ФАРМАЦЕВТИЧЕСКАЯ МИКРОБИОЛОГИЯ - В. А. Галынкин - 2015
ЧАСТЬ I. БИОЛОГИЯ МИКРООРГАНИЗМОВ
ГЛАВА I. ПРОКАРИОТЫ (БАКТЕРИИ)
1.1 Строение прокариотической клетки
Все живые организмы распределяются в трех сферах обитания: животный мир, растительный мир и мир простейших. На нашей планете насчитывается до 3 млн. видов животных и около полумиллиона видов растений. В 1886 г. немецкий биолог Э. Геккель предложил выделить все одноклеточные микроорганизмы (простейшие, грибы, бактерии), у которых отсутствует дифференцировка на органы и ткани, в отдельное царство — Protista (протисты, первосущества), включив в него организмы, во многих отношениях занимающие промежуточное положение между растениями и животными. В дальнейшем с учетом строения клеток протисты были подразделены на две четко разграниченные группы — низшие (прокариоты) (рис. 1) и высшие (эукариоты) (рис. 2).
Рис. 1. Схема строения прокариотической клетки [1].
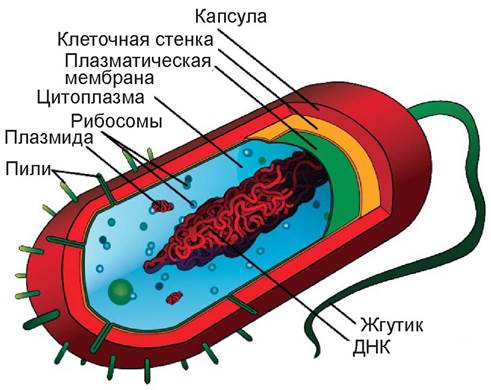
Рис. 2. Модель организации эукариотической клетки [1].
Величина клеток большинства прокариот находится в пределах 0,2-10,0 мкм (Таблица 1). Самыми крупными из до сих пор выделенных прокариот являются клетки Epulopiscium flshelsoni, обитающие в кишечнике глубоководной рыбы-хирурга, — до 600 мкм в длину и до 100 мкм в диаметре, и клетки Тhiomargarita namibiensis, найденные в прибрежных водах Чили и Намибии, — от 400 до 750 мкм в диаметре. В то же время есть очень маленькая морская водоросль Nanochlorum eukaryotum, имеющая, однако, настоящее ядро, хлоропласты и митохондрии. Резюмируя эти данные, можно сказать, что на сегодняшний день размеры известных прокариотических микроорганизмов колеблются от 0,05 до 750 мкм.
Таблица 1. Размеры отдельных представителей живого мира
Организм |
Размер, мкм |
Нанобактерии |
d<0,05 |
Поксвирусы |
d~0,3 |
Микоплазмы |
d = 0,3 |
Nanochlorum eukaryotum |
d~1,0-2,0 |
Е. coli |
1,1-1,5 x 2,0-6,0 |
Спирохеты |
1,5 x 50,0 |
Oscillatoria sp. |
d~7,0 |
Эритроцит |
d~7,0 |
Epulopiscium fishelsoni |
100,0x600,0 |
Thiomargarita namibiensis |
d~750,0 |
К низшим отнесены протисты, клетки которых по строению существенно отличаются от всех других организмов (бактерии и сине-зеленые водоросли), это — прокариоты(доядерные).
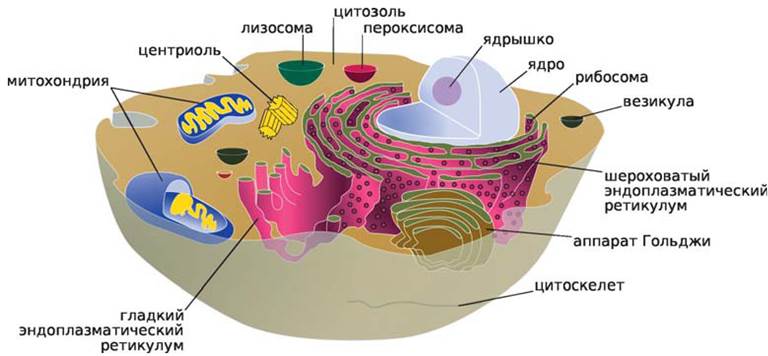
У высших протистов клетки сходны с растительными и животными клетками, это — эукариоты, т. е. микроорганизмы, имеющие истинное ядро (от греч. эу — истинный, карио — ядро). Ядро отделено от окружающей его цитоплазмы двухслойной ядерной мембраной с порами. В ядре находятся 1.. .2 ядрышка — центры синтеза рибосомальной РНК и хромосомы — основные носители наследственной информации, состоящие из ДНК и белка. При делении хромосомы распределяются между дочерними клетками в результате сложных процессов — митоза и мейоза. Цитоплазма эукариот содержит митохондрии, а фотосинтезирующих организмов — хлоропласты. Цитоплазматическая мембрана, окружающая клетку, переходит внутри цитоплазмы в эндоплазматическую сеть; имеется также мембранная органелла — аппарат Гольджи, как компонент цитоплазмы (рис. 2).
К эукариотам отнесены микроскопические водоросли (кроме сине-зеленых), микроскопические грибы (плесени и дрожжи).
Бактерии освоили самые разнообразные среды обитания: они живут в почве, пыли, воде, воздухе, на внешних покровах животных и растений и внутри организма. Их можно обнаружить даже в горячих источниках, где они живут при температуре около 60°С или выше. Численность бактерий трудно определить: в 1 г плодородной почвы может находиться до 100 млн., а в 1 см 3 парного молока — 3000 бактерий. Жизнедеятельность микроорганизмов имеет важное значение для всех остальных живых существ, т. к. бактерии и грибы разрушают органическое вещество и участвуют в круговороте веществ в природе. К тому же бактерии приобретают все большее значение в жизни людей, и не потому, что они вызывают различные заболевания, а потому, что их можно использовать для получения многих необходимых продуктов.
Прокариотические клетки устроены проще, чем эукариотические клетки. В них нет четкой границы между ядром и цитоплазмой, отсутствует ядерная мембрана; ДНК не образует структур, похожих на хромосомы эукариот, поэтому у прокариот происходят только процессы митоза. У большинства прокариот отсутствуют внутриклеточные органеллы, ограниченные мембранами, а также митохондрии и хлоропласты; рибосомы свободно лежат в цитоплазме.
К прокариотическим организмам отнесены синезеленые водоросли, бактерии, риккетсии, актиномицеты и микоплазмы [2, 3].
В настоящее время описано более 3,5 тыс. видов бактерий, но их число постоянно возрастает. Разобраться в этом поразительном многообразии возможно благодаря систематике. Систематика (от греч. systema — целое, составленное из частей; systematicus — упорядоченный) — наука о классификации организмов, их эволюционном родстве и взаимоотношениях друг с другом.
Классификация (от лат. classis — разряд, группа) — это распределение множества организмов на основе учета их общих признаков на классы, группы (таксоны); составная часть систематики.
Таксономия (от греч. taxis — расположение по порядку, закон) — теория классификации, систематизации живой природы.
Термины «систематика» и «таксономия» часто употребляют как синонимы, однако систематика представляет собой более широкое понятие.
Систематика включает в себя три самостоятельные составные части: классификацию, идентификацию и номенклатуру. Классификация, как уже упоминалось, — это распределение организмов на таксономические группы.
Идентификация — это определение принадлежности изучаемого организма к тому или иному таксону (классу, порядку, семейству, роду, виду и пр.).
Номенклатура — это свод правил присвоения названий таксонам и список этих названий. Номенклатура — это заключительный этап систематики после классификации, выполняет функции «информационного языка» и до некоторой степени независима от классификации.
До второй половины XIX в. классификация основывалась на внешних проявлениях организма — фенотипах (морфология, подвижность, окраска по Граму, наличие капсулы, способность образования эндоспор, культурально-биохимические свойства и некоторые другие признаки), так как наследственная структура организмов — генотипы — была еще недоступна для исследования.
Следовательно, традиционная, или классическая, систематика, основанная на изучении внешних, проявляющихся в процессе жизнедеятельности признаков, — целиком фенотипическая систематика (феносистематика). Расширение доступной исследователю информации о фенотипе и использование вычислительной техники для ее обработки привело к появлению нового направления — численной (числовой, или нумерической) таксономии.
Возникновение (в 50-х гг. XX в.) и успешное развитие молекулярной биологии способствовали становлению нового направления в систематике, названного отечественными учеными геносистематикой. Геносистематика, в отличие от феносистематики, занимающейся изучением множества признаков, базируется на исследовании только одного вещества — наследственного материала (ДНК) клетки, в котором запрограммировано индивидуальное развитие организма. Иначе говоря, геносистематика — это раздел систематики, предметом исследования которого являются генотипы, или генетические программы, созданные в процессе биологической эволюции на Земле. Разница между феносистематикой и геносистематикой заключается в том, что они принципиально отличаются объектами исследования.
В классификации родственных микроорганизмов используют следующие таксономические категории: царство (regnum), отдел (divisio), секция (section), класс (classis), порядок или отряд (ordo), семейство (familia), род (genus), вид (species). Название микроорганизмам присваивают в соответствии с правилами Международного кодекса номенклатуры бактерий.
В микробиологии, как и в биологии, для обозначения видов бактерий принята двойная (бинарная) номенклатура, предложенная еще в XVIII в. К. Линнеем. Согласно номенклатуре, название рода пишется латинскими буквами с прописной: первое слово обозначает родовую принадлежность микроба (какой- либо морфологический признак, фамилию ученого, открывшего этот микроб, и др.); второе слово — название вида — пишется со строчной буквы. Видовое название микроорганизма, как правило, представляет собой производное от существительного, дающего описание либо цвета колонии, либо источника обитания микроорганизма, вызываемого им процесса или болезни и других отличительных признаков. Например, Escherichia coli указывает, что микроб открыл Эшерих, coli — обитатель кишечника; Bacillus anthracis — микроб образует спору, anthracis — возбудитель сибирской язвы; Azotobacter — микроорганизм, фиксирующий атмосферный азот.
Основной номенклатурной единицей служит вид. В. Д. Тимаков (1973) дает ему следующее определение: «Вид — это совокупность микроорганизмов, имеющих единое происхождение и генотип, сходных по морфологическим и биологическим свойствам, обладающих наследственно закрепленной способностью вызывать в среде естественного обитания качественно определенные специфические процессы». Вид подразделяют на подвиды или варианты. Если при изучении выделенных бактерий обнаруживают отклонение от типичных видовых свойств, то такую культуру рассматривают как подвид. Существуют также и инфраподвидовые подразделения, обусловленные отклонением какого-либо небольшого наследственного признака: антигенного — серовар, биохимического — биовар, отношения к фагам — фаговар, патогенности — патовар и др. Введение в слова общей части «вар» (вариант) рекомендовано во избежание возможных недоразумений, ранее применявшийся термин «тип» использован для обозначения номенклатурного типа.
В микробиологии используют следующие термины: «чистая и смешанная культура», «клон» и «штамм». Под термином «культура» понимают микроорганизмы, выращенные на плотной или в жидкой питательной среде в условиях лаборатории. Культуру микроорганизмов, состоящую из особей одного вида, называют чистой культурой. Смешанной культуройназывают смесь неоднородных организмов, выросших в питательной среде при посеве исследуемого материала (молока, почвы, воды, патологического материала) или при попадании в питательную среду, засеянную одним видом микроба, еще и другого вида микроба из внешней среды. Клон — это культура, полученная из одной популяции клетки определенного вида микроба. Штамм — чистая культура определенного вида микроба, выделенная из того или иного объекта и отличающаяся от эталонного штамма незначительными изменениями свойств (например, чувствительностью к антибиотикам, ферментацией углеводов и др.).
Следовательно, прокариоты составляют отдельную классификационную группу микроорганизмов, они существенно отличаются от эукариотных микроорганизмов, к которым принадлежат грибы, растения, животные и человек (табл. 2). В литературе традиционно принято называть представителей прокариот «бактериями» [4]. В зависимости от особенностей клеточной оболочки бактерии подразделяют на четыре основные категории: (1) грамотрицательные бактерии, (2) грамположительные бактерии, (3) бактерии, лишенные клеточных стенок, (4) архебактерии. Первые две группы имеют ригидные клеточные стенки, жесткий каркас которых составляет пептидогликан муреин, содержащий мурамовую кислоту. К ним относится большинство возбудителей инфекционных заболеваний и сапротрофных микроорганизмов. Бактерии, лишенные ригидной клеточной стенки и поэтому не имеющие постоянной формы клеток, называют микоплазмами. Среди них имеются виды, патогенные для человека (Mycoplasma pneumoniae, M. hominis, M. fermentans), животных и растений.
Архебактерии не содержат муреина в клеточной стенке, что делает их устойчивыми к β-лактамным антибиотикам. По особенностям молекулярного строения они значительно отличаются от других прокариот. Преимущественно это почвенные или водные микроорганизмы, обитающие в экстремальных условиях (в среде с высоким содержанием солей, сильнокислой, при высокой температуре). Кроме того, среди них имеются симбионты в пищеварительном тракте теплокровных.
1.2 Морфология
Прокариоты — это одноклеточные микроорганизмы, диаметр клеток которых обычно составляет от 0,2 до 2 мкм (рис. 1). По форме клеток их подразделяют на три основные группы: сферические (кокки), цилиндрические (бактерии, бациллы) и спиралевидные (спириллы, спирохеты) (рис. 3).
Рис. 3. Представители мира прокариот.
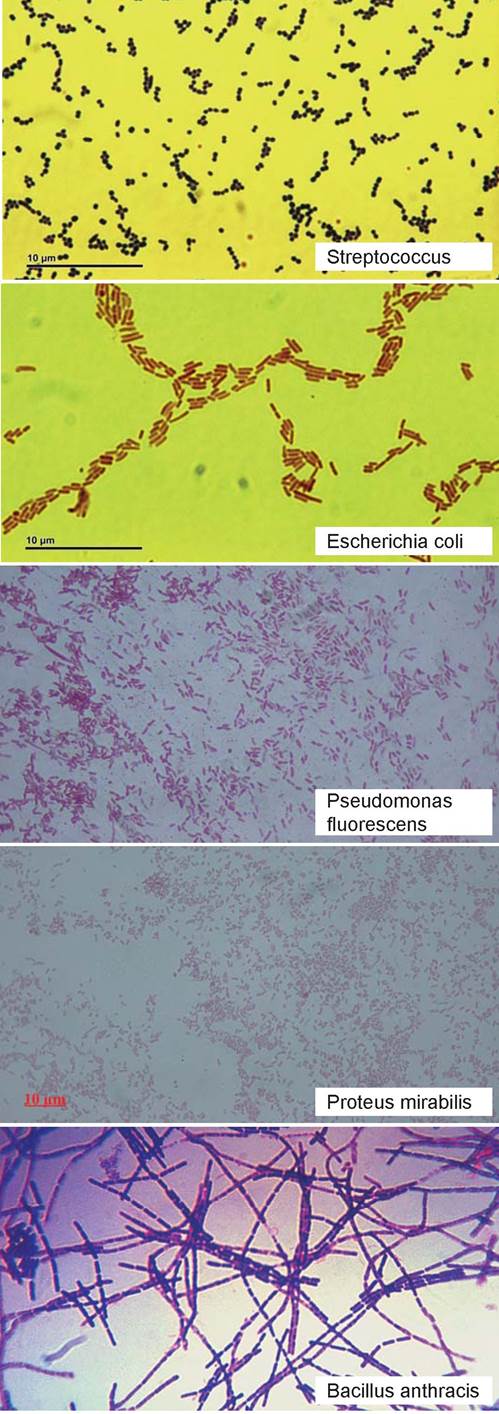
Таблица 2. Морфология прокариотов и эукариотов
Признак |
Прокариоты |
Эукариоты |
|
1. Цитологические свойства |
|||
Нуклеоид (нуклеоплазма, генофор) отделен от цитоплазмы мембраной |
— |
+ |
|
Диаметр клетки: |
|||
обычно 0,2-2 мкм |
+ |
— |
|
обычно > 2 мкм |
— |
+ |
|
Митохондрии |
- |
+ |
|
Хлоропласты (у фототрофов) |
- |
+ |
|
Вакуоли, если присутствуют, окружены мембраной |
- |
+ |
|
Аппарат Гольджи |
- |
D |
|
Лизосомы |
- |
D |
|
Эндоплазматический ретикулум |
- |
+ |
|
Локализация рибосом: |
|||
рассеяны в цитоплазме |
+ |
— |
|
прикреплены к эндоплазматическому ретикулуму |
— |
+ |
|
Ток цитоплазмы, эндоцитоз, экзоцитоз |
- |
D |
|
Диаметр жгутиков (если присутствуют): |
|||
0,01-0,02 мкм |
+ |
- |
|
около 0,2 мкм |
- |
+ |
|
Эндоспоры |
D |
- |
|
2. Химические признаки |
|||
Полигидроксибутират как запасное вещество в виде включений в цитоплазму |
D |
— |
|
Тейхоевые кислоты в клеточной стенке |
D |
- |
|
Полиненасыщенные жирные кислоты в составе мембран |
Редко |
Обычно |
|
Стеролы в составе мембран |
D |
Обычно |
|
Диаминопимелиновая кислота в составе клеточных стенок |
D |
- |
|
Пептидогликан, содержащий мурамовую кислоту, в составе клеточных стенок |
D |
- |
|
3. Метаболические свойства |
+ |
D |
|
Клетка поглощает питательные вещества в виде малых молекул Крупные молекулы или |
|||
частицы должны быть предварительно гидролизованы внеклеточными ферментами |
|||
Компоненты систем дыхания и фотосинтеза (у фототрофов) локализованы |
+ |
- |
|
в цитоплазматической мембране или ее инвагинантах |
|||
Хемолитотрофный тип метаболизма |
D |
- |
|
Фиксация N2 |
D |
- |
|
Способность к диссимиляционному восстановлению NO3 до N2O или N2 |
D |
- |
|
Метаногенез |
D |
- |
|
Аноксигенный фотосинтез |
D |
- |
|
Особенности размножения |
|||
Деление клеток без образования системы веретена |
+ |
— |
|
Деление клеток путем митоза с участием системы веретена |
- |
+ |
|
Мейоз |
- |
D |
|
Молекулярно-биологические свойства |
|||
Число хромосом в одном нуклеоиде или ядре |
Обычно 1 |
Обычно>1 |
|
Хромосомы кольцевые |
+ |
- |
|
Хромосомы линейные Рибосомы: |
|||
70 S |
+ |
— |
|
80 S |
— |
+ |
|
Рибосомальные РНК: |
|||
16 S, 23 S, 5 S |
+ |
— |
|
18 S, 28 S, 5,85 S, 5 S |
— |
+ |
|
По форме клеток их подразделяют на три основные группы: сферические (кокки), цилиндрические (бактерии, бациллы) и спиралевидные (спириллы, спирохеты) (рис. 4). Кокки по своей форме могут быть сферическими, эллипсовидными, бобовидными и ланцетовидными. По расположению клеток различают
диплококки, стрептококки, тетракокки, сардины, стафилококки.
Рис. 4. Морфология бактерий. А. Сферические клетки, располагающиеся группами (1 — Staphylococcus), парами (2, 3 — Neisseria), цепочками (4 — Streptococcus), тетрадами (5 — Micrococcus), кубическими пакетами (6 — Sarcina). В. Цилиндрические (палочковидные) клетки. С. Вибрионы. D. Спириллы. Е. Спирохета — схема строения: а — наружная мембрана, b — протоплазматический цилиндр, с — периплазматические жгутики [2].
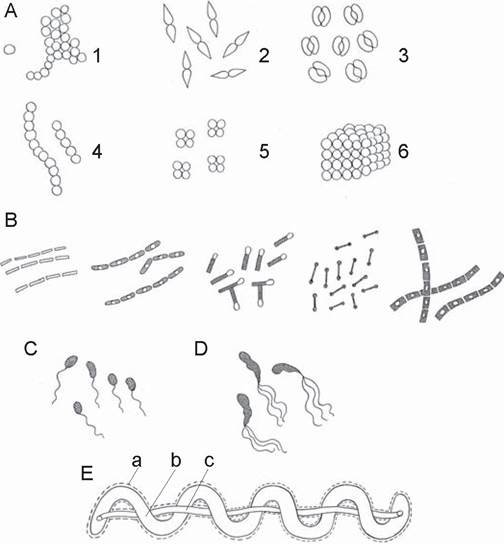
Микрококки (micrococcus) характеризуются одиночным, парным или беспорядочным расположением клеток. Род включает условно-патогенные (M. luteus) и сапротрофные виды.
Диплококки образуют пары. К ним относятся возбудители менингита и гонореи.
Стрептококки (streptos — извитой) располагаются цепочками. Среди них имеются патогенные виды (S. pyogenes).
Тетракокки располагаются по четыре, сарцины (sarcio — соединяю) образуют пакеты по 8, 16 и более клеток. В основном это сапротрофные виды.
Стафилококки (staphylos — гроздь) образуют скопления в виде виноградной грозди. Среди них имеются патогенные (S. aureus) виды.
Цилиндрические (палочковидные) формы микроорганизмов включают бактерии (палочки, не образующие споры), бациллы и клостридии (спорообразующие формы). Они различаются по размерам, форме и расположению клеток. Среди них имеются представители нормальной микробиоты человека (Escherichia coli) и патогенные виды (возбудители кишечных инфекций, столбняка, сибирской язвы, анаэробной инфекции и др.
Спиралевидные формы включают спириллы, имеющие жесткую клеточную стенку, и спирохеты, клеточная стенка которых эластична и обеспечивает их подвижность. Spirillum minor — возбудитель содоку — болезни, передающейся через укус грызунов. Спирохеты — возбудители многих инфекционных заболеваний (см. ниже).
Кроме указанных основных форм, среди прокариот имеются виды, отличающиеся более сложным строением. Это актиномицеты, к которым относится большинство продуцентов антибиотиков и некоторые патогенные виды.
1.3 Актиномицеты
Актиномицетов относят к грамположительным бактериям. С бактериями их сближает отсутствие настоящего ядра (прокариоты), их относят к мицелиальным прокариотам. Однако по морфологическим, физиологическим, биологическим и экологическим признакам актиномицеты составляют самостоятельную группу. Вегетативное тело актиномицетов (мицелий) представлено очень тонкими (в 5-7 раз тоньше, чем грибные, ветвящимися гифами (рис. 5).
Рис. 5. Актиномицеты. Схема роста воздушного и субстратного мицелия родов: 1 — Nocardia, 2 — Actinoplanes, 3-5 — Streptomyces, 6 — Streptoverticillium [2].
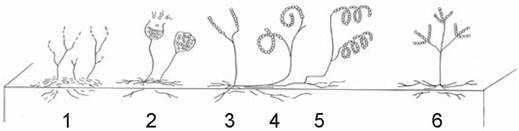
Размножаются актиномицеты участками мицелия или спорами. На питательных средах актиномицеты образуют сначала кожистые колонии (субстратный мицелий), которые затем покрываются воздушным мицелием. Сама колония врастает в агар субстратным мицелием. Питание актиномицетов не специализировано. Они используют различные животные и растительные остатки.
Большинство актиномицетов ведут сапротрофный образ жизни, и только некоторые из них приспособились к паразитическому существованию на растениях.
Среди патогенных актиномицетов наибольший интерес представляют виды рода Streptomyces, вызывающие паршу у растений. Наиболее известны обыкновенная парша клубней картофеля и парша корнеплодов свеклы, моркови.
Паршу картофеля вызывает Streptomyces scabies. Заболевание развивается на клубнях во время вегетации растения. В местах заражения появляются трещины, небольшие бородавки, происходит опробковение пораженной ткани, образуются язвы.
Оксигенные фототрофные бактерии (цианобактерии) — обитатели воды и почвы, вызывают цветение водоемов, некоторые виды образуют токсины, опасные при употреблении цветущей воды. Могут служить источником биологически активных веществ (фикоцианин) или употребляться как пищевая добавка (Spirulina maxima).
В микробиологии существуют два различных подхода к систематике, обусловливающие два вида классификации. В основе первого лежит идея создания естественной (филогенетической) классификации прокариот, т. е. построения единой системы, объективно отражающей родственные отношения между разными группами микробов и историю их эволюционного развития. Второй подход к систематике преследует практические цели и служит для идентификации, т. е. установления принадлежности микроорганизма к определенному виду. Это искусственная классификация (традиционная). Современные системы классификации микроорганизмов, по существу, все искусственные. На их основе созданы определители для идентификации того или иного микроорганизма: «Определитель бактерий и актиномицетов» Н. А. Красильникова (1949).
1.4 Классификация прокариотических микроорганизмов
В определителе бактерий Д. X. Берджи [3] все прокариотические микроорганизмы объединены в царство Procaryotae, которое подразделяется на четыре отдела, которые делятся на секции, классы, порядки, семейства, роды, виды.
Отдел I. Gracilicutes (от лат. gracilus — тонкий, стройный, cutes — кожа). Включает в себя грамотрицательные микроорганизмы. В отделе девять секций.
Секция 1. Спирохеты. Порядок Spirochaetales. Включает в себя два семейства: Spirochaetaceae (четыре рода), Leptospiraceae (один род).
Секция 2. Спиралевидные и изогнутые аэробы (микроаэрофилы). Одно семейство — Spirillaceae, в котором шесть родов. Патогенны для человека и животных микроорганизмы рода Campylobacter.
Секция 3. Грамотрицательные неподвижные изогнутые бактерии. Одно семейство — Spirosomonaceae, в котором патогенных три рода.
Секция 4. Аэробные грамотрицательные палочки, округлые и кокки. Восемь семейств, два из которых имеют патогенные микроорганизмы. Семейство Pseudomonadaceae включает в себя четыре рода, более 25 видов, среди которых имеются патогенные (A. mallei и др.). Семейство Neisseriaceae имеет 16 родов. Роды Neisseria и Moraxella содержат патогенные для человека и животных микроорганизмы.
Роды Bordetella, Brucella и Francisella не внесены в семейства: содержат патогенные для человека и животных микроорганизмы.
Секция 5. Грамотрицательные факультативные анаэробы. Три семейства: Enterobacteriaceae, Vibrionaceae и Pasteurellaceae. Семейство Enterobacteriaceae имеет 14 родов(Escherichia, Salmonella, Citrobacter, Klebsiella, Enterobacter, Erwinia, Shigella, Proteus, Yersinia и др.). Семейство Vibrionaceae имеет два рода. В род Vibrio включены патогенныемикроорганизмы. Семейство Pasteurellaceae имеет три основных рода: Pasteurella, Haemophilus и Actinobacillus. Содержат патогенные виды микроорганизмов.
Секция 6. Строгие анаэробы. Изогнутые грамотрицательные палочки. Одно семейство — Bacteroidaceae, в котором 13 родов, среди которых имеются патогенные.
Секция 7. Диссимилирующие и разлагающие сульфат бактерии. Семь непатогенных родов.
Секция 8. Анаэробные грамотрицательные кокки. Одно семейство — Vellonellaceae, в котором три рода.
Секция 9. Риккетсии и хламидии. Два порядка: Rickettsiales и Chlamydiales. Порядок Rickettsiales имеет три семейства: Rickettsiaceae, Bartonellaceae и Anaplasmataceae. Семейство Rickettsiaceae имеет три трибы, в которые внесено восемь родов. Семейство Bartonellaceae содержит два рода, a Anaplasmataceae — четыре. Порядок Chlamydiales имеет одно семейство Chlamydiaceae и один род — Chlamydia. Все семейства содержат патогенные микроорганизмы.
Отдел II. Firmicutes (от лат. firmis — крепкий, cutes — кожа). В отдел включены главным образом грамположительные бактерии.
Секция 12. Грамположительные кокки. Два семейства: Micrococcaceae и Deinococcaceae. Семейство Micrococcaceae имеет четыре рода: Micrococcus, Stomatococcus, Planococcus, Staphylococcus.
В секцию кроме указанных двух семейств внесены десять самостоятельных родов: Streptococcus, Leuconostos, Pedicoccus, Sarcina и др.
Секция 13. Спорообразующие грамположительные палочки и кокки. Шесть родов: Bacillus, Clostridium, Sporolactobacillus, Sporosarcina и др. Первые два рода имеют патогенные виды.
Секция 14. Неспорообразующие грамположительные палочки. Семь родов: Lactobacillus, Listeria, Erysipelotrix и др. Имеются патогенные.
Секция 15. Неспорообразующие внутриклеточные грамположительные палочки. 21 род:
Corynebacterium, Micobacterium, Propionibacterium, Eubacterium, Asotobacterium, Bifidobacterium, Actinomices и др.
Секция 16. Микобактерии. Одно семейство Mycobacterioœae. Семейство имеет один род Mycobacterium, в котором 49 видов: Мус. tuberculosis, Мус. bovis, Мус. avium, Мус. paratuberculosis, Мус. lepra и др.
Секция 17. Nocardioforms. Девять родов: Nocardia, Pseudococcus, Pseudonocardia и др.
Отдел III. Tenericutes. Объединены грамотрицательные прокариоты без клеточной стенки, но имеющие цитоплазматическую мембрану. В отделе десятая секция — микоплазмы, класса Mollicutes (от лат. molli — мягкий, cutes — покров, кожа). В классе один порядок — Mycoplasmatales — и три семейства: Mycoplasmataceae, Acholeplasmataceae, Spiroplasmataceae. В основном патогенные микоплазмы включены в семейство Mycoplasmataceae.
Секция 11. Эндосимбионты.
Отдел IV. Mendosicutes. Прокариоты, среди которых нет патогенных бактерий; метанобразующие, сероокисляющие, галофилы, микоплазмоподобные, термоацидофильные и другие наиболее древние по происхождению бактерии (архебактерии).
1.5 Строение клетки
Клеточная стенка обеспечивает поддержание жесткости структуры клетки, постоянства ее формы и механической прочности. Кроме того, она является осмотическим барьером, имеющим зоны избирательной проницаемости для веществ различной химической природы. В качестве опорного каркаса она содержит пептидогликан муреин. Основу муреина (рис. 6) составляют цепи чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенные β-1,4-гликозидными связями. Остатки мурамовой кислоты соединены полипептидными цепочками, в состав которых входят α-аланин, D-аланин, лизин, D-глутаминовая и мезодиаминопимелиновая кислоты.
Рис. 6. Пептидогликан E. coli. A-N-ацетилмурамовая кислота; B-N-ацетил- глюкозамин; ala — аланин; glu — глутаминовая кислота, DAP — диаминопимелиновая кислота.
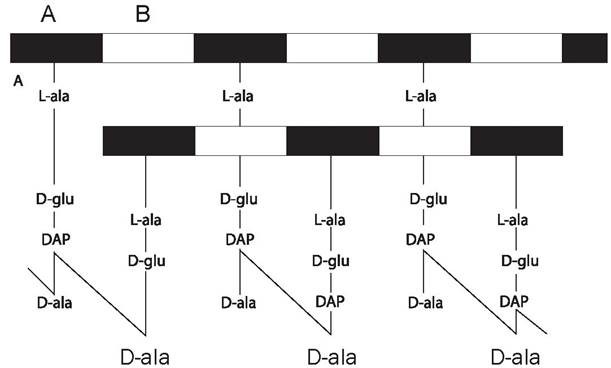
Аминокислоты D-ряда и мурамовая кислота уникальны для прокариот, своеобразие структуры клеточной стенки служит основой избирательного действия некоторых антибиотиков, например пенициллина и других β-лактамов. Клеточные стенки грамположительных и грамотрицательных бактерий существенно различаются по своей структуре.
У грамотрицательных бактерий муреиновая сеть однослойная, иногда двуслойная и составляет не более 10% сухой массы клеточной стенки. На ней располагаются белки, липопротеиды, липополисахариды и фосфолипиды, входящие в состав внешней мембраны (рис. 7). Стабилизация этих компонентов обеспечивается ионами Са2+ и Mg2+. Существенное значение для структуры и функции внешней мембраны имеет липид А. Его скелет содержит дисахарид, состоящий из остатков D-глюкозамина, соединенных β-1,6-связью, имеющих в положении 1 и 4 фосфатные группы. Скелет этерифицирован жирными кислотами С12, С14 и С16. Липид А имеет уникальную конформацию — компактную и высокоупорядоченную, благодаря чему создает в мембране вязкую структуру, которая затрудняет диффузию желчных кислот, детергентов и некоторых антибиотиков. Липид А обеспечивает токсичность и пирогенность липополисахарида. Антигенная специфичность грамотрицательных бактерий, главным образом, определяется углеводами О-замещенных, боковых цепей, выступающих наружу с поверхности клетки. Внешняя мембрана определяет высокую устойчивость грамотрицательных бактерий по сравнению с грамположительными к антимикробным агентам.
Рис. 7. Модель строения цитоплазматической мембраны грамотрицательных бактерий: Справа представлена липополисахаридная молекула. Глю — глюкоза; Глю-N — глюкозамин; NА = N-АцГлю-N-ацетилглюкозамин; Гал — галактоза; Геп — гептоза; КДО — 2-кето- 3-дезоксиоктоновая кислота; М — муреин; НМ — наружная мембрана; ПМ — плазматическая мембрана; ПП — периплазматическое пространство [7].
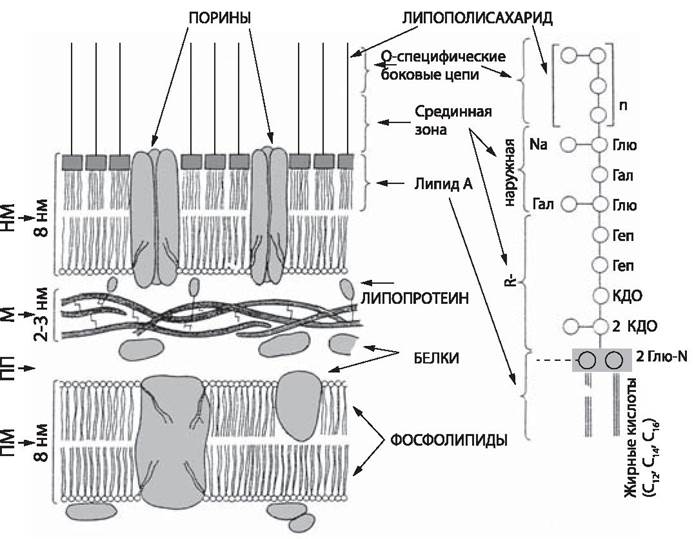
Антибиотики широкого спектра, например, бета- лактамы, вызывают освобождение липополисахаридов из внешней мембраны, что может привести к эндотоксическому шоку у больного.
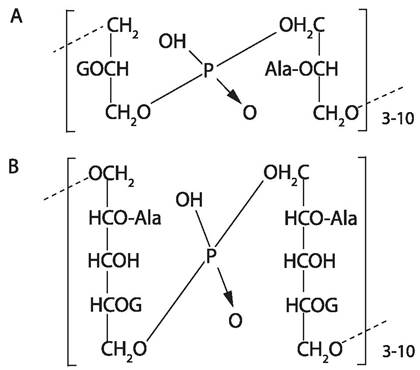
У грамположительных бактерий внешняя мембрана отсутствует, а муреиновая сеть составляет 30-70% сухой массы клеточной стенки и достигает 40 слоев. Характерно наличие тейхоевых и тейхуроновых кислот (рис. 8), обеспечивающих отрицательный заряд клетки и способствующих сорбции катионов из окружающей среды. У некоторых микроорганизмов могут присутствовать добавочные компоненты — липиды, воска, миколовые кислоты, протеины, полисахариды.
Рис. 8. Модель строения цитоплазматической мембраны Гр+ бактерий. А — глицеролтейхоевая кислота; В — рибитолтейхоевая кислота G — гликозил, Ala — аланил.
Различие в структуре клеточной стенки двух групп микроорганизмов выявляют с помощью окрашивания по Граму. Препарат обрабатывают раствором кристаллического фиолетового, затем йода. Образующийся комплекс красителя с йодом располагается на протопласте. При обработке препарата спиртом он удерживается клеточной стенкой грамположительных бактерий и вымывается — у грамотрицательных. Способность окрашиваться по Граму — важный таксономический признак, с которым коррелируют другие свойства бактерий.
S-слой (surface — поверхность) располагается на поверхности клеток всех прокариот и покрывает целиком всю клетку. Он состоит из структурных единиц — протеинов или гликопротеинов, образующих монослой, структура которого типична для двухмерных кристаллов (решетка гексагональной, косой или квадратной симметрии). Взаимодействие между субъединицами и подлежащими структурами происходит за счет нековалентных связей. S-слой обеспечивает защиту клетки от внешних воздействий, однако при продолжительном культивировании он может быть утрачен без потери жизнеспособности штамма.
Капсулы и слизь образуются у некоторых бактерий снаружи от клеточной стенки, как ее внешний слой. Способность к их формированию не является видовым признаком: могут существовать капсульные и бескапсульные штаммы. У патогенных микроорганизмов капсула обеспечивает защиту от фагоцитоза, повышая вирулентность штамма (у пневмококков). У микробов, обитающих в почве и на растениях, капсула защищает клетки от высыхания, солнечной радиации, биоцидов. Капсулы и слизь создают для микробных клеток осмотические условия, благоприятствующие сорбции питательных веществ из субстрата, способствуют адгезии клеток между собой и субстратом. Многие экзоферменты локализуются в капсуле, где происходят превращения веществ, поступающих в клетку (рис. 9).
Рис. 9. Капсулы вокруг клеток Clostridium.
У большинства бактерий капсулы и слизь имеют полисахаридную природу. У некоторых бацилл это полипептиды в основном D- и L-глутаминовой кислоты.
Капсульные полисахариды обладают антигенной специфичностью и используются для изготовления вакцин (у пневмококков, менингококков), для идентификации и классификации (у сальмонелл). Растворимые слизи (декстран Leuconostoc dextranicum, L. mesenteroides, ксантан Xanthomonas campestris) получают в промышленных масштабах и широко используют в фармации и других областях.
Протопласты и сферопласты — это структуры, полностью (протопласты) или частично (сферопласты) утратившие клеточную стенку, например, под действием лизоцима или пенициллина. Это осмотически лабильные элементы, которые могут существовать только в гипертонических растворах. Они сохраняют биологическую активность и способны в специальных условиях ревертировать в нормальные клетки. Используются в клеточной инженерии для получения гибридных форм микроорганизмов.
L-формы, получившие свое название в честь института Листера в Лондоне, образуются в условиях, приводящих к нарушению синтеза клеточной стенки; например, у больного туберкулезом возбудитель под влиянием лекарственных веществ может превратиться в L-форму. При этом микобактерии теряют характерную кислотоустойчивость, что затрудняет их выявление и диагностику заболевания. Для таких клеток характерны неправильные формы, иногда нитевидные, способные проходить через поры бактериальных фильтров. Лабильные L-формы способны ревертировать в нормальные клетки. Стабильные формы не образуют клеточной стенки, поскольку ее утрата связана с изменением генотипа (мутацией).
Периплазматическое пространство располагается между слоем муреина и цитоплазматической мембраной. В нем находятся ферменты гидролазы, расщепляющие вещества, поступающие в клетку, и полимеразы, участвующие в синтезе клеточной стенки и капсулы, а также белки, принимающие участие в транспорте субстратов в цитоплазму, и белки — рецепторы хемотаксических стимулов.
Цитоплазматическая мембрана располагается под клеточной стенкой и отделяет от внешней среды цитоплазму. Имеет толщину 6-8 нм и составляет 8-15% сухого вещества клетки. Ее структура соответствует общему принципу организации мембран про- и эукариотических клеток. Она состоит из двух слоев молекул липидов, у которых гидрофобные цепочки жирных кислот ориентированы перпендикулярно ее плоскости, а гидрофильные полярные части молекул соединены с молекулами белков за счет полярного, электростатического и гидрофобного взаимодействия. Молекулы липидов и белков не сохраняют фиксированной ориентации, но находятся в постоянном движении внутри остова данной мембраны (рис. 10)
Рис. 10. Структура бактериальной клеточной стенки. Вверху: грамположительная клеточная стенка. 1. Цитоплазматическая мембрана, 2 — ПГ, 3 — фосфолипид, 4 — протеин, 5 — липотейхоевая кислота. Внизу: грамотрицательная клеточная стенка. 1 — внутренняя мембрана, 2 — периплазматическое пространство, 3 — внешняя мембрана, 4 — фосфолипид, 5 — ПГ, 6 — ЛП, 7 — протеин, 8 — ЛПС, 9 — порины.
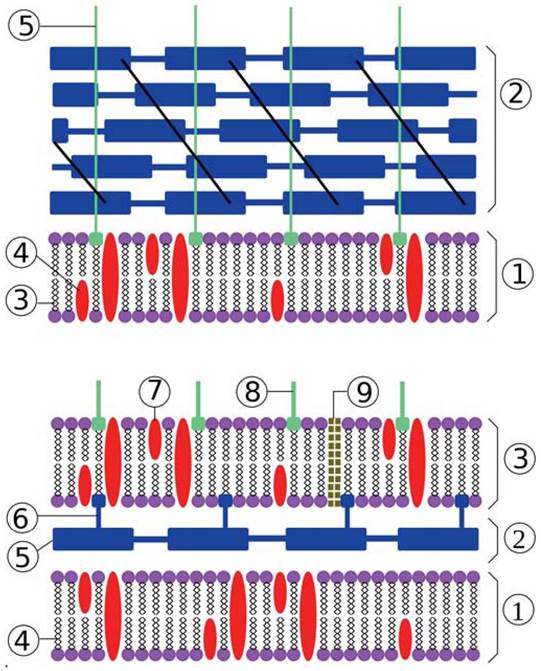
Структура грамположительной клеточной стенки. Клеточная стенка содержит до 40 слоев пептидогликана. Молекулы тейхоевых кислот ковалентно связаны с пептидогликаном (ПГ). Липотейхоевые кислоты (ЛТК) содержат липидные «хвостики», закрепленные в гидрофобной области ЦПМ. Клеточные стенки могут иметь белковые слои на поверхности. Белковые структуры располагаются или островками (как показано на схеме), или тесно упаковываются, образуя S-слой.
Структура грамотрицательной клеточной стенки. Тонкий слой пептидогликана сверху покрыт внешней мембраной (ВМ), прикрепленной к нему липопротеидами (ЛП). Между двумя мембранами — периплазматическое пространство со слоем пептидогликана (ПГ) внутри него. Внешняя мембрана на внутренней поверхности содержит фосфолипиды, на внешней — липополисахариды (ЛПС). ЛПС состоят из липидной части, обращенной внутрь внешней мембраны, формируя ее гидрофобную область, и полисахаридной части, обращенной во внешнюю среду.
Липиды, в основном, полярные фосфолипиды, составляют около 40% массы мембраны. Они выполняют структурные функции и обеспечивают конформационные изменения молекул ферментов, необходимые для проявления их активности.
Белки мембран подразделяют на периферические и интегральные. Первые находятся на поверхности, вторые погружены в толщу мембраны. Белки-порины выстилают поры мембран, обеспечивая ее проницаемость. Определенные белки мембран выполняют каталитические функции или являются рецепторами, связывающими вещества, необходимые для клетки.
Цитоплазматическая мембрана служит осмотическим барьером, в ней локализуются системы активного транспорта веществ в клетку и из клетки.
Мезосомы — особые структуры, образуемые путем инвагинации (впячивания) мембраны, содержат ферменты системы окислительного фосфорилирования и выполняют у прокариот функции митохондрий.
Цитоплазма составляет внутреннюю среду клетки. Это сложная высокогетерогенная система, в ней располагается генетический материал клетки (нуклеоид, плазмиды), 70 Sрибосомы, ферментные системы, выполняющие метаболические функции, резервные вещества (полисахариды, липиды, полигидроксимасляная кислота, полифосфаты, сера у серобактерий). Основная часть метаболических процессов осуществляется в цитоплазме. Мембранные структуры, располагающиеся в цитоплазме, наиболее развиты в клетках эукариот, у бактерий имеются их аналоги (мезосомы, вакуоли, лизосомы).
Жгутики предназначены для передвижения бактерии (рис. 11).
Рис. 11. Органоиды движения у микроорганизмов: А — Tetrahymena thermophila; Б — строение жгутика бактерий.
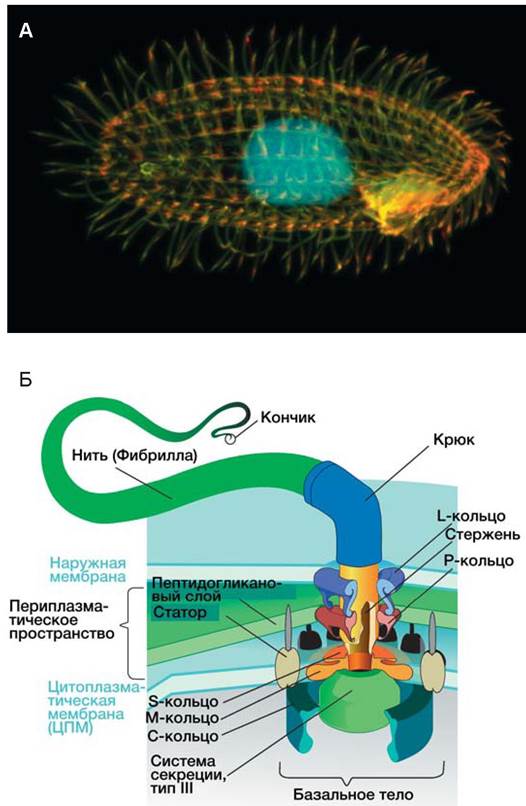
Перемещаться без жгутиков способны цианобактерии, скользящие бактерии и спирохеты. Число жгутиков и их расположение на клетке — таксономический признак, характерный для определенных видов.
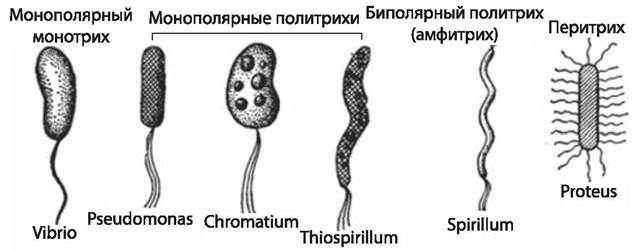
Монотрихи — бактерии с одним жгутиком на конце (Vibrio cholerae); амфитрихи имеют полярно расположенные жгутики на двух концах (Spirillum volutans); лофотрихи — пучок жгутиков на одном конце (Alcaligenes faecalis); у перитрихов жгутики расположены по всей поверхности клетки (E. coli, Salmonella spp.) (рис. 12).
Рис. 12. Основные типы расположения жгутиков бактерий [4]
Жгутики построены из белка — флагеллина, их диаметр 10-20 нм, длина до 20 мкм. Жгутик закреплен в цитоплазматической мембране и клеточной стенке с помощью базального тельца, состоящего из центрального стержня и двух пар (у грамотрицательных бактерий) и одной пары (у грамположительных) дисков. Жгутики вращаются благодаря тому, что через диски проходит поток заряженных частиц (Н+, ОН-, Nа+) за счет разности потенциалов внутри и вне клетки. Жгутики находятся под контролем системы, воспринимающей информацию о состоянии окружающей среды. Поэтому они позволяют клеткам перемещаться в область с оптимальными условиями (таксис). Существует около 35 генов, участвующих в сборке и функционировании базального тельца и еще более 20 генов, определяющих направление движения в ответ на внешние стимулы. Вся система находится под контролем главного оперона, который регулируется системой цАМФ — белок-активатор.
Фимбрии (пили или волоски) располагаются на поверхности клеток многих бактерий. Их число на клетке может доходить до 10 000. Их диаметр 3-25 нм, длина около 12 мкм. Они обеспечивают сцепление клеток, например, при конъюгации, их адгезию к субстрату. Белки фимбрий служат рецепторами, обеспечивающими биологическое узнавание. Некоторые фимбрии являются капиллярами, связанными с мезо- сомами, и участвуют в водно-солевом обмене.
F-волоски (F-пили) по структуре напоминают фимбрии, однако на клетке их не более одного-двух. Они находятся лишь у клеток, способных к передаче генетического материала при конъюгации, и принимают участие в этом процессе как рецепторы и связывающие структуры. Кроме того, они являются рецепторами специфических фагов.
Споры прокариот — это особая структура, предназначенная для сохранения в неблагоприятных условиях. Споры по сравнению с вегетативными клетками намного устойчивее к воздействию высокой температуры, радиации, химических агентов. Споры образуются внутри бактериальной клетки обычно при истощении в культуральной среде питательных веществ и накоплении продуктов обмена. Биохимическим сигналом для спорообразования служит снижение концентрации в клетке гуаниловых нуклеотидов — ГТФ и ГДФ. Спорообразование зависит от плотности популяции: при малой концентрации клеток споры не образуются. Спорообразование восстанавливается, если к такой культуре добавить фильтрат поздней экспоненциальной фазы культуры, в которой клетки начали спорулировать (см. чувство кворума). Образование спор требует синтеза ДНК: блокада репликации прекращает экспрессию ранее индуцированных генов споруляции. Образование споры начинается с накопления белкового материала, при этом расходуются запасные питательные вещества. Белки споры содержат значительно больше цистина, чем вегетативные клетки. Предполагают, что многочисленные дисульфидные связи в белке обеспечивают высокую механическую прочность оболочек спор. На ранней стадии споруляции образуются особые белки, которые связываются с ДНК, слегка раскручивая ее, что изменяет геометрию пиримидиновых оснований и повышает их устойчивость к ультрафиолетовому излучению. В период споруляции образуется специфическое вещество — дипиколиновая (пиридин- 2,6-дикарбоновая) кислота, которая в виде соли кальция входит в состав оболочки споры.
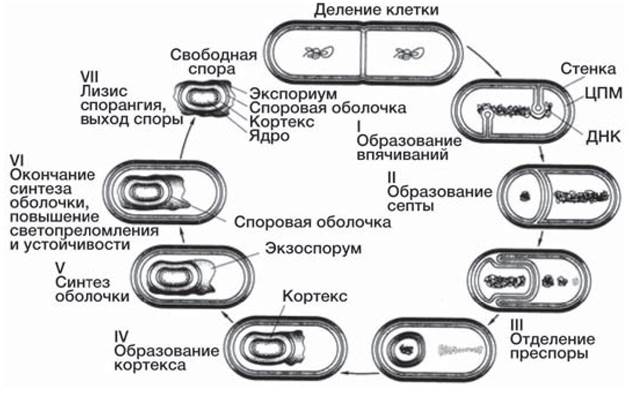
Морфологические изменения клетки в процессе споруляции (рис. 13) начинаются с образования асимметрично расположенной септы. Подобно обычной септе, образующейся при делении клеток, она состоит из двух мембран с тонким слоем пептидогликана между ними. Большая часть клетки является материнской, а меньшая — проспорой, содержащей нуклеоид. Далее септа деградирует, а мембрана материнской клетки обволакивает проспору, которая таким образом получает двойную мембрану. Между этими мембранами располагается слой пептидогликана, формирующий кортекс споры. На следующей стадии спорообразования происходит сборка белковых элементов оболочки споры. По окончании созревания споры материнская клетка лизируется. Зрелая спора содержит минимальное количество свободной воды и повышенное по сравнению с вегетативной клеткой количество липидов. На долю ее оболочки приходится до 50% сухой массы. Все эти особенности обеспечивают ее устойчивость к факторам внешней среды.
Спорообразование присуще преимущественно палочковидным микроорганизмам (бациллам, клостридиям). К ним относятся возбудители сибирской язвы, столбняка, анаэробной инфекции, ботулизма и некоторые сапротрофные виды. Помимо этого, споры образуют виды родов Thermoactinomyces, Sporosarcina, Sporomusa. Все они за исключением последнего по Граму окрашиваются положительно.
Спора — это покоящаяся форма. В благоприятных условиях споры прорастают. При этом спора набухает, поглощая воду, возрастает ее метаболическая активность, выделяется дипиколиновая кислота, спора утрачивает свою устойчивость. Наконец, оболочка споры разрывается, и из нее выходит вегетативная клетка (рис. 13).
Рис. 13. Цикл развития Bacillus subtilis: 1 — вегетативная клетка; 2 — образование септы споруляции; 3-4 — образование двойной мембраны споры; 5 — формирование кортекса (а); 6 — формирование белковой оболочки (б); 7 — зрелая спора; 8 — прорастание споры [2].
Способность микроорганизмов к спорообразованию учитывают при выборе методов дезинфекции и стерилизации, имея в виду высокую устойчивость спор к биоцидным агентам. Наиболее устойчивые виды используют в качестве тест-культур для оценки эффективности стерилизации: Bac. stearothermophilus — паром под давлением, некоторые виды Вас. subtilis — сухим паром, Bac. pumilus — радиационной.
Клетки прокариот (от греч. pro — до, karion — ядро) не имеют оформленного ядра. Иными словами, генетический материал (ДНК) прокариот находится прямо в цитоплазме и не окружен ядерной мембраной.
1.6 Генетический аппарат
Бактериальная хромосома представляет собой замкнутую кольцевую ДНК длиной около 1,3 м.
ДНК бактерий представлена одиночными кольцевыми молекулами длиной около 1 мм. Каждая такая молекула состоит примерно из 5 х 106 пар нуклеотидов. Суммарное содержание ДНК (геном) в бактериальной клетке намного меньше, чем в эукариотической, а, следовательно, меньше и объем закодированной в ней информации. В среднем, такая ДНК содержит несколько тысяч генов, что примерно в 500 раз меньше, чем в клетке человека.
Плазмиды и эписомы
Плазмиды и эписомы — это небольшие фрагменты ДНК, отличающейся от основной массы ДНК. Они часто реплицируются вместе с ДНК хозяина, но не нужны для выживания его клетки.
Сначала было принято различать эписомы и плазмиды: эписомы внедряются в ДНК хозяина, а плазмиды — нет. К эписомам относятся F-факторы и так называемые умеренные фаги. Сейчас обе группы называют одним общим термином «плазмиды». Плазмиды широко распространены в природе, и в последние годы их считают внутриклеточными паразитами или симбионтами, устроенными еще проще, чем вирусы. Вопрос о том, можно ли вирусы считать живыми организмами, будет подробно рассмотрен далее. Что касается плазмид, то здесь дело обстоит еще сложнее — ведь они представляют собой только молекулы ДНК. Плазмиды контролируют свою репликацию и число копий в клетке, которое у разных плазмид может колебаться от 1 до 100. Для плазмид характерно явление несовместимости, т. е. неспособности близкородственных плазмид существовать в одной клетке; на этом основана их классификация. Основная таксономическая единица у плазмид — Inc-группа (incompatibility — несовместимость). Плазмиды, относящиеся к одной 1 пс-группе, обладают многими общими свойствами: сходной молекулярной массой, высокой степенью гомологии ДНК — и контролируют сходные фенотипические признаки.
Механизм несовместимости состоит в угнетении репликации, проникшей в клетку плазмиды специфическим белком или РНК, вырабатываемым под контролем плазмиды, имеющейся в клетке. Кроме того, под контролем последней на поверхности клетки синтезируется специфический белок или липопротеин, препятствующий вхождению в клетку родственной плазмиды.
Плазмиды придают своим клеткам-хозяевам целый ряд особых свойств. Некоторые плазмиды являются «факторами резистенции» (R-плазмиды, или R-факторы: от англ. R = resistance — устойчивость), т. е. факторами, придающими устойчивость к антибиотикам. Примером может служить пенициллиназная плазмида стафилококков, которая трансдуцируется различными бактериофагами. В этой плазмиде содержится ген, кодирующий фермент пенициллиназу, которая разрушает пенициллин и, таким образом, придает устойчивость к пенициллину. Передача и распространение таких факторов среди бактерий (в результате полового размножения) очень мешают врачам. Другие плазмидные гены определяют устойчивость к дезинфицирующим средствам, способствуют таким заболеваниям, как стафилококковая импетиго; помогают молочнокислым бактериям превращать молоко в сыр; придают способность усваивать такие сложные вещества, как углеводороды, что можно использовать для борьбы с загрязнениями океана или для получения кормового белка из нефти.
1.7 Индивидуальный рост и бесполое размножение клеток
Отношение поверхность/объем у бактериальных клеток очень велико. Это способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта. В благоприятных условиях бактерии растут очень быстро. Рост прежде всего зависит от температуры и рН среды, доступности питательных веществ и концентрации ионов. Облигатным аэробам обязательно нужен еще и кислород, а облигатным анаэробам, наоборот, нужно, чтобы его совсем не было.
Достигнув определенных размеров, бактерии переходят к бесполому размножению (бинарному делению), т. е. начинают делиться с образованием двух дочерних клеток. Переход к делению диктуется отношением объема ядра к объему цитоплазмы. Перед клеточным делением происходит репликация ДНК, во время которой мезосомы удерживают геном в определенном положении (см. рис. Строение клетки). Мезосомы могут прикрепляться и к новым перегородкам между дочерними клетками и каким-то образом участвовать в синтезе веществ клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20 мин.; интервал между делениями называется временем генерации.
Образование перегородки (рис. 14) начинается после прекращения репликации ДНК, а затем до самого деления проходит время 20 мин (так называемый D-период). Нормальный период времени, необходимый для полной репликации хромосом, составляет около 40 мин. независимо от скорости роста. Эти величины определяют момент инициации репликации ДНК в клетках, растущих с любой данной скоростью. Например, если время удвоения клетки равно 30 мин., то репликация ДНК заканчивается за 20 мин. до деления, а инициация происходит за 40 мин. до этого, так что инициация репликации хромосом, разделенных между дочерними клетками, совпадает с делением, приведшим к образованию их прародителей. Очевидно, инициации должны происходить в клетках такой культуры каждые 30 мин.
Рис. 14. Схематическое изображение клеточного цикла бактерий, показывающее распределение во времени процессов инициации репликации ДНК, возникновения перегородки и деления клетки в 50-минутном цикле.
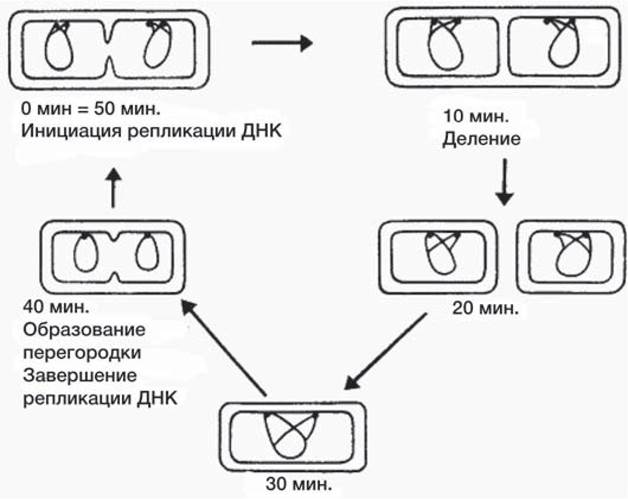
Если время генерации изменяется до 50 мин. (рис. 14), то инициация репликации ДНК для дочерних клеток происходит за 60 мин. до их разделения, т. е. за 10 мин. до деления, приводящего к образованию родительской клетки. Таким образом, мы видим, что репликация ДНК может инициироваться в любой момент клеточного цикла, но этот момент для любого заданного времени генерации будет фиксирован. Клетка в своей жизни проходит разные состояния: фазу роста и фазы подготовки к делению и деления. Клеточный цикл — переход от деления к синтезу веществ, составляющих клетку, а затем опять к делению — можно представить на схеме в виде цикла, в котором выделяют несколько фаз (рис. 15).
Рис. 15. Клеточный цикл прокариот.
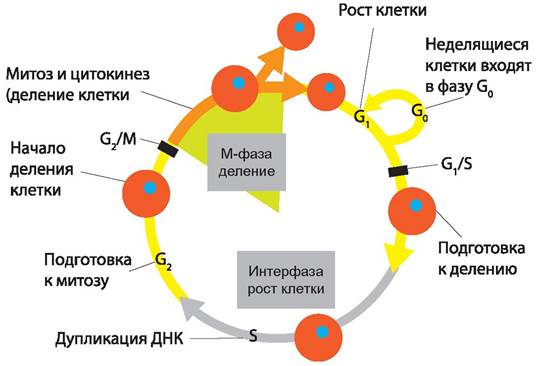
После деления клетка вступает в фазу синтеза белков и роста, эту фазу называют G1.
Для того чтобы вступить в митоз, клетка должна достичь определенной массы: размеры вновь образованных дочерних клеток вначале варьируют гораздо сильнее, чем на более поздней стадии, когда эти клетки снова вступают в митоз. Так происходит из-за того, что меньшие клетки остаются в фазе G1, пока не достигнут примерно той же величины, что и более крупные клетки. По-видимому, существует механизм регулирования массы клеток, вступающих в митоз, хотя он и работает с большими допусками.
Часть клеток из этой фазы переходит в фазу G0, эти клетки функционируют и потом погибают без деления (например, эритроциты). Но большинство клеток, накопив необходимые вещества и восстановив свой размер, а иногда и без изменения размеров после предыдущего деления, начинают подготовку к следующему делению. Эта фаза называется фаза S — фаза синтеза ДНК, затем, когда хромосомы удвоились, клетка переходит в фазу G2 — фазу подготовки к митозу. Затем происходит митоз (деление клетки), и цикл повторяется заново. Фазы G1, G2, S вместе называются интерфазой (т. е. фазой между делениями клетки).
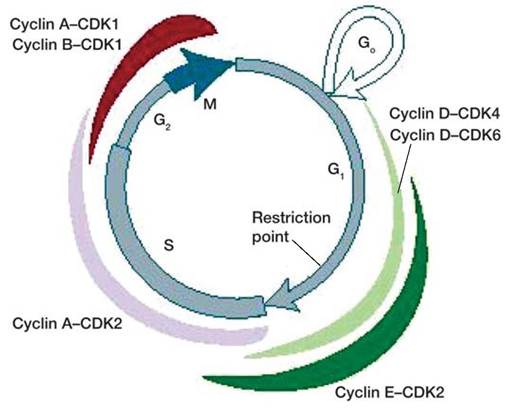
Жизнь клетки и переход от одной фазы клеточного цикла к другой регулируется изменением концентраций белков циклинов, как это показано на рисунке 16.
Рис. 16. Жизненный цикл клетки регулируется белками циклинами, концентрация которых меняется на разных фазах цикла. Толщина цветных секторов соответствует концентрации циклинов.
1.8 Половое размножение или генетическая рекомбинация
У бактерий наблюдается и половое размножение, но в самой примитивной форме. Половое размножение бактерий отличается от полового размножения эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесенная ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства, или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смешением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового размножения.
Известны три способа получения рекомбинантов. Это — в порядке их открытия — трансформация, конъюгация и трансдукция.
При трансформации клетки донора и реципиента не контактируют друг с другом. Этот процесс открыл в 1928 г. Гриффит (Griffith), работая с пневмококками — бактериями, вызывающими пневмонию. У пневмококков имеются колонии двух типов, которые различаются по внешнему виду. Одни колонии — шероховатые (R — от англ. rough — шероховатый), другие — гладкие (S — от англ. smooth — гладкий, ровный). R-штаммы не патогенны и не образуют капсулы; S-штаммы патогенны, и у них имеются толстые капсулы. Гриффит обнаружил, что если мышам ввести живые R-клетки и мертвые (убитые нагреванием) S-клетки, то мыши погибают через несколько дней, а в крови у них можно обнаружить живые S-клетки. На этом основании Гриффит сделал вывод, что из мертвых S-клеток высвобождается какой-то фактор, который придает R-клеткам способность образовывать капсулу и предохраняет их от разрушения в организме животного-хозяина. Оказалось, что такая «трансформация» наследуется. Поскольку молекулы «наследственности» в то время еще не были известны (хотя, правда, и предполагали, что это белки), очень много усилий было потрачено на то, чтобы идентифицировать трансформирующий фактор.
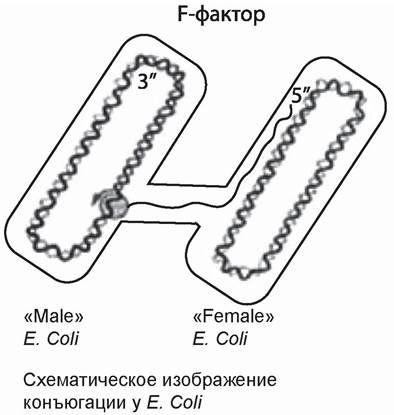
В заключение следует сказать, что половое размножение (в любой форме) — довольно редкое событие у бактерий. Но поскольку число бактерий в каждой колонии огромно, половое размножение наблюдается сравнительно часто. Такое размножение более примитивно, чем у эукариот; полный обмен геномами (суммарной ДНК) происходит только при конъюгации, что действительно встречается лишь изредка. Половое размножение бактерий имеет особое значение потому, что именно таким путем передается устойчивость к антибиотикам и дезинфицирующим средствам. В середине двадцатого века был описан половой процесс у бактерий. Это процесс, при котором бактерии обмениваются своей генетической информацией. На рисунке 17 представлена схема этого процесса. Он называется конъюгацией. Во время конъюгации образуется цитоплазматический мостик, по которому происходит перенос молекулы ДНК из одной клетки в другую. У кишечной палочки имеется молекула ДНК, которая называется F-фактор (fertility factor — фактор плодовитости). Молекула F-фактора способна встроиться в геномную ДНК. В F-факторе кодируется специальный белок, который образует половые ворсинки, они называются F-пили. Эти самые ворсинки прикрепляются к другой клетке, которые F-фактор не содержат, и F-фактор инициирует репликацию. В процессе репликации образуется две копии молекулы ДНК, причем одна копия остается в исходной клетке, а вторая копия переносится в другую клетку. То есть, генетическая информация из одной клетки попадает в другую.
Рис. 17. Половой процесс у бактерий (Бидл и Тейтум, 1946; У. Хейс, 1952)
Донорная хромосома содержит такие же гены, как и тот кусок ДНК, который был перенесен в клетку. Однако варианты генов в исходной, донорной клетке, и в клетке-реципиенте могут отличаться. Например, в исходной клетке ген кодировал синтез фермента лактазы (расщепляет молочный сахар лактозу), а в рецепиенте такой же ген испорчен, то есть лактазу не кодирует из-за какой-то мутации. При этом бактерия не способна использовать сахар лактозу в среде.
ДНК реципиента и хозяйская ДНК обмениваются гомологичными (то есть содержащими одинаковые гены) кусками. Образуется новое сочетание генов в хозяйской клетке. Среди ее старых генов оказывается встроен кусок с новым геном, прибывшим из клетки- донора. Этот процесс обмена кусками ДНК называется рекомбинацией. Та ДНК, которая в процессе рекомбинации оказалась не включенной в хромосому, деградирует и исчезает. Новый ген проявляет себя — клетка оказывается способной расщеплять тот сахар, который раньше использовать не могла. В такой ситуации ген лактазы называют генетическим «маркером», он маркирует участок хромосомы, связанный с определенным свойством бактерии (способностью расщеплять сахар, которую может детектировать исследователь). Процесс репликации у кишечной палочки продолжается 20 минут, а процесс конъюгации длится 3-5 минут. За это время успевает перейти не вся хромосома, а только ее кусочек. Чем дольше длится конъюгация, тем больший кусочек успевает перейти из одной клетки в другую. Этот процесс позволяет определить какие маркеры поступили в клетку, если исходно клетки различались по нескольким генам. F-фактор способен встраиваться в разные участки хромосомы, и когда начинается передача, разные маркеры попадают в другую клетку. После конъюгации клетки встряхивали, и мостики между ними разрывались. Это встряхивание проводили через 2, 3, 5 минут, и смотрели, какие маркеры (и, соответственно, какой фрагмент хромосомы) за это время войдут. По этим данным строили генетическую карту (расположение друг относительно друга генетических маркеров). Генетическая карта кишечной палочки была построена в 60-х годах. На этой карте были гены-маркеры, расположенные по всей кольцевой хромосоме, а координаты генов на карте обозначались в минутах. Итоговая карта, построенная в 60-х годах, имела координаты в промежутке от 0 до 90 минут. Поэтому один известный микробиолог шутил, что кишечная палочка — это удивительный организм, у которой жизнь длится 20 минут, а половой процесс — 90 минут.
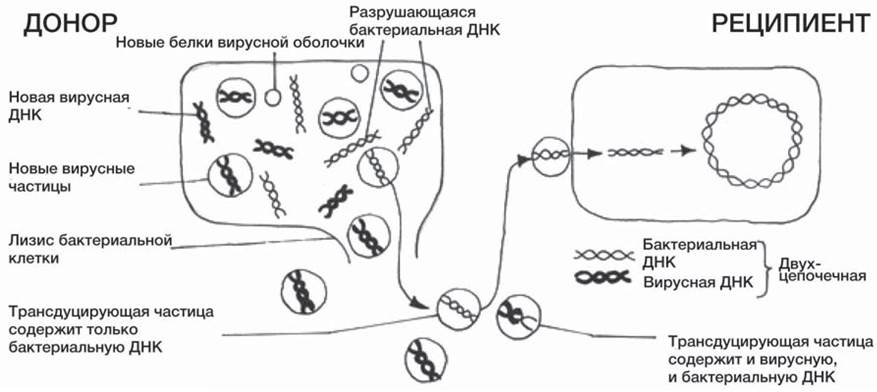
При трансдукции небольшой двухцепочечный фрагмент ДНК попадает из клетки-донора в клетку- реципиент вместе с бактериофагом (одна из групп вирусов). Возможный механизм трансдукции изображен на рисунке 18 ниже.
Некоторые вирусы способны встраивать свою ДНК в ДНК бактерий; такая встроенная ДНК реплицируется одновременно с ДНК хозяина и передается от одного поколения бактерий к другому. Время от времени такая ДНК активируется и начинает кодировать образование новых вирусов. ДНК хозяина (бактерии) разрывается, а высвобожденные фрагменты иногда захватываются внутрь новых вирусных частиц, порой даже вытесняя ДНК самого вируса. Такие новые «вирусы», или трансдуцирующие частицы, затем переносят ДНК в клетки других бактерий.
Рис. 18. Механизм трансдукции