половым размножением этих групп. Половой процесс сумчатых и базидиальных грибов имеет две характерные общие особенности: во-первых, разрыв между плазмогамией и кариогамией и появление дикариотической фазы и, во-вторых, отсутствие у зиготы состояния покоя: мейотическое деление диплоидного ядра происходит сразу же после слияния гаплоидных ядер дикариона.
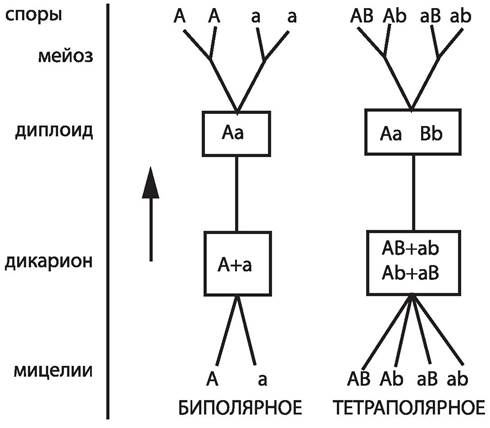
По характеру половой дифференцировки у грибов различают гомоталличные (обоеполые) и гетероталличные (раздельнополые) формы. У гомоталличных грибов к слиянию способны клетки одного и того же мицелия. На одном и том же мицелии формируются и мужской и женский половые органы (например, оогонии и антеридии у оомикот). У гетероталличных грибов на мицелии, выросшем из одной споры, половые органы не закладываются и, соответственно, зиготы не образуются. Они развиваются лишь при встрече двух мицелиев, отличающихся друг от друга по половому знаку (+ и -, или мужской и женский). Понятие гетероталлизма относится к гаплоидной стадии, так как определение пола у грибов происходит в основном генотипически. Гетероталлизм, или раздельнополость, у грибов может быть двух типов: биполярный, когда пол определяется одной парой аллелей, и тетраполярный, когда пол определяется двумя парами аллелей, локализованных в разных хромосомах и независимо комбинирующихся. В случае биполярного определения пола все гифы, выросшие из спор одного плодового тела, распадаются на две группы, и при соединении мицелиев из этих двух разных групп происходит половой процесс. В случае тетраполярного определения пола гифы, выросшие из спор одного плодового тела, распадаются на четыре половые группы. При этом группа I сливается только с группой II, и группа III только с группой IV. Численные соотношения этих групп для шляпочных грибов (базидиомицетов), у которых тетраполярный гетероталлизм распространён очень широко, соответствует отношению 1:1:1:1 (рис. 27).
Рис. 27. Схема распределения пола у грибов.
У несовершенных (анаморфных) грибов половой процесс отсутствует, и жизненный цикл эти грибы проводят в гаплоидном состоянии. В определённой степени отсутствие полового процесса в этой группе компенсируется гетерокариозом и происходящим на его основе парасексуальным процессом. Гетерокариоз, или разноядерность, — это наличие в клетках мицелия генетически разных ядер. Гетерокариоз характерен для многих групп грибов и обеспечивает адаптацию грибов к изменяющимся условиям среды. В таком мицелии ядра иногда могут сливаться, образуя диплоидное гетерозиготное ядро. Такое ядро делится митотически, при этом происходит митотическая рекомбинация и затем вегетативная гаплоидизация этих диплоидных ядер путем потери ими части хромосом. Этот сложный процесс, включающий в качестве существеннейшего момента митотическую рекомбинацию, получил название парасексуального процесса. Он известен у разных групп грибов, и имеет особое значение для несовершенных грибов, лишённых настоящего полового процесса.
2.4 Особенности деления ядра
Митоз и мейоз у грибов отличаются рядом специфических особенностей. У большинства видов грибов деление ядра происходит по закрытому типу, то есть с сохранением ядерной оболочки. Центриоли имеются лишь у псевдогрибов и некоторых грибов, имеющих жгутиковые стадии, у остальных видов веретено деления формируется более просто устроенными белковыми структурами — полярными тельцами веретена (ПТВ). Фазы митоза чередуются быстро, а хромосомы имеют небольшие размеры; в сочетании эти факторы затрудняют микроскопическое исследование, поэтому ранее считалось, что деление ядер у грибов происходит амитотически. Телофаза митоза происходит несинхронно, в результате чего могут образовываться гетероплоидные дочерние ядра, то есть содержащие неравное число хромосом. Чаще всего при гетероплоидии наблюдается различное число В-хромосом. Митоз и образование новых клеток (цитокинез) у мицелиальных (не дрожжевых) грибов происходят независимо друг от друга — ядра перемещаются в дочернюю клетку уже после отделения её перегородкой (септой) от материнской (у грибов с неклеточным мицелием цитокинез вообще наблюдается редко, при регенерации повреждённых участков и при образовании репродуктивных органов).
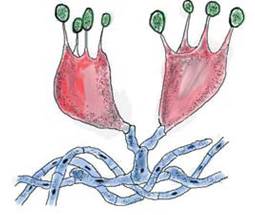
Репродукционное бесполое размножение происходит с помощью особых клеток — спор (рис. 22, 28), образующихся без участия полового акта. Репродукционное половое размножение включает обмен генетическим материалом при слиянии ядер (кариогамии) и редукционное деление (мейоз), связанные с образованием определенных морфологических структур.
Рис. 28. Рисунок двух базидий (показаны красным цветом) с базидиоспорами (показаны зелёным)
Бесполые репродуктивные структуры называют анаморфами, а половые — телеоморфами. Грибы, имеющие половую стадию развития, называют совершенными. Грибы, в жизненном цикле которых отсутствует половое размножение, называют несовершенными, дейтеромицетами или митоспоровыми грибами.
Анаморфы и телеоморфы строго приурочены к определенной фазе жизненного цикла гриба, образуются при наличии особых условий внешней среды и выполняют дифференцированные функции (табл. 5).
Таблица 5. Функциональная и морфологическая дифференцировка таллома
Фаза цикла развития |
Функция |
Морфологическая структура |
Вегетативная |
Прорастание |
Проростковые гифы, почкующиеся клетки |
Разрастание по субстрату |
Столоны, мицелий, ризоморфы, тяжи |
|
Поселение на хозяине |
Апрессории, гифоподии, инфекционные и перфорационные гифы, гаустории, ловчие гифы |
|
Перенесение неблагоприятных условий |
Хламидоспоры, склероции |
|
Репродукционная бесполая |
Размножение и распространение |
Спорангиеносцы (спорангиофоры), спорангиеспоры, конидиеносцы (конидиофоры), конидии, зооспоры Гаметы, гаметангии, оогонии, аскогонии, антеридии, трихогины, ооспоры, зигоспоры, аски (сумки), аскоспоры, базидии, базидиоспоры, плодовые тела |
2.5 Ультраструктура грибной клетки
Клеточная стенка
Клеточная стенка (рис. 2, 29) обеспечивает механическую прочность клетки, постоянство ее формы, служит барьером проницаемости и защищает клетку от внешних воздействий. Характерная черта клеточной стенки — способность к росту и интенсивной перестройке в течение развития грибов (в их онтогенезе).
Рис. 29. Модель цитоскелета дрожжевой клетки [12].

Компоненты клеточной стенки можно подразделить на две группы: структурные компоненты (сеть микрофибрилл) и соединения, заполняющие пространство между ними (матрикс). Первые представлены полисахаридами, включающими полиаминосахара (хитин и хитозан) и глюканами, имеющими β- (1, 3), β- (1,4), β- (1, 6) связи. Вторые являются маннопротеинами, галактоманнопротеинами, глюкуронман- нопротеинами, ксиломаннопротеинами и α- (1, 3) глюканами. У мицелиальных и дрожжевых грибов имеются существенные различия в химическом составе и структуре клеточной стенки. Так содержание хитина составляет у мицелиальных грибов 0,2-26,2% от сухой массы клеточных стенок, а у дрожжевых 1-4%. Помимо полисахаридов в клеточной стенке присутствуют белки и липиды. Некоторые белки являются ферментами. Липиды клеточной стенки определяют ее гидрофобность и принимают участие в синтезе компонентов клеточной стенки, активируя хитинсинтетазу.
У многих грибов, особенно дрожжевых, внешняя часть клеточной стенки образует капсулу — сильно оводненный слизистый слой полисахаридной природы. В капсуле локализованы многие ферменты, она принимает участие в поглощении питательных веществ из субстрата, участвует в адгезии клеток между собой и субстратом, защищает клетку от внешних воздействий (высушивания, радиации и т. п.).
Различия в размере капсулы в одной и той же культуре связаны с возрастным и физиологическим состоянием клеток.
Органеллы грибной клетки
Органеллы грибной клетки характерны для большинства эукариот. В ней имеется ядро, содержащее наследственную информацию в виде ДНК, оформленной в хромосомы. Ядро заполнено нуклеоплазмой, имеет ядрышко — место синтеза прорибосом, окружено двойной оболочкой (ядерной мембраной). Ядерная мембрана имеет поры, которые связывают нуклеоплазму с цитоплазмой.
Митохондрии содержат ферменты дыхательной цепи и окислительного фосфорилирования, а также цикла трикарбоновых кислот, обеспечивая энергетические потребности клетки.
Рибосомы 80 S — это органеллы синтеза белка. Часть рибосом располагается в цитоплазме беспорядочно, однако большинство их прикреплено к мембранам эндоплазматического ретикулума, митохондриям и другим органеллам.
Одним из основных компонентов клеточных органоидов являются мембраны. С ними связано большинство метаболических процессов. Они составляют от 40 до 90% общей массы клеток.
Цитоплазматическая мембрана служит осмотическим барьером, в ней локализуется система активного транспорта, она способна к пиноцитозу и фагоцитозу.
Эндоплазматический ретикулум (ЭР) расположен в цитоплазме в виде сети канальцев, цистерн и трубочек, не имеющих фиксированной ориентации, выполняет транспортную функцию, связывает цитоплазматическую мембрану с ядерной, образует поверхности раздела в цитоплазме, на его мембранах могут располагаться рибосомы.
Аппарат Гольджи представляет собой систему вакуолек, он обеспечивает экскрецию (выведение) по типу обратного пиноцитоза с помощью пузырьков, аккумулирующих выводимые продукты, и транспорт веществ, синтезированных в ЭР, к другим органоидам. Кроме того, аппарат Гольджи — место синтеза новых мембран и образования лизосом.
Лизосомы содержат около сотни ферментов, преимущественно гидролаз, осуществляющих функцию пищеварения.
Вакуоли образуются из ЭР и выполняют многообразные функции. В них могут накапливаться вредные продукты метаболизма, они участвуют в компартментализации веществ в клетке (их разделении и концентрации), в них накапливаются необходимые клетке метаболиты (полифосфаты, аминокислоты, пуриновые и пиримидиновые основания). Вакуоли обеспечивают тургор клеток.
Новообразование мембран ЭР и Гольджи происходит на матрице ядерной мембраны. Цитоплазма содержит включения, которые служат запасными питательными веществами: гликоген, волютин (полифосфат), липиды.
Цитоскелет наряду с мембранными структурами обеспечивает внутриклеточную организацию и высокую биологическую упорядоченность всех метаболических процессов, протекающих в клетке. Цитоскелет представляет собой развитую сеть белковых нитей (филаментов), из которых наиболее важная роль принадлежит микрофиламентам и микротрубочкам. Те и другие состоят из глобулярных белковых субъединиц, которые в клетке могут легко соединяться между собой и разъединяться. Помимо этого, существуют вспомогательные белки, которые либо связывают филаменты друг с другом или с другими клеточными структурами, либо влияют на скорость и степень полимеризации филаментов. Цитоскелет связан с движением органоидов клетки и амебоидным движением, присущим некоторым грибам.
Другие подвижные клетки грибов (зооспоры, планогаметы) передвигаются с помощью жгутиков. Жгутики грибов по строению отличаются от жгутиков бактерий, но сходны с аналогичными органеллами простейших и многих подвижных гамет растений и животных.
2.6 Экологические группы грибов
Грибы широко распространены в природе на самых различных субстратах. Они предпочитают водные или влажные местообитания, но встречаются и в относительно сухих средах. Грибы являются гетеротрофами, потребляют органические соединения углерода. Азот, фосфор, сера, ионы металлов могут поглощаться в неорганической форме. Питательные вещества грибы получают, принимая участие в разложении органического вещества или паразитируя на животных и растениях.
Многие грибы переносят значительные изменения температуры, мезофильные грибы предпочтительно развиваются в пределах 24-3 0°С. Термофильные — в диапазоне 33-55°С, психрофильные — от -2°С до +20°С. Свет, особенно в коротковолновой области, может влиять на спороношение грибов.
Экологические группы грибов сформировались в процессе эволюции. Сапротрофные грибы принимают участие в минерализации органических веществ, образовании гумуса, ксилотрофные грибы разлагают древесину, кератинофилы способны жить на волосах, перьях, рогах павших животных. Симбиотрофные грибы образуют микоризу, имеющую огромное значение для жизни многих растений. Лишайники — это стабильные симбиотические ассоциации гриба с водорослями или цианобактериями. Грибы-паразиты растений и животных описаны ниже.
Грибы, как и другие организмы, способны существовать в относительно узком диапазоне температур, влажности, почвенных условий и других факторов среды. Эти условия определяют их географическое распространение. Распространению популяций препятствуют географические преграды (океаны, пустыни, горные цепи) и способствуют агенты, действующие как средства переноса: воздух, вода, животные и человек.
Для некоторых грибов характерна способность находиться длительное время в неизменных и ограниченных зонах — эндемических областях (очагах). К таким грибам относятся возбудители глубоких микозов Coccidioides immitis и Paracoccidioides brasiliensis, которые встречаются в зонах с определенными климатическими условиями (Центральная Америка для первого гриба и Южная Америка для второго). Дерматофиты Trichophyton soudanense обнаруживается только в Африке, а T. concentricum — в Южной Океании. Многие грибы, в том числе возбудители многих микозов человека и фитопатогенные грибы, являются космополитами, их можно обнаружить в любой местности, где условия для них окажутся благоприятными.
2.7 Использование грибов в промышленности
С древних времен человек использует дрожжи Saccharomyces cerevisiae для изготовления хлеба, пива, вина. Современная промышленная микробиология использует грибы для получения разнообразных продуктов: антибиотиков, алкалоидов, белков, витаминов, гербицидов, ферментов, коферментов, ингибиторов ферментов, полисахаридов, липидов, органических кислот и др. В табл. 6 приведены антибиотики и другие БАВ, выпускаемые промышленностью для медицины и сельского хозяйства; в табл. 7 — сведения о практическом применении ферментов грибов; в табл. 8 приведены органические кислоты, которые получаются с помощью грибов. Некоторые дрожжевые грибы используют для получения кормового белка на непищевом сырье. Многие грибы из класса базидиомицетов (макромицеты, образующие крупные плодовые тела) обладают целебными свойствами, их культивируют поверхностным и глубинным (в ферментерах) способами и используют как лекарственные препараты и пищевые добавки (табл. 9).
Таблица 6. Биологически активные вещества грибов, выпускаемые промышленностью
Применение |
Вторичный метаболит |
Продуцент |
Действие |
Медицина |
Пенициллины |
Penicillium chrysogenum |
Антибактериальное |
Цефалоспорины |
Cephalosporium acremonium |
Антибактериальное |
|
Гризеофульвин |
P. griseofulvum |
Антигрибное |
|
Фузидин |
Fusidium coccineum |
Антибактериальное |
|
Циклоспорин |
Trichoderma polysporum |
Иммунодепрессивное |
|
Эргоалкалоиды |
Claviceps purpurea |
Нейротропное |
|
Сельское хозяйство |
Зеараленон |
Gibberella zeae |
Стимулятор роста крупного рогатого скота |
Гибберелины |
G. fujikuroi |
Регуляторы роста растений |
Таблица 7. Ферменты грибов, выпускаемые промышленностью
Продуцент |
Фермент |
Применение фермента |
Aspergillus niger, A. oryzae |
α-амилаза |
Расщепление крахмала |
A. niger |
Амилоглюкозидаза |
Получение крахмальных сиропов |
Aureobasidium pullulans |
Глюкоамилаза |
Расщепление крахмала |
A. niger |
Глюкозооксидаза |
Получение глюконовой кислоты |
Aspergillus species |
Протеиназы (кислые, нейтральные, щелочные) |
Расщепление белков (в хлебопечении, виноделии) |
Saccharomyces cerevisiae |
Инвертаза |
Расщепление сахарозы |
Aspergillus species, Rhisopus species |
Пектиназы |
Осветление фруктовых соков |
Mucor species |
Реннин |
Створаживание молока |
Mucor species, Aspergillus species |
Глюкозоизомераза |
Получение сиропов с высоким содержанием фруктозы |
Mucor species, Aspergillus species, Penicillium species |
Липазы |
Получение детергентов, в молочной промышленности |
A. niger, Trichoderma roseum, Penicillium notatum |
Целлюлазы |
В целлюлозно-бумажной промышленности |
Fusarium species, Penicillum species. Trichoderma species |
Ксиланазы |
В целлюлозно-бумажной промышленности, в сельском хозяйстве |
Таблица 8. Органические кислоты, получаемые с помощью грибов
Кислота |
Продуцент |
Глюконовая |
AspergiNum niger |
Койевая |
A. oryzae |
Лимонная |
A. niger , Candida lipolytica |
Итаконовая |
A. terreus |
Трео-изолимонная |
C. brumptii C. lipolytica |
Аллоизолимонная |
PeniciMium purpurogenum |
а-Кетоглутаровая |
C. lipolytica |
Фумаровая |
Rhizopus delemar |
Ябочная |
C. hydrocarbofumarica SchizophyNum commune, C. hydrocarbofumarica + Pichia membranaefaciens |
Тетрадекандикарбоновая |
C. doacea |
Таблица 9. Лечебные свойства базидиомицетов [13]
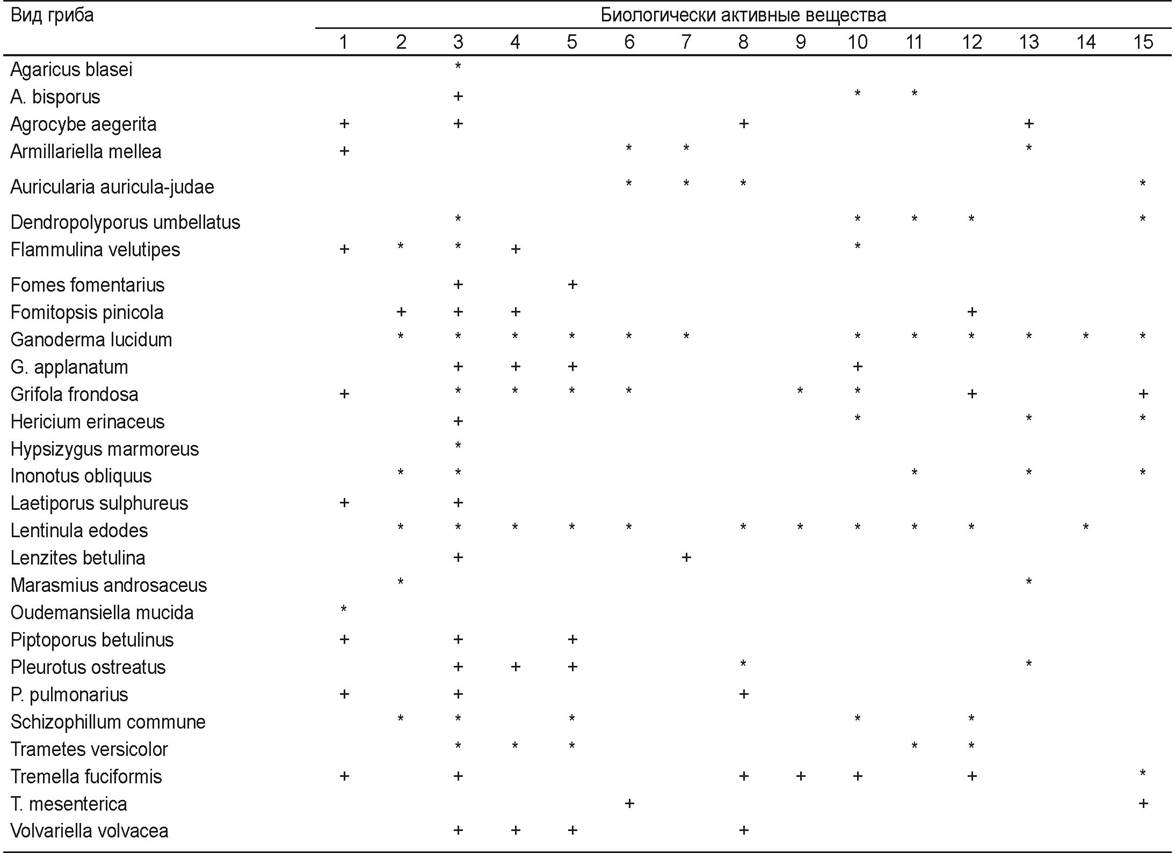
Примечание: * - коммерческий препарат (лекарственный или пищевая добавка), + - некоммерческий продукт
Биологически активные вещества:
1 - антифунгальные
2 - противовоспалительные
3 - противоопухолевые
4 - антивирусные
5 - антибактериальные и антипротозойные
6 - регулирующие давление крови
7 - потенцирующие сердечно-сосудистую систему
8 - антихолестеринемические, антилипидемические
9 - антидиабетические
10 - иммуномодулирующие
11 - тонизирующие почки
12 - гепатопротективные
13 - тонизирующие нервную систему
14 - потенцирующие половую функцию
15 - применяющиеся при хроническом бронхите
2.8 Грибы — возбудители болезней человека и животных
Грибы могут наносить вред человеку и животным путем отравления их метаболитами, вызывая состояние повышенной чувствительности к различным веществам, входящим в состав их клеток или продуцируемыми ими (микогенная аллергия), и вызывая инфекционные заболевания (микозы).
Отравление может происходить при поедании ядовитых или испорченных грибов, при неправильном приготовлении и хранении шляпочных грибов. Их токсины действуют на пищеварительную, нервную системы и на другие ткани тела.
Другая группа заболеваний, вызванных отравлением метаболитами грибов (микотоксикозы), связана с тем, что токсинообразующие грибы заселяют растения, продуцируя при этом или во время хранения урожая ядовитые вещества, действующие и после переработки растительных продуктов в корма или продовольствие.
Микогенные аллергии возникают у чувствительных людей в ответ на антигенные вещества грибов. Они проявляются в виде кожной сыпи, насморка, конъюктивита, диарреи, астматических явлений и т. п. Аллергенами могут быть клетки мицелия, споры гриба и продукты их обмена или распада. Эти заболевания могут возникать и развиваться у персонала биотехнологических предприятий, использующих грибы как продуценты БАВ, и у населения близлежащих районов за счет загрязнения воздуха при плохой очистке заводских выбросов. Меры предупреждения состоят в строгом соблюдении величины санитарной зоны у предприятия, техники безопасности (защитная одежда, маски, респираторы, герметизация оборудования), тщательной очистки воздуха, контактировавшего с микроорганизмами.
Опасна также работа в складских помещениях, где плесневые грибы могут развиваться при неправильном хранении сырья в условиях высокой влажности, образовании конденсата на поверхностях и т. д. Такие помещения должны быть снабжены приточно-вытяжной вентиляцией, а температуру необходимо поддерживать постоянной, чтобы предотвратить появление конденсата.
Микозы человека. В последние десятилетия наиболее часто встречаются оппортунистические микозы, возникающие при ослабленном иммунитете и вызываемые условно-патогенными грибами, которые встречаются среди представителей родов Penicillium, Aspergillus, Mucor, Alernaria, Cladosporium, Fusarium, Candida, Geotrichum, Saccharomyces, Rhodotorula, Sporobomyces, Trichosporon (табл. 10). Они могут входить в состав нормальной микробиоты человека и животных и активируют свои паразитарные свойства под влиянием нерациональной антибиотической и кортикостероидной терапии, при использовании иммунодепрессантов и при ослаблении реактивности организма в связи с предшествующим заболеванием.
Таблица 10. Основные возбудители микозов человека
Возбудитель |
Резервуар |
Способ заражения |
Болезнь |
||
Класс |
Телеоморфа |
Анаморфа |
|||
1 |
2 |
3 |
4 |
5 |
6 |
Basidiomycetes |
Filobasidiella neoformans |
Cryptococcusneoformans |
Почва, помет птиц, летучих мышей |
Вдыхание |
Криптококкоз |
Ascomycètes или |
Не обнаружена |
Madurella spp. |
Почва, растения |
Инокуляция |
Мицетома |
Deuteromycetes |
Ceratocystis stenoceras |
Sporotrix schenkii |
Растения,почва |
Инокуляция |
Споротрихоз |
Различные Dathideales |
Phialophora spp. |
Растения, почва |
Инокуляция |
Хромомикоз |
|
Различные Eurotiales |
Aspergillus fumigatus, другиевиды Aspergillus иPenicillium |
Распространены повсеместно |
Вдыхание, инокуляция |
Аспергиллез |
|
Arthrodermasрр. (Onygenales) |
Trichophyton spp. |
Почва, животные, человек |
Через кожу и волосы |
Дерматомикозы |
|
Arthroderma sрр. |
Micrisporum |
Почва, животные, человек |
Через кожу и волосы |
Дерматомикозы |
|
Различные Onygenales |
Epidermophytonspp. |
Почва, животные, человек |
Через кожу и волосы |
Дерматомикозы |
|
Ajellomyces capsulates |
Histoplasmacapsulatum |
Почва, растения |
Вдыхание |
Бастомикоз |
|
Не обнаружена |
Paracoccidioidesbrasiliensis |
Растения, почва |
Инокуляция |
Паракокцидиоидоз |
|
Не обнаружена |
Coccidioidesimmitis |
Почва, остатки растений |
Вдыхание |
Кокцидиоидоз |
|
Не обнаружена |
Candida albicans |
Животные, человек и их окружение |
Контакт, эндогенно |
Кандидоз |
|
Различные Endomycetes |
Другие виды Candida |
Животные, человек и их окружение |
Контакт, эндогенно |
Кандидоз |
|
Не обнаружена |
Malassezia furfur |
Человек |
Через кожу |
Разноцветный (отрубевидный) лишай |
|
Не обнаружена |
Pneumocystis carinii |
Человек |
Эндогенно |
Пневмоцистная пневмония |
|
Zygomycetes |
Различные Mucorales |
Распространены повсеместно |
Вдыхание |
Мукоромикоз |
|
Entomophthora coronata Rhinosporidium seeberi |
Водная среда, насекомые Вредая среда |
Энтомофтороз Риноспоридиоз |
Наряду с этим существуют и микозы, вызванные патогенными грибами. К ним относятся дерматомикозы и глубокие микозы (кокцидиоидоз, паракокцидиоидоз, бластомикоз, гистоплазмоз). Всего патогенных грибов насчитывают около 100 видов, тогда как грибов-оппортунистов — несколько сотен видов.
В таблице 10 приведены наиболее важные возбудители (этиологические агенты) микозов человека. Этиологический агент — либо один паразитический вид, либо группа близких микроорганизмов. Клиническая картина микоза сильно варьирует в зависимости от фонового заболевания, локализации очага или тяжести поражения. Например, аспергиллы могут вызывать поражение кожи, подкожных тканей, легких; грибы рода Candida — слизистых оболочек рта и половых органов, кожи, ногтей, бронхов, легких и других органов.
Большинство возбудителей микозов — космополиты. Заражение может происходить контактным путем, через одежду, обувь (дерматомикозы), от больных животных (микроспория, трихофития). Микозы стоп, от которых страдают примерно пятая часть населения Земли, передаются при хождении босыми ногами в банях, бассейнах и т. п. Некоторые микозы представляют собой раневые инфекции (мицетомы, споротрихоз, хромиомикоз, лобомикоз). Большинство глубоких микозов — респираторные инфекции (гистоплазмоз, бластомикоз, кокцидиоидоз, «плесневые» микозы).
Профилактика и терапия
Грибы, в том числе патогенные, окружают нас постоянно. Поэтому самый надежный способ профилактики — это здоровый образ жизни, способствующий укреплению иммунной системы, защищающей человека от чужеродных агентов. Сюда относится правильное питание, занятие спортом, прогулки на свежем воздухе, отказ от вредных привычек (курение, алкоголь).
Микозы ног можно предупредить, соблюдая личную гигиену. При работе в запыленных помещениях следует пользоваться респиратором.
Предрасполагающим фактором к развитию кандидоза является дисбактериоз и авитаминоз. Поэтому для его предупреждения используют препараты — эубиотики, содержащие полезные молочнокислые бактерии и витамины.
Методы терапии зависят от характера заболевания. При поверхностных микозах используют местные средства (производные имидазола, ундециленовой кислоты, нитрофенола и др.). При диссеминированных и генерализованных формах применяют антибиотики гризеофульвин, нистатин, амфотерицин В и др.
Производные имидазола (амиказол, изоконазол, кетоконазол, клотримазол, миконазол, сульконазол, тиоконазол, эконазол) и триазола (итраконазол, флуконазол) активны против мицелиальных грибов и дрожжей. Их часто применяют для лечения кандидозов.
При лечении микозов, вызванных видами Candida и Cryptococcus, используют 5-фторцитозин, первоначально предлагаемый как противоопухолевый препарат.
Производные аллиламина (батрафен, ламизил) эффективны в отношении дерматофитов, Candida albicans, но применяются, в основном, при лечении онихомикоза (грибковое поражение ногтей).
При диссеминированных и генерализованных формах применяют противогрибковые антибиотики, среди которых наибольшее распространение получили полиеновые (амфотерицин В, нистатин, пимарицин, леворин, фунгизон и др.) и гризеофульвин.
Амфотерицин В эффективен при лечении заболеваний, вызванных видами Candida, Aspergillus, Blastomyces, Coccidioides, Histoplasma. Другие полиеновые антибиотики (нистатин, пимарицин) имеют аналогичный амфотерицину. В спектр действия, но они более токсичны для млекопитающих.
Гризеофульвин является антибиотиком узкого спектра действия, он эффективен против дерматофитов, но не против дрожжевых грибов, в частности, рода Candida. Гризеофульвин вмешивается в митотический процесс при размножении грибов, воздействуя на синтез белка микротрубочек веретена.
Все эти средства применяются под контролем врача после установления диагноза заболевания. Диагностика заболевания требует установления его микотической природы методами микроскопии, а если это возможно — выделения чистой культуры возбудителя. Полезными оказываются и иммунологические методы (серодиагностика, кожные пробы). Данные лабораторных исследований должны соответствовать клинической картине заболевания, хотя симптомы микозов часто бывают неспецифичными.
Микозы животных
Домашние животные часто страдают от дерматофитии, вызываемой грибами родов Trichophyton и Microsporum, которые способны передаваться человеку. Среди возбудителей генерализованных микозов наиболее распространена Candida albicans, которая вызывает ощутимые потери в животноводстве и птицеводстве, особенно среди молодняка. Развитию кандидоза способствует применение антибиотиков в качестве профилактического средства для предупреждения бактериальных инфекций.
У грызунов выявлены системные заболевания кокцидиоидозом, криптококкозом, гистоплазмозом и споротрихозом.
Некоторые грибы поражают только животных и неизвестны у человека. Так Histoplasma farciminosum вызывает глубокий микоз у лошадей, мулов и ослов, а Pityrosporum pachydermatis — дерматомикоз у собак, коров, лошадей, свиней и носорогов. Грибы рода Saprolegnia (Oomycetes) паразитируют на рыбах.