МИКРОБИОЛОГИЯ БИОЛОГИЯ ПРОКАРИОТОВ ТОМ I - А. В. ПИНЕВИЧ - 2006
ГЛАВА 8. ЦИТОЛОГИЯ ПРОКАРИОТОВ
Ученый обязан считаться и с взглядами, противоречащими своим научным воззрениям; он должен взвешивать все мнения и выбирать только те, за которыми стоят самые веские доводы.
Л. С. Берг. Наука, ее содержание, смысл и классификация (Петроград, 1920).
Считается, что основу биологического образования, по крайней мере, в России закладывает «общая» цитология. На самом деле преподают цитологию ядерных организмов, в основном животных. Что касается строения прокариотов, то с ним мало знакомы даже специалисты-микробиологи, и современные успехи молекулярной генетики увеличили отрыв, от цитологических тылов.
Цитология прокариотов появилась после того, как к бактериальным объектам стали применять методы электронной микроскопии. На первых порах охотились за эстетичными изображениями, что при грубости методик породило множество артефактов. Следом пришла трезвая ориентация на щадящую химическую фиксацию, а затем на сверхбыструю криофиксацию. После изобретения методов молекулярного зондирования in situ в микробиологии наступил ренессанс световой микроскопии. В настоящее время получены данные о компартментализации ключевых процессов у прокариотов и их динамическом развертывании в онтогенезе.
Хотя прокариоты беднее морфологическими деталями, чем эукариоты, этот «недостаток» компенсируется большей возможностью изучать клеточное строение с помощью индуцированного мутагенеза. Кроме того, благодаря ярче выраженной способности к физиологической адаптации, ультраструктурные ответы прокариотов можно анализировать в более широком интервале внешних воздействий.
8.1. Прокариотный морфотип
Когда мы обсуждали глобальную фенотипическую систему, решающее значение имела организация мембранного аппарата (см. главу 2). Используя этот критерий, мы выяснили, что существуют только два глобальных морфотипа — эукариотный и прокариотный. Теперь же зададимся вопросом, насколько они гетерогенны.
Эукариотный морфотип един в своей основе, хотя разные филогенетические группы эукариотов различаются по ультраструктурным деталям. Напротив, прокариотный морфотип гетерогенен и подразделяется на четыре бактериальных мор- фотипа и один археотный морфотип.
8.1.1. Бактериальные морфотипы
В 1978 г. Нил Гиббонс и Роберт Мюррей предложили таксономическую систему, которая на многие годы стала венцом фенотипической классификации бактерий. Вслед за Роджером Стэниером эти авторы рассматривали понятия «бактерии» и «прокариоты» как эквивалентные, хотя в то время уже была опубликована и стала завоевывать признание концепция Карла Воза о «других» прокариотах — археях.
В основу своей системы Гиббонс и Мюррей положили тип клеточной оболочки, прежде всего — присутствие в ней ригидного слоя (или слоев) и их химический состав.
Клеточная оболочка (англ. cell envelope) — это комплекс окружающих цитоплазму мембран и немембранных структур.
Основой оболочки является клеточная мембрана, или цитоплазматическая мембрана (англ. cytoplasmic membrane, СМ). У многих бактерий помимо нее имеется наружная мембрана (англ, outer membrane, ОМ).
Наряду с мембранами в состав клеточной оболочки в большинстве случаев входит клеточная стенка (англ. cell wall, CW). Она состоит из одного или нескольких ригидных слоев (англ. rigid layer).
Архитектурный план клеточной оболочки бактерий можно дать в виде схемы (рис. 4) или записать следующим образом:
клеточная оболочка = СМ + (ОМ) + (ригидные слои).
Мембраны клеточной оболочки разграничивают три генеральных компартмента — цитоплазматический, периплазматический и экзоплазматический (рис. 4).
Рис. 4. Оболочка, ригидные слои стенки и генеральные компартменты бактериальной клетки. СМ — цитоплазматическая мембрана; ОМ — наружная мембрана.
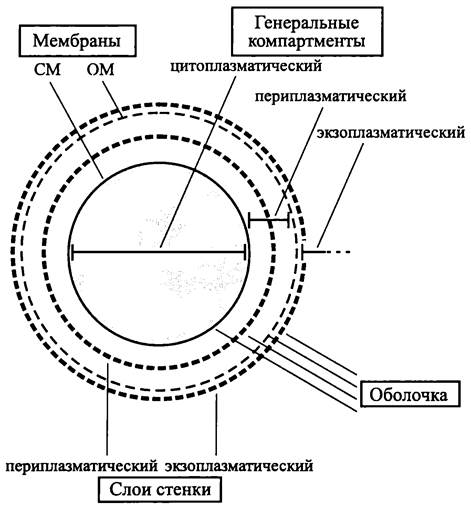
Очевидно, что при отсутствии ригидных слоев клеточная оболочка образована СМ и ОМ или только СМ. Такие клетки не имеют постоянной формы. Если они окружены одновременно СМ и ОМ, их называют сферопластами, а если они окружены только СМ — протопластами.
Ригидные слои различаются по молекулярному составу, но чаще всего их основу образует бактериальный пептидогликан, или муреин (лат. murus — стенка). Этот азотсодержащий гетерополимер подробно рассматривается в разделе 8.4.3.1. Сейчас мы только отметим, что он состоит из спирализованных гликановых цепей, которые ковалентно связаны друг с другом при помощи боковых пептидных цепочек. В результате клетка одета мономолекулярной сетью, или саккулусом (лат. sacculus — мешочек).
Система Гиббонса и Мюррея впервые позволила упорядочить бактериальные морфотипы и на этой основе создать стройную фенотипическую систему. Однако в дальнейшем она утратила таксономическое значение. Как известно, в настоящее время основой для классификации прокариотов служит их положение на родословном древе, и вместо строения клеточной оболочки ведущим таксономическим признаком является первичная структура рРНК (см. разделы 2.5 и 7.2). Тем не менее, рациональное зерно, заложенное Гиббонсом и Мюрреем в классификацию прокариотных морфотипов, сохранилось.
Поскольку этот вопрос принципиально важен для цитологии прокариотов, остановимся на нем подробно.
Основной, или грамотрицательный бактериальный морфотип (рис. 5, Б1). Гиббонс и Мюррей разделили царство прокариотов (Procaryotae) на четыре мегатаксона с разными морфотипами. Каждому из этих мегатаксонов соответствует ранг отдела:
1. Gracilicutes;
2. Fimnicutes;
3. Mollicutes (синоним — Tenericutes);
4. Mendocutes (синоним — Mendosicutes).
Рис. 5. Прокариотные морфотипы. А — археотный; Б1 —основной бактериальный (грамотрицательный); Б2 — упрощенный бактериальный (грамположительный); Б3 —усложненный бактериальный; Б4 — трихомный; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; IСМ — интрацитоплазматические мембраны; МР —микроплазмодесмы.

В отдел Gracilicutes (лат. gracilis — тонкий и cutis — кожа; «тонкокожие») вошли бактерии с однослойным муреиновым саккулусом (см. раздел 8.4.3.1). Второй особенностью этих бактерий служит наружная мембрана, или ОМ — уникальная мембранная структура, расположенная за пределами СМ.
Наружная мембрана относится к числу унитарных липопротеиновых биомембран, однако она имеет особый молекулярный состав и выполняет специфические функции. В частности, ее внешний полумембранный листок состоит из амфифильных липополисахаридов. Отсюда второе название этой мембраны — «липополисахаридная».
Термин «грациликутный» (англ. gracilicute) когда-то широко использовался в отечественной литературе, но теперь он редко употребляется. Дело в том, что корреляция между малой толщиной муреинового саккулуса и присутствием ОМ не универсальна. Как правило, толщина муреинового слоя действительно различается в клетках с ОМ или в клетках без ОМ (в первом случае <10 нм, а во втором >50 нм). Однако существуют отклонения от этой закономерности. Например, ОМ имеется у всех цианобактерий, однако у некоторых одноклеточных штаммов толщина муреинового саккулуса >10 нм, а у некоторых нитчатых штаммов она составляет 15-35 нм, а у Oscillatoria sр. даже >700 нм.
О толщине ригидного слоя можно косвенно судить по характеру окрашивания бактерий на микроскопических препаратах. Для этого используется простая методика, которую предложил в 1883 г. датский врач Ганс Грам.
Этот классический метод состоит в том, что зафиксированный материал прокрашивается анилиновым красителем кристаллическим фиолетовым, протравливается смесью Люголя (раствор J2 в водном КJ), а затем обрабатывается водным спиртом или водным ацетоном.
По современным представлениям, механизм «окрашивания по Граму» следующий. Проникшая в клетку растворимая хлорная форма красителя после протравливания переходит в йодную форму (изоморфное замещение) и выпадает в мелкозернистый осадок, который равномерно прокрашивает цитоплазму. При обработке водным спиртом или ацетоном из СМ экстрагируются липиды, в результате чего она становится пористой и не препятствует вымыванию комплекса иода с красителем. Однако помимо СМ обычно существует барьер, устойчивый к органическим растворителям — муреиновый ригидный слой. Если он состоит из многочисленных слоев с мелкими порами, то комплекс иода с красителем удерживается внутри, и клетки окрашиваются по Граму положительно. В том случае, если он образован монослоем с крупными порами, клетки окрашиваются по Граму отрицательно.
Если строго соблюдать стандартную продолжительность отдельных этапов окрашивания по Граму (нем. Gramdauer), то окажется, что бактерии по этому признаку не образуют непрерывного вариационного ряда, а подразделяются на альтернативные группы — «грамотрицательные» и «грамположительные». Объяснение этому, на первый взгляд, парадоксальному явлению заключается в том, что существуют два альтернативных варианта архитектуры бактерий:
— клетки с многослойным муреином; не имеют ОМ;
— клетки с монослойным муреином; имеют ОМ.
Среди бактерий, не имеющих ОМ, встречаются «грамвариабельные» формы (например, некоторые представители низших актиномицетов). Кроме того, характер окрашивания по Граму зависит от таких факторов, как возраст культуры, стресс, клеточная диссоциация и дифференциация. Наконец, известны примеры, когда клетки одновременно имеют ОМ и толстый муреиновый слой (см. выше).
Таким образом, тип окрашивания по Граму служит интегральной характеристикой, отражающей физические свойства ригидного слоя. Химическая природа этого слоя несущественна. В частности, грамположительные археи имеют мощный псевдомуреиновый саккулус или гетерополисахаридный ригидный слой, а грамотрицательные археи — ригидный белковый S-слой (см. раздел 8.5.3). Отрицательно окрашиваются по Граму прокариотные клетки, не имеющие ригидного слоя и окруженные только СМ — бактериальные или археотные микоплазмы (см. ниже).
Как правило, тонкий муреиновый саккулус встречается у бактерий, у которых есть ОМ. Поэтому в понятие «грамотрицательная бактерия», которое указывает на характер диагностического окрашивания, стали вкладывать архитектурный смысл. Однако если употреблять термин «грамотрицательный», не уточнив, что имеется в виду — тинкториальные (от лат. tinctura — окрашивание) свойства объекта или его морфотип, появляется путаница.
Пытаясь избежать этого, канадский бактериолог Терри Биверидж (Т. J. Beveridge) в начале 1980-х годов ввел в бактериологическую литературу понятие грамотрицательного морфотипа.
В 1994 г. автор учебника предложил называть грамотрицательный морфотип «бимембранным» (лат. bi — двойной и membrana — перепонка, «с двумя мембранами»). Радж Гупта дал в 1998 г. грамотрицательному морфотипу еще одно название — «дидермы» (греч. dis — дважды и derma — кожа; «двухкожие»). Видно, что оно заимствует у термина «грациликутный» афористичность, а у термина «бимембранный» — архитектурный смысл. Наконец, Томас Кавалье-Смит назвал бактерии, обладающие грамотрицательным морфотипом, «негибактериями» (лат. negativus — отрицательный). Поскольку дальнейшая судьба терминов «бимембранный», «дидермы» и «негибактерии» неясна, мы воспользуемся термином «грамотрицательный морфотип», несмотря на то, что он несет двойной смысл.
Для понимания природы бактериальных морфотипов важно усвоить, что подавляющая часть фил «Берги-2001» (22 из 24) — это объекты с грамотрицательным строением. Исключение составляет фила ВХIII Firmicutes, большинство представителей которой имеет грамположительное строение, а также фила BXIV Actinobacteria, состоящая только из грамположительных форм.
У бактерий, принадлежащих к грамотрицательному морфотипу, ОМ окружает всю клетку и служит внешней границей уникального периплазматического компартмента (греч. peri — около и plasma — оформленное тело; «вокруг цитоплазмы»). Внутреннюю границу этого компартмента образует СМ. Интерьер периплазматического компартмента представлен периплазматическим пространством со специфическим содержимым — периплазмой.
Периплазматический компартмент выполняет в бактериальной клетке комплекс пограничных функций. Он не только обеспечивает механическую и осмотическую защиту, но также может принимать участие в двигательных, транспортных, метаболических, сигнальных и генетических процессах.
СМ и ОМ структурно взаимодействуют, но как именно, окончательно не выяснено. Согласно первой модели, СМ и ОМ сообщаются друг с другом при помощи посредника. В этом случае от СМ отпочковываются мембранные везикулы, которые пересекают периплазматический компартмент в радиальном направлении и сливаются с ОМ, что внешне напоминает эукариотный экзоцитоз (рис. 6, A). Согласно второй модели, взаимодействие между СМ и ОМ осуществляется с помощью трубчатого анастомоза (рис. 6, Б). Однако реальнее всего третья модель (рис. 6, В). В данном случае СМ и ОМ соприкасаются в отдельных участках, образуя зоны мембранной адгезии — «контакты Байера» (см. раздел 8.4.1.4). По современным представлениям, именно там находится секреторный аппарат (см. раздел 8.6.1).
Рис. 6. Структурное взаимодействие между цитоплазматической и наружной мембраной. А — с помощью мембранных везикул; Б — с помощью анастомоза; В — с помощью зоны адгезии; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; BJ — контакт Байера.
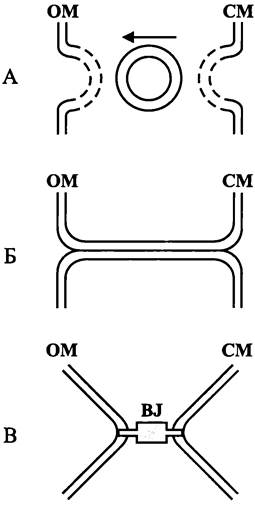
Какой бы ни была природа этого взаимодействия, участки, где оно осуществляется — это мост, по которому доставляется материал для сборки ОМ.
Упрощенный, или грамположительный, бактериальный морфотип (рис. 5, Б2). Предки некоторых бактерий в ходе морфофункциональной эволюции потеряли ОМ. Поскольку бактерии, лишенные ОМ, обычно имеют многослойный муреиновый саккулус, Гиббонс и Мюррей назвали данный тип строения «фирмикутным» (лат. firmus — прочный и cutis — кожа; «толстокожие») и отнесли такие формы к отделу Firmicutes.
Как уже указывалось, мощный ригидный слой обеспечивает положительное окрашивание по Граму. Термин «грамотрицательный», как и термин «грамположительный», первоначально указывал на характер диагностического окрашивания, и только когда Биверидж ввел термин «грамотрицательное строение», он получил дополнительный архитектурный смысл.
Автор учебника обозначил грамположительный морфотип как «унимембранный» (лат. unus — один и membrana — перепонка; «с одной мембраной»). Радж Гупта дал таким бактериям название «монодермы» (греч. monos — один и derma — кожа; «однокожие»), а Томас Кавалье-Смит предложил для них сразу два термина — «позибактерии» (лат. positivus — положительный) и «унибактерии» (лат. unus — один).
По современным представлениям, бактерии, обладающие грамотрицательным морфотипом, находятся в основании родословного древа. В свою очередь, грамположительный морфотип рассматривается как результат вторичного упрощения, которое в целом характерно для морфофункциональной эволюции прокариотов.
Усложненный бактериальный морфотип (рис. 5, Б3). Бактериальная клетка может иметь в той или иной степени выраженную эндомембранную систему — интрацитоплазматические мембраны (англ, intracytoplasmic membranes, ICM). Мы подробно рассмотрим их в разделе 8.3.5, однако уже сейчас отметим, что ICM бактерий принципиально отличаются от ICM эукариотов.
Прежде всего, ICM имеются не у всех бактерий, а только у представителей некоторых физиологических групп, причем они могут образовываться адаптивно. Един
ственным исключением служат цианобактерии — они постоянно содержат ICM, где расположен фотосинтетический аппарат.
Морфология бактериальных ICM отличается от морфологии эукариотного ваку- ома. Как правило, это монотонный комплекс, состоящий из повторяющихся структур сферической, трубчатой, пластинчатой или дискоидной формы.
Основная функция бактериальных ICM — это ассимиляция энергии, хотя известны случаи, когда они, подобно мембранам вакуома, принимают участие в конструктивном метаболизме.
Как уже отмечалось, в клетках эукариотов СМ и ICM изолированы друг от друга, и функциональное взаимодействие между ними осуществляется с помощью везикулярного транспортного аппарата. Детали этого взаимодействия хорошо изучены. Напротив, характер взаимодействия между СМ и ICM у бактерий до конца не выяснен и, возможно, он различается у разных объектов (см. раздел 8.3.5).
Посредники между СМ и ICM не обладают свойствами везикулярного транспортного аппарата, а представляют собой перемычки-анастомозы. С помощью таких мембранных каналов СМ и ICM превращаются в непрерывную структуру, или «мембранный континуум» (лат. con-tinuo — связывать друг с другом).
За исключением «тилакоидных центров» цианобактерий (см. раздел 8.3.5), молекулярный состав анастомозов неизвестен. Механизм образования новых анастомозов также не изучен — возможно, они возникают путем вторичного слияния ICM и СМ. При сильно развитой эндомембранной системе анастомозы между ICM и СМ встречаются редко, и не исключено, что это отражает дегенерацию когда-то сильно развитого континуума, который существовал у прародительских форм бактерий.
ICM характерны для фототрофных бактерий и часто встречаются у хемолитотрофных бактерий. У остальных представителей домена Bacteria ICM наблюдаются редко и незакономерно (см. табл. 7).
По предложению автора учебника усложненный бактериальный морфотип был назван «тримембранным» (лат. tres — три и membrana — перепонка; «с тремя мембранами»).
Трихомный морфотип (рис. 5, Б4. Термин «трихом» (греч. trichoma —волосы) традиционно используется в ботанике применительно к разным филаментозным структурам — нитчатым водорослям, выростам на стеблях растений, опушению на листьях и т. д. В середине 1960-х годов Старр (М. Starr) ввел этот термин в микробиологию для того, чтобы обозначить им цепочку плотно прилегающих друг к другу бактериальных клеток, с ветвлением или без ветвления. В настоящее время термин «трихом» используется бактериологами в более конкретном ультраструктурном смысле и только по отношению к бактериям, обладающим грамотрицательным морфотипом.
Трихом — это квази-многоклеточное образование, т. е. (не)разветвленный филаментозный или сфероидный агрегат, в котором клетки окружены общей ОМ и соединены трубчатыми микроплазмодесмами, пронизывающими поперечные клеточные стенки.
Трихомные формы образуют сборную группу. Они встречаются в филах ВVI Chloroflexi (например, Chloroflexus aurantiacus), ВХ Cyanobacteria (например, Prochlorothrix hollandica), ВХIIProteobacteria (например, Beggiatoa alba), BXX Bacteroidetes (например, Saprospira grandis) и BXXI Fusobacteria (например, Leptotrichia buccalis).
Благодаря микроплазмодесмам индивидуальные цитоплазматические компартменты сливаются в обобществленный цитоплазматический компартмент. Растворенные в обобществленной цитоплазме («симцитоплазме») молекулы могут диффундировать по всему трихому, что обеспечивает интеграцию метаболических и сигнальных процессов на цитоплазматическом уровне. Однако индивидуальные органеллы (хромосома, набор рибосом, ICM, цитоскелет и включения) все же остаются в пределах своих индивидуальных цитоплазматических компартментов. Благодаря тому, что трихом окружен общей ОМ, в нем также существует обобществленный пери- плазматический компартмент, содержащий «симпериплазму». Это, в свою очередь, интегрирует транспортные и сигнальные процессы на периплазматическом уровне.
Рост трихома осуществляется путем деления отдельных клеток, а размножение трихома — с помощью фрагментации, которая может быть случайной или запрограммированной. Во втором случае разлом проходит по «некридию» (греч. nekros — мертвый), или интеркалярной клетке, погибшей в результате апоптоза.
Таким образом, трихом можно рассматривать как квази-синцитий. Он образуется благодаря тому, что при делении интеркалярных клеток поперечные перегородки не расслаиваются. Кроме того, в делении не участвует ОМ.
Бактерии, обладающие трихомным морфотипом, либо содержат ICM, либо лишены ICM.
Способность образовывать трихом генетически детерминирована. Также генетически детерминирована способность образовывать мультимерные или олигомерные трихомы. По предложению автора учебника образование олигомерных трихомов было названо «бревитрихомией» (лат. brevis — короткий и греч. trichoma — волосы; «с коротким трихомом»). Число клеток в трихоме не постоянно и варьирует в норме или при патологии. В частности, старые или голодающие трихомы рассыпаются на одиночные клетки. Кроме того, существуют спонтанные или индуцированные одноклеточные мутанты трихомных бактерий с высокой частотой реверсии к дикому типу. Но одноклеточные бактерии дикого типа трихомов не образуют.
Связь между бактериальными морфотипами и присутствием ригидного слоя клеточной стенки. У большинства представителей домена Bacteria в составе оболочки имеется ригидный слой, который представлен муреиновым саккулусом (см. разделы 8.4.3.1 и 8.5.2.1). В некоторых случаях вместо него или наряду с ним эту роль выполняет экзоплазматический белковый S-слой (см. раздел 8.5.2.3).
Среди бактерий, не имеющих муреина, встречаются и формы, обладающие грамотрицательным морфотипом, и формы, обладающие грамиоложительным морфотипом. В первом случае это представители фил BXV Planctomycetes и BXVI Chamydiae; во втором случае это микоплазмы (фила ВХIII Firmicutes, класс II Mollicutes). Гиббонс и Мюррей поместили их в отдел Mollicutes (лат. mollis —мягкий и cutis — кожа; «мягкокожие»). Позже они заменили это название на Tenericutes (лат. tener — нежный и cutis — кожа; «нежнокожие»).
Наконец, у хламидий имеется ригидный слой, представленный периплазматиче- ским белковым саккулусом (см. раздел 8.4.3.2).
8.1.2. Археотный морфотип
Поскольку у архей нет муреина, Гиббонс и Мюррей в 1978 г. отнесли их к отделу Mendocutes (лат. mendosus — уродливый и cutis —кожа; «уродливокожие»).
Археи не синтезируют липополисахаридов и не имеют ОМ. Их мембранная система максимально рационализирована и состоит из одной СМ. Они не имеют ана-
лога бактериальных ICM или вакуома (рис. 5). Хотя в цитоплазме некоторых архей встечаются мембранные структуры, они представляют собой газовые везикулы, ограниченные неунитарной мембраной (см. раздел 8.3.11.2). При электронно-микроскопическом исследовании клеток метаногенных архей находили ICM, однако в 1980-е годы было доказано, что они имеют артефактную природу.
Таким образом, археи обладают таким же планом строения, что и бактерии упрощенного, или грамположительного морфотипа. Однако сходство имеет конвергентную природу, поскольку у этих прокариотов липидный матрикс СМ состоит из уникальных липидов (см. раздел 8.3.3).
Археи конвергируют с бактериями по негативному цитологическому признаку, отсутствию вакуома, и относятся к прокариотам. Вместе с тем, археи конвергируют с грамположительными бактериями по такому негативному признаку, как отсутствие наружной мембраны. По отношению к ним может быть использован термин «квази- грамположительный» мофотип.
Связь между археотным морфотипом и присутствием ригидного слоя. У большинства представителей домена Archaea имеется клеточная стенка. Ее ригидный S-слой (см. раздел 8.5.3.1) образован экзоплазматическими белковыми субъединицами.
У шаровидных представителей экстремально галофильных архей из филы АII Euryarchaeota, например, Halococcus morrhuae ригидный слой состоит из гетерополисахарида (8.5.3.3). Очень редко в роли ригидного слоя выступает аналог бактериального муреина — археотный псевдомуреин (см. раздел 8.5.3.2).
Наконец, в виде исключения среди архей встречаются лишенные клеточной стенки микоплазмы (например, Thermoplasma acidophilum).
8.1.3. Связь между прокариотным морфотипом и размером прокариотной клетки
Прокариоты имеют микроскопическую величину (греч. micros — небольшой и skopos — наблюдатель; «мелкий для смотрящего»).
В большинстве случаев линейный размер прокариотной клетки составляет 1- 3 мкм. Поэтому для ее визуального наблюдения нужен световой микроскоп — оптический прибор, увеличивающий до тысячи раз. В свою очередь, субклеточные структуры прокариотной клетки не видны в световой микроскоп, поскольку их размеры обычно лежат за нижним краем оптической шкалы. Они не отражают волны видимого света (ʎ = 350-750 нм), и для их наблюдения используется электронный микроскоп, создающий сфокусированный пучок электронов с длиной волны, на порядок меньшей.
«Затерянный» мир, в который с помощью своего уникального метода наблюдения впервые проник Левенгук (см. раздел 1.1), заселяют не просто микроскопические существа, а организмы, имеющие прокариотное строение.
Встает вопрос, что определяет нижнюю границу размера прокариотной клетки, и существует ли связь между прокариотным морфотипом и микроскопической величиной прокариотов? Иными словами, прокариоты так малы из-за своего строения? Или прокариотный морфотип является следствием их микроскопического размера?
Оказывается, размер прокариотной клетки определяется прокариотным морфотипом.
8.1.3.1. Причины микроскопического размера прокариотной клетки
В живой клетке всегда перераспределяются вещество, энергия и информация, и она обменивается ими с окружающей средой. Поэтому принципиально важна функциональная специализация органелл, а также эффективность пограничных процессов. В первом случае встает вопрос о компартментализации (см. раздел 8.2), а во втором — о соотношении между поверхностью клетки и ее объемом.
Линейный размер большинства прокариотов приближается к размеру митохондрии и на порядок меньше линейного размера ядерных клеток, что дает 1000-кратную разницу в объеме.
У относительно более крупных одноклеточных ядерных организмов (протистов) физиологические процессы были бы крайне неэффективными, если субстраты и продукты биохимических реакций диффундировали бы по всему объему цитоплазмы, а также молекула за молекулой пересекали СМ. Поэтому такие клетки разделены мембранами на высокоспециализированные отсеки-компартменты, которые имеют автогенное или симбиогенное происхождение и находятся в тесном взаимодействии друг с другом. Кроме того, ядерная клетка сообщается с окружающей средой при помощи цитоза, при котором отдельные порции жидкой, твердой или смешанных фаз в обоих направлениях пересекают СМ.
У относительно более мелких прокариотов, как правило, отсутствует мембранная компартментализация (см. раздел 8.2), и у них, за редчайшими исключениями, нет цитоза. Поэтому метаболизм прокариотной клетки зависит в основном от внутриклеточной диффузии, а также от «поштучного» транспорта молекул через СМ. Подобно грибам, прокариоты являются осмотрофными организмами — они диффузионным способом импортируют питательные субстраты и экспортируют продукты метаболизма.
При использовании такой «экстенсивной» транспортной стратегии принципиальное значение приобретает количественное отношение поверхности клетки (S) к ее объему (V), или поверхностно-объемный коэффициент. В простейшем случае, когда клетка имеет форму сферы, он равен:
S/V = (4πR2)/(4/3πR3) = 3/R.
Таким образом, с уменьшением линейного размера клетки увеличивается ее относительная поверхность, что создает предпосылки для более эффективного осмотрофного питания. Именно в этом и заключается одна из главных причин микроскопического размера прокариотов.
8.1.3.2. Рациональный и нерациональный дизайн прокариотной клетки
Как известно, среди геометрических фигур при равном объеме минимальную поверхность имеет сфера, поэтому такой дизайн прокариотной клетки нерационален. Однако это не означает, что шаровидные прокариоты, или кокки встречаются реже других. Напротив, они очень широко распространены в природе, хотя обычно имеют малые размеры. Данное ограничение не распространяется на некоторых фотоавтотрофов, в частности на шаровидных цианобактерий, а также на тех шаровидных гетеротрофных бактерий, которые находят нетривиальные решения проблемы, как обеспечить высокий поверхностно-объемный коэффициент (см. раздел 8.1.3.3).
У палочковидных прокариотов, например, Е. coli довольно рациональный дизайн, и при равной массе тела их поверхностно-объемный коэффициент в 2-3 раза выше, чем у равновеликой сферы.
Более рациональным дизайном, чем «гладкие» сферы и палочки, обладают клетки той же общей формы, но снабженные выростами (см. раздел 8.3.9.1).
Еще более рациональный дизайн имеют нитчатые бактерии, у которых поверхностно-объемный коэффициент в 5 раз выше, чем у сферы. Однако дальнейшее уменьшение диаметра с одновременным увеличением длины повышает риск механического повреждения клетки.
Самый рациональный дизайн имеют плоские и одновременно с этим очень тонкие археи «квадратной» или «треугольной» формы (см. раздел 8.3.9.1). У них поверхностно-объемный коэффициент в 5-10 раз выше, чем у сферы эквивалентной массы.
8.1.3.3. Функциональные последствия микроскопического размера прокариотной клетки
Микроскопический размер прокариотной клетки накладывает отпечаток на целый ряд аспектов ее биологии.
Плотная упаковка систем жизнеобеспечения. Из-за микроскопического размера прокариотной клетки в ней помещается мало генетического материала, даже если он упакован очень плотно. В частности, вследствие высокой компактизации кольцевая молекула ДНК с контурной длиной ~1 мм занимает объем ~1 мкм3 (см. раздел 8.3.6.1). Для сравнения отметим, что ДНК эукариотной клетки имеет контурную длину ~1 м и упаковывается в объеме ~1000 мкм3.
Из-за ограниченного размера прокариотной клетки ее цитоплазма приобретает очень густую консистенцию (содержание воды <70%) и примерно на 85% состоит из макромолекул (табл. 5). Благодаря относительно высокому содержанию осмотически активных веществ (~15%) на СМ оказывается высокое гидростатическое давление, или цитоплазматический тургор, максимальная величина которого составляет 20 бар (см. раздел 8.3.9.1).
Таблица 5. Усредненный молекулярный состав прокариотной клетки
Компонент |
% сухого веса |
ДНК |
5 |
РНК* |
15 |
Растворимые белки** |
65 |
Простые органические молекулы |
10 |
Неорганические ионы |
5 |
* рРНК - 85%; тРНК - 15%; мРНК - 5%;
** 103 типов; 2,5106 молекул на клетку.
Роль силы трения для прокариотной клетки (специфика существования при малых числах Рейнольдса). Прокариоты растут и развиваются только в жидкой гидрофильной среде, которая может находиться в статическом или динамическом состоянии (см. раздел 7.3).
При пассивном или активном взаимодействии с набегающим водным потоком (соответственно у седентарных и самостоятельно перемещающихся форм) клетка
испытывает на себе сочетанное действие сил инерции и сил трения. Какая из них имеет большее значение?
Преимущественное воздействие на объект силы инерции или силы трения оценивается в гидродинамике числом Рейнольдса (величина, названная в честь английского физика и инженера Осборна Рейнольдса; О.Reynolds, 1842-1912). В потоке жидкости, омывающем физическое тело, могут доминировать три силы — сила инерции, сила трения и сила давления, причем обычно рассматриваются только первые две из них.
Сила инерции рассчитывается по формуле:
FHH = l3pv/t = l2pv2,
где v — характерная скорость потока жидкости; I — характерный линейный размер объекта; р — плотность жидкости.
В свою очередь, сила трения рассчитывается по формуле:
FTp = μvl,
где μ — динамический коэффициент вязкости жидкости; v — характерная скорость потока жидкости; I — характерный линейный размер объекта.
Число Рейнольдса (Re) отражает соотношение массовых сил с вязкостными силами в жидкости и рассчитывается по формуле:
Re = ГНн/FТр = (I2рv2)/μvI = рvI/μ.
Для быстро плывущего человека число Рейнольдса составляет ~104, для аквариумной рыбки гуппи ~102, тогда как для бактерий и архей, движущихся с характерной скоростью ~10 мкм с-1, оно составляет ~5 • 10-5. Таким образом, в случае прокариотов числитель дроби Fhн/FТр ничтожно мал по сравнению со знаменателем. Это означает, что инерция для них несущественна, зато они очень сильно зависят от силы трения.
Прокариоты живут в мире «без инерции» — в частности, после выключения жгутикового мотора Е. coli проплывает по инерции за 5 • 10-6 с расстояние не более 0,1Ä (что составляет 1 • 10-6 длины тела этой бактерии) и останавливается как вкопанная.
В мире малых чисел Рейнольдса бессмысленно использовать для движения возвратно-поступательный механизм (это все равно, что для человека плавать в бассейне с медом со скоростью ~1 см • мин-1). Выход из этого тупика прокариоты нашли в том, что «изобрели» гребной винт, роль которого выполняет вращающийся жгутик (см. раздел 8.6.2).
За счет трения на поверхности клетки постоянно удерживается жидкая оболочка, или водная «шуба». По толщине она приближается к диаметру клетки и служит дополнительным диффузионным барьером, через который транспортируются питательные субстраты и отходы жизнедеятельности. Эта локальная среда повторяет форму клетки, и относительная величина ее поверхности служит дополнительным критерием рациональности клеточного дизайна.
Локальная среда обновляется только в ходе пассивной диффузии (для того, чтобы «проветрить» ее только на 10%, клетке пришлось бы двигаться со скоростью 700 мкм • с-1, что на порядок выше реально возможной). Таким образом, движение не способствует улучшению режима питания, поскольку прокариотная клетка постоянно увлекает за собой свою локальную среду.
Проникающая способность прокариотов. Благодаря своим микроскопическим размерам свободноживущие прокариоты способны колонизировать монолитные, на первый взгляд, субстраты. Они проникают в микротрещины и микрокапилляры, пронизывающие осадочные и изверженные породы, почвенные частицы, толщу льда, а также поверхность строительных материалов.
Одни симбиотические прокариоты вступают в эпифитные и эпизойные ассоциации, колонизируя микроскопическую текстуру поверхности макроорганизмов. Другие проникают внутрь своих хозяев, образуя эктосимбиозы и эндосимбиозы, в том числе на внутриклеточном уровне. Меньший, по сравнению с эукариотной клеткой, размер бактерий был необходимым элементом эндоцитобиотического сценария происхождения митохондрий и пластид.
Для болезнетворных форм малый размер является одним из факторов вирулентности, поскольку за счет этого они проникают в макроорганизм через микроповреждения покровов, а затем разносятся по органам и тканям.
Каталитическая активность прокариотов. Хотя прокариотная клетка ничтожно мала, популяция в целом обладает огромной суммарной площадью поверхности. Приведем «скромный» пример — при диаметре клетки 2,5 мкм общая поверхность кокков, содержащихся в одном миллилитре популяции средней плотности (106 клеток • мл-1), составляет около 1 см2.
На поверхности прокариотной клетки иммобилизованы экзоферменты, и через нее же они секретируются во внешнюю среду. Таким образом, популяция прокариотов представляет собой гигантский тонкодисперсный катализатор. Действие прокариотных экзоферментов вызывает макроскопические изменения в окружающей среде и обеспечивает глобальный круговорот биогенных элементов.
Применительно к интересам человека действие прокариотных экзоферментов может быть позитивным или негативным.
Позитивное действие выражается в том, что экзоферменты участвуют в разнообразных биотехнологических процессах (биотехнологию можно определить, как эксплуатацию ферментативной активности микроорганизмов или клеточных культур для получения целевых продуктов, производство которых химическим путем невозможно или нерентабельно).
Негативное действие выражается в том, что экзоферменты вызывают разрушение инертных материалов (пример — порча облицовки зданий и архитектурных украшений, а также коррозия металлов в результате образования серной кислоты тионовыми бактериями из рода Acidithiobacïllus) или биологических структур (примером служит коллагеназная активность представителей рода Fusobacterium, вызывающих ряд заболеваний у человека, в частности парадонтоз).
8.1.3.4. «Гигантские» прокариоты
Итак, мы установили, что микроскопический размер прокариотов напрямую связан с осмотрофным способом питания.
Если диаметр бактерий или архей составляет 1-3 мкм, «диффузионная» проблема обычно решается на морфогенетическом уровне — клетка имеет вытянутую форму или содержит отростки, так что поверхностно-объемный коэффициент оказывается более высоким, чем у равной по массе сферической клетки. Представители рода Hyphomicrobium образуют гифы длиной до нескольких десятков микрометров, диаметр которых меньше 0,2 мкм.
Обитатель ротовой полости человека Spirochaeta plicatilis достигает в длину 250 мкм, хотя диаметр клетки не больше 0,75 мкм (существуют спириллы значительно большего диаметра, например, обитатель пищеварительного тракта головастиков Sporospirillum praeclarum имеет размер 4 x 100 мкм).
В том случае, когда трихомы бактерий или цепочки архей состоят из десятков и сотен клеток, они имеют длину больше 100 мкм. Однако диаметр такой полимерной структуры, как правило, не превышает 1 мкм, причем каждая клетка решает диффузионную проблему тривиальным способом — благодаря удлиннению по продольной оси. Гораздо реже трихомы состоят из клеток диаметром до 8 мкм, примером чего служит цианобактерия «Lyngbya majuscula».
Бактерии сферической, эллипсоидной или цилиндрической формы в виде исключения могут иметь диаметр >10 мкм. В качестве примеров особо крупных бактерий можно привести серных хемолитотрофов Macromonas mobilis (диаметр до 15 мкм) и Thiovulum majus (до 25 мкм), оксигенного фототрофа Prochloron didemni (до 30 мкм), а также факультативного хемогетеротрофа Beggiatoa gigantea (до 50 мкм). В данном случае диффузионная проблема приобретает особую остроту и решается разными способами (см. ниже).
Археи по размерам несколько меньше бактерий, и среди них не встречаются крупные кокки и палочки.
В частности, сферические представители филы AI Crenarchaeota (Acidianus infemus, Ignicoccus islandicus, Metallosphaera sedula и др.) имеют диаметр меньше 3 мкм. Нитчатые представители этой филы (Thermofilum pendens, Thermoproteus tenax и др.) вытянуты в длину на 100 мкм, но их диаметр меньше 0,4 мкм.
Сферические представители филы АII Euryarchaeota (Archaeoglobus fulgidas, Ferroglobus placidas, Halococcus morrhuae, Methanococcus vannielii, Methanosphaera stadtmanae, Pyrococcus furiosas и др.) в диаметре не превышают 2 мкм. Отдельные палочки (например, Halorubrum saccharovorum) имеют диаметр до 1 мкм и длину до 12 мкм, а цепочки из палочек (например, Methanosaeta concilii) достигают максимальной длины 150 мкм при размере клетки 1 x 5 мкм.
Археи диаметром больше 5 мкм, тем более «гигантские» археи неизвестны.
Среди прокариотов особо крупный размер (диаметр >50 мкм) имеют только несколько видов «гигантских» бактерий. Они решают диффузионную проблему одним из трех нетривиальных способов.
Увеличение площади поверхности без изменения размера и формы клетки: образование «ворсинок» СМ. Абсолютным чемпионом по величине среди бактерий считается Epulopiscium fishelsoni (греч. epulo — сотрапезник). Первое сообщение о ней опубликовал австралийский протистолог Клименте (К. Clements) в 1989 г. Эта бактерия, которую одно время принимали за протиста, относится к филе ВХIII Firmicutes и принадлежит к числу некультивируемых форм (см. раздел 7.1). Нишей для нее служит пищеварительный тракт рыбы-хирурга Acanthurus nirgofuscus (сем. Acanthuridae), которая живет в Красном море, а также на Большом Барьерном рифе, расположенном в Коралловом море по краю материковой отмели вдоль северо-восточного побережья Австралии.
Веретеновидные клетки Е. fishelsoni можно разглядеть невооруженным глазом, поскольку их максимальный размер составляет 80 х 600 мкм (по массе это примерно в 106 раз больше Е. coli).
Е. fishelsoni имеет ряд нетривиальных цитологических признаков. Например, при размножении несколько дочерних клеток образуется внутри материнской, что имеет общую основу с механизмом образования эндоспор (см. III том учебника).
Для решения диффузионной проблемы Е. fishelsoni прибегают к способу, напоминающему тот, который используют млекопитающие и птицы с целью увеличения всасывающей поверхности тонкой кишки. В данном случае СМ образует ворсовидный слой, состоящий из трубчатых инвагинаций длиной до 1 мкм и диаметром около 0,2 мкм. Даже не прибегая к математическим расчетам, мы видим, что это приводит к значительному увеличению площади поверхности СМ.
Уменьшение объема цитоплазмы без изменения размера и формы клетки: образование включений. Одну из групп некультивируемых прокариотов образуют «бросающиеся в глаза» (англ, morphologically conspicuous) гигантские серные бактерии. Они живут на поверхности или в верхнем слое морских и пресноводных осадков, получая энергию путем окисления S2- или S0 с помощью О2, аниона NO3- или катиона Fe3+. Особенность экологии этих бактерий заключается в том, что они находятся в зоне хемоклина — на пересечении встречных градиентов концентрации указанных субстратов. Абиогенный сульфид, а также сульфид, образующийся в анаэробной зоне при разложении органики или при сульфатном дыхании, легко подвергается спонтанному окислению. Поэтому серные бактерии вынуждены конкурировать с химическими окислителями. В свою очередь, биологическое окисление сульфида, которое связано или не связано с получением энергии, является одним из способов адаптации серных бактерий к этому токсичному соединению.
Чтобы обеспечить динамический баланс между оптимальной концентрацией сульфида и окислителя, серные бактерии используют особые цитофизиологические стратегии.
Примером служит «гигантская» серная бактерия Achromatium oxaliferum (класс «Gammaproteobacteria»), которая повсеместно встречается в неглубоких водоемах на поверхности богатого сульфидом ила. Ее клетки имеют сферическую, эллипсоидную или цилиндрическую форму при максимальном линейном размере 125 мкм и максимальном объеме 3,5 • 104мкм3 (в 104 раз больше, чем у клетки Е. coli). В результате окисления сульфида кислородом в периплазматическом компартменте A. oxaliferum накапливаются серные глобулы (см. раздел 8.4.4.1), содержимое которых затем окисляется до сульфата. Уникальной особенностью A. oxaliferum служит то, что помимо серных глобул клетки содержат многочисленные кристаллы кальцита (СаСОз). В результате на долю цитоплазмы и органелл приходится не больше 2% внутреннего объема клетки. В данном случае проявляется полифункциональность цитологических структур у прокариотов, поскольку смысл запасания кальцита заключается еще и в нейтрализации образуемого сульфата.
Уменьшение объема цитоплазмы без изменения размера и формы клетки: образование вакуолей. Можно значительно уменьшить объем цитоплазмы, оттесняя ее на периферию крупными центральными вакуолями (см. раздел 8.3.11.1). В данном случае речь идет о «нитратных» и «кислородных» вакуолях у гигантских серных бактерий из родов Beggiatoa, Thiomargarita и Thioploca.
Здесь также проявляется полифункциональность цитологических структур, поскольку эти вакуоли играют роль анаэробных или аэробных «легких», которые запасают анионы NO3- или О2, используемые для окисления экзогенного сульфида. В настоящее время нитратные вакуоли обнаружены и у одноклеточных, и у трихомных бактерий.
«Гигантская» одноклеточная некультируемая бактерия Thiomargarita namibiensis (класс «Gammaproteobacteria») живет в поверхностном слое ила на шельфе у юго-западной оконечности Африканского континента, где из-за сильного апвеллинга, т. е. подъема воды к поверхности аноксические условия создаются уже на небольшой глубине (<300 м). Сферические клетки Т. namibiensis имеют диаметр до 750 мкм и образуют цепочки, погруженные в общий слизистый чехол. Основную
часть цитоплазматического компартмента (98%) занимает вакуоль, в которой накапливаются анионы NO3- до концентрации 0,8 М. Периферический слой цитоплазмы имеет толщину 0,5-2 мкм, а в периплазматическом компартменте откладываются светопреломляющие серные глобулы, в результате чего видная невооруженным глазом цепочка клеток похожа на жемчужное ожерелье (лат. margarita — жемчуг). Поскольку клетки Т. namibiensis лишены подвижности, эта бактерия получает анионы NO3-из морской воды только при периодическом взмучивании ила. Благодаря эффективной адаптации Т. namibiensis к естественной среде обитания урожай биомассы этой бактерии составляет ~50 г • м-2.
«Гигантские» трихомные некультивируемые бактерии рода Thioploca (класс «Gammaproteobacteria») живут в поверхностном слое ила на всем протяжении континентального шельфа вдоль восточного пробережья Южной Америки. Гиперпопуляция этих бактерий занимает площадь ~104 км2, а ее продуктивность составляет ~1 кг • м-2. Цилиндрические или бочковидные клетки Т. аrаuсае и Т. chileae имеют максимальный диаметр 50 мкм. Макроскопический трихом, состоящий из сотен клеток, достигает длины в 7 см. Несколько десятков трихомов сплетаются в жгут диаметром ~1 мм (греч. ploca — косичка), одетый общим чехлом. До 80% объема цитоплазматического компартмента занимает центральная вакуоль, в которой аккумулируются анионы NO3-до концентрации 0,8 М (что в 2 • 104 раз превышает их содержание в морской воде). Подвижные трихомы Thioploca sp. обладают положительным хемотаксисом к анионам NO3-, и для их накопления периодически совершают вылазки в морскую воду.
Свойством накапливать анионы NO3- в нитратных вакуолях обладают и «гигантские» представители серных бактерий из рода Beggiatoa (класс «Gammaproteobacteria»), клетки которых имеют максимальный диаметр 150 мкм. Некоторые виды получены в чистой культуре. В отличие от Thioploca, они образуют одиночные трихомы. Эти бактерии образуют густые маты вокруг глубоководных горячих газовых выбросов у берегов Южной Америки или вокруг «холодных» мелководных газовых выбросов в Балтийском море у берегов Дании. Для накопления нитрата они используют такую же миграционную стратегию, что и морские представители рода Thioploca.
«Гигантские» трихомные бактерии из класса «Gammaproteobacteria» живут вблизи мелких морских гидротерм у берегов Калифорнии и образуют монофилетическую группу с видами родов Beggiatoa, Thiomargarita и Thioploca. Клетки этих бактерий имеют диаметр до 120 мкм и содержат центральную вакуоль, внутри которой накапливаются не анионы NO3-, а молекулы кислорода. Внутриклеточный резерв кислорода используется для дыхания при кратковременном попадании в аноксические условия, когда поверхностные слои воды смешиваются с придонными слоями, обогащенными сульфидом.
8.1.3.5. «Карликовые» прокариоты
Наряду с «гигантами» в мире прокариотов встречаются «карлики», клетки которых имеют диаметр <0,5 мкм.
Соответствующих бактерий называют «нанобактериями» (лат. nanus — карлик), «пикобактериями» (исп. pico — тонкий, как острие) или «ультрамикробактериями» (англ. ultramicrobacteria, UMB). Они проходят через мембранные фильтры, и их еще называют «фильтрующимися» бактериями.
«Карликовые» прокариоты находятся в двойственной ситуации:
С одной стороны, при уменьшении радиуса клетки поверхностно-объемный коэффициент (S/V = 3/R) возрастает, что является универсальным способом решения диффузионной проблемы. Они также обладают повышенной проникающей способностью, что позволяет им колонизировать субстраты, недоступные для бактерий обычного размера;
С другой стороны, у этих бактерий нерациональный дизайн, поскольку сфера имеет минимальную поверхность для геометрической фигуры с заданным объемом. Кроме того, когда размер клетки предельно мал, СМ уже не может вместить минимально необходимый набор белков, а в цитоплазматическом компартменте уже нельзя упаковать полноценный геном со всем ассортиментом продуктов его экспрессии.
«Карликовые» прокариоты нашли два выхода из создавшейся ситуации. В одном случае они ведут автономное существование. При этом их геном имеет нормальный размер (>2 т. п.н.), и в метаболическом отношении они полноценны. В другом случае они являются облигатными внутриклеточными симбионтами. При этом они имеют геном небольшого размера (1 т.п. н.) и из-за различных генетических дефектов находятся в метаболической зависимости от своих хозяев.
Свободноживущие ультрамикробактерии. Большинство морских бактерий, а также бактерий, обитающих в олиготрофных пресноводных водоемах, имеет диаметр <1 мкм и объем <0,3 мкм3. Более того, среди представителей пикопланктона численно преобладают клетки диаметром <0,5 мкм и объемом <0,1 мкм3. Однако в данном случае уменьшение размера служит способом адаптации к стрессу на бедной среде. Такие бактерии ультрамикроскопические только в естественных местообитаниях, а при росте на богатых питательных средах они увеличиваются до обычного размера (диаметр >1 мкм, объем >0,3 мкм3).
Истинные свободноживущие ультрамикробактерии, сохраняющие свои размеры при культивировании в лабораторных условиях, довольно редки.
Наиболее известна среди них морская бактерия Sphingopyxis alaskensis (класс «Alphaproteobacteria»), ряд штаммов которой был выделен у побережья Аляски, а также в Северном и Японском морях. Ультрамикробактерии известны также среди представителей широко распространенного морского кластера SAR11 (класс «Alphaproteobacteria»).
Пресноводные ультрамикробактерии, относящиеся к филе BXIV Actinobacteria, обнаружены в одном из озер Австрии, где их концентрация составляет 3 • 105 клеток • мл-1. Они имеют вибриоидную форму и диаметр 0,2 мкм.
Почвенные ультрамикробактерии, в свою очередь, были обнаружены среди представителей класса «Betaproteobacteria», филы BXIV Actinobacteria, филы ВХХ Bacteroidetes и филы ВХХII Verrucomicrobia.
Непатогенные эндосимбиотические ультрамикробактерии. Ситуации, когда непатогенные ультрамикробактерии поселяются в разных компартментах эукариотной клетки, довольно редки и связаны только с ядерными структурами. В частности, виды рода Holospora (класс «Alphaproteobacteria»), клетки которых имеют размер 0,5-1 мкм, размножаются в микронуклеусе или макронуклеусе инфузории Paramecium caudatum.
Сама возможность репродукции непатогенных ультрамикробактерий в цитоплазматическом компартменте прокариотной клетки противоречит классическим представлениям о том, что прокариоты не имеют эндоцитоза (см. раздел 2.3 и главу 6).
Однако можно указать на одно исключение из этого правила. В начале 1970-х годов были опубликованы, а в 2004 г. подтверждены данные о том, что ультрамикробактерии, относящиеся к классу «Alphaproteobacteria» и имеющие размер 0,4 x 1 мкм, размножаются в митохондриальном матриксе ресничных инфузорий Halteria geleiana и Uriotrichia ovata, а также клещей Ixodes ricinus.
Следует также напомнить, что от представителей класса «Alphaproteobacteria» произошли митохондрии, и по строению они представляют собой редуцированные грамотрицательные клетки.
Патогенные ультрамикробактерии. Репродукция представителей этой сборной группы ультрамикробактерий в большей степени, чем у непатогенных симбионтов, зависит от метаболических систем хозяина.
Примером патогенных ультрамикробактерий, которые размножаются в пери- плазматическом компартменте другой бактерии, является Bdellovibrio bacteriovorus (класс «Deltaproteobacteria»). Средний размер его клеток составляет 0,3 x 1 мкм. После соприкосновения с хозяином этот активно подвижный паразит пробуравливает его ОМ. Очутившись в периплазматическом компартменте, он трансформируется в репродуктивную форму — спиралевидный «бделлопласт», который растет за счет продуктов лизиса хозяйской клетки и дробится на зрелые вибрионы, покидающие опустошенную клетку через разрывы ее оболочки.
Примером эктопатогенных ультрамикробактерий служат плеоморфные паразиты высших животных — микоплазмы (фила ВХIII Firmicutes). Они не способны образовывать ни муреин, ни другие ригидные слои клеточной стенки. Клетки этих ультрамикробактерий имеют диаметр ~0,2 мкм, и их метаболическая зависимость от хозяев является следствием уменьшения размера генома. В случае Mycoplasma genitalium он составляет только 580 т. п. н. (475 открытых рамок считывания), что уступает на порядок размеру генома Е. coli (соответственно 4639 т. п. н. и 4288 открытых рамок считывания). Микоплазмы лишены цитохромов, а также ферментов цикла Кребса и неспособны синтезировать de novo пиримидиновые и пуриновые основания.
Эндопатогенные ультрамикробактерии довольно многочисленны. Они могут размножаться в разных компартментах эукариотной клетки — ядре, фагосоме и цитоплазме.
Примером патогенных ультрамикробактерий, которые репродуцируются в фагосоме, являются хламидии (фила BXVI Chlamydiae), вызывающие у человека дис- симинативные или контагиозные заболевания, в частности пневмонию, пситтакоз и трахому. Репродуктивные клетки хламидий, или «ретикулярные тела» имеют диаметр ~0,5 мкм и накапливаются внутри фагосомы, мембрана которой модифицирована таким образом, что лизосомы с ней не сливаются. Вследствие того, что размер генома хламидий составляет ~1 млн. п. н., они обладают дефектным метаболизмом и находятся в облигатной зависимости от хозяина. В частности, они не имеют собственной системы для выработки энергии и получают от него АТФ в обмен на АДФ (с помощью АДФ/АТФ-транслоказы). Конвергентный аналог этого фермента имеется только в митохондриях.
Представителями патогенных ультрамикробактерий, которые репродуцируются в цитоплазме или ядре, служат риккетсии (класс «Alphaproteobacteria»), вызывающие у человека разные формы риккетсиозного тифа, а также риккетсиозный сифилис и сыпную лихорадку. Природным резервуаром и вектором для переноса этих патогенов от грызунов к человеку служат клещи. Риккетсии не образуют экзотоксинов — вызываемые ими патологии связаны с разрушением хозяйских клеток изнутри. Диаметр риккетсий составляет ~0,3 мкм, и они имеют небольшой геном (в случае Rickettsia rickettsii, этиологического агента сыпной лихорадки Скалистых гор, его размер составляет ~1 млн. п. н.), вследствие чего эти патогены находятся в облигатной метаболической зависимости от своего хозяина. Поскольку у них отсутствуют ферменты гликолиза и глюконеогенеза, они импортируют из цитоплазмы НАДН и УДФ-глюкозу (что неспособны делать другие прокариоты, а также эукариоты). Источником энергии для риккетсий служит окисление глутамата через цикл Кребса. В дополнение к этому они занимаются «энергетическим паразитизмом», при котором обменивают АДФ на АТФ хозяина.
Симбиотические ультрамикроархеи. Среди архей известен только один такой пример — Nanoarchaeum equitans, единственный представитель новой филы «Nanoarchaeota» (см. раздел 3.5). Размер генома этого археота составляет ~500 т. п. н., а диаметр клетки ~0,4 мкм. Он является облигатным эктосимбионтом другого археота — гипертермофила Ignicoccussp. Зависимость от старшего партнера, скорее всего, связана с потерей метаболической автономии, однако в чем конкретно заключается стратегия этой уникальной ассоциации, еще не установлено.
8.2. Компартментализация у прокариотов
Несмотря на свой микроскопический размер, прокариоты имеют сложное внутреннее строение, что связано с компартментализацией.
Мы не раз подчеркивали основное структурное различие между эукариотами и прокариотами — если первые обязательно содержат ICM, то у вторых эти мембранные структуры встречаются только у некоторых групп бактерий. Кроме того, у бактерий ICM обычно образуются в результате адаптации к «автотрофному» питанию (см. раздел 8.3.5).
То, что бактерии в большинстве случаев обходятся без ICM, а археи их не имеют вообще, дало повод считать прокариотные клетки не только не компартментализо- ванными, но и внутренне изотропными. Однако это неверно.
Во-первых, установлено, что прокариотная клетка анизотропна. Для обозначения анизотропии на молекулярном уровне предложено новое понятие — «гиперструктура».
Одним из примеров гиперструктуры является кластер рецепторных белков, участвующих в хемотаксисе, своеобразный «нос» бактериальной клетки (см. разделы 8.4.2 и 8.6.2.2). Он расположен на одном из полюсов Е. coli. Другой пример гиперструктуры — это локализованный в центральной области клетки прокариотный аналог эукариотного ядрышка, в котором происходит биогенез рибосом. Третьим примером гиперструктуры является реплисома, расположенная в экваториальной зоне и обеспечивающая репликацию хромосомы и сегрегацию дочерних хромосом (см. III том учебника). Границы гиперструктуры строго не заданы, и поэтому есть разница между этим понятием и понятием органеллы (хотя в функциональном плане гиперструктура сходна с органеллой, имеющей особое расположение).
Во-вторых, прокариотная клетка разделена на отсеки, т. е. у нее существует истинная компартментализация (англ. compartment — отсек). Однако в данном случае компартментализация достигается другими средствами, чем у эукариотов, а некоторые типы прокариотных компартментов уникальны как в структурном, так и в функциональном отношении.
Чем же обеспечивается компартментализация у прокариотов и каковы ее принципы?
Органеллы-микрокомпартменты, или просто «компартменты» подразделяются на две основные группы — генеральные и специализированные. Выполняемые ими функции также подразделяются на генеральные и специализированные.
Генеральные микрокомпартменты необходимы для жизнедеятельности клетки, поскольку на их основе осуществляются основополагающие, или генеральные функции. Генеральными функциями являются все «информационные» функции (хранение, воспроизведение и процессинг генных последовательностей), а также важнейшие функции из числа «операционных» (биогенез клеточных структур, транспорт и метаболизм).
В любой клетке существуют два генеральных микрокомпартмента, разделенные унитарной мембраной, — цитоплазматический и экзоплазматический. Бактерии, обладающие грамотрицательным морфотипом, имеют еще и третий генеральный микрокомпартмент — периплазматический, который расположен между СМ и ОМ (рис. 4).
Внутри цитоплазматического генерального микрокомпартмента находятся множественные генеральные микрокомпартменты, лишенные собственной мембранной границы. К ним относятся органеллы трансляции — рибосомы, а также близкие к ним по размерам органеллы посттранскрипционного и посттрансляционного процессинга — деградосомы, шаперонины и протеасомы (см. разделы 8.3.10.1-8.3.10.3).
Специализированные микрокомпартменты выполняют адаптивные функции, и их присутствие в клетке не служит условием сохранения жизнеспособности.
Специализированные микрокомпартменты (табл. 6) находятся внутри генеральных микрокомпартментов. Соответственно, они подразделяются на:
— цитоплазматические;
— периплазматические;
— экзоплазматические.
Иногда специализированный микрокомпартмент размещается сразу в нескольких генеральных компартментах, т. е. имеет смешанную локализацию. Одним из примеров этого служит вращающийся жгутик.
Если специализированный микрокомпартмент находится внутри клетки, его называют функциональным включением (англ. functional inclusion), или просто «включением». Определение «функциональное» никогда не используется по отношению к цитоплазматическим включениям ядерных организмов — они редки и существенной физиологической роли не играют.
Если специализированный микрокомпартмент находится на наружной поверхности клетки, его обычно назвают «придаточной структурой» (англ. appendage). Если придаточные структуры, например, чехол, окутывают всю клетку, они называются «покровом» (англ, cover). Другие придаточные структуры, например, фимбрии, занимают только часть клеточной поверхности, и тогда это одна, несколько или множество идентичных поверхностных органелл.
Граница специализированного микрокомпартмента обеспечивается унитарной мембраной или иными способами (табл. 6). В соответствии с этим специализированные микрокомпартменты подразделяются на:
— ограниченные мембраной;
— ограниченные твердой фазой
— ограниченные комбинированным способом.
Таблица 6. Специализированные микрокомпартменты у прокариот
Ограниченные |
Ограниченные твердой фазой |
Ограниченные комбинированным способом |
||
Мембрана унитарная |
Мембрана неунитарная |
Мембрана унитарная |
Мембрана неунитарная |
|
В цитоплазматическом компартменте |
||||
Анаммоксосомы; нитратные и кислородные вакуоли |
Газовые везикулы |
Полифосфатные, полигликозидные и цианофицино- вые гранулы; микрокристаллы |
Магнитосомы; ацидокальцисомы; керитомические вакуоли |
Карбоксисомы и энтеросомы; полигидро- ксиалканоатные гранулы |
В периплазматическом компартменте |
||||
- |
Серные глобулы |
Микрокристаллы |
- |
- |
В экзоплазматическом компартменте |
||||
- |
Шипы; газовые баллоны |
Чехлы и капсулы; микрокристаллы; целлюлосомы |
- |
- |
Со смешанной локализацией |
||||
- |
- |
Система секреции III типа; вращающиеся жгутики; фимбрии IV типа |
- |
- |
Примечание: (-) —данный тип компартмента не выявлен.
Ограниченные мембраной специализированные микрокомпартменты окружены унитарной или неунитарной мембраной, а их содержимое находится в жидком или газообразном состоянии.
В свою очередь, ограниченные твердой фазой специализированные микрокомпартменты представляют собой микроскопические скопления твердого материала, ультрамикроскопические частицы или надмолекулярные комплексы. Границей такого микрокомпартмента служит поверхность образующей его фазы.
Что касается специализированных микрокомпартментов, ограниченных комбинированным способом, то они имеют твердофазное содержимое и окружены унитарной или неунитарной мембраной.
При классификации специализированных микрокомпартментов необходимо учитывать не только локализацию и тип строения, но и функцию. По характеру функций специализированные микрокомпартменты подразделяются на адгезионные, балластные, барьерные, запасающие, локомоторные, экспортные и энзиматические. Существуют также специализированные микрокомпартменты, которые совмещают сразу несколько из перечисленных функций.
Конкретные примеры специализированных микрокомпартментов мы рассмотрим в разделах, которые посвящены соответствующим генеральным компартмен- там.
8.3. Цитоплазматический компартмент
Границу цитоплазматического компартмента прокариотной клетки образует СМ. Каковы общие принципы организации этого компартмента и какую роль в нем играют мембранные структуры, в частности ICM?
Для ответа на поставленные вопросы необходимо вернуться к определению клетки (см. раздел 5.1).
Автором образного термина «клетка» был английский натуралист Роберт Гук (R. Нооке), который впервые употребил его в своем сочинении «Micrographia or Some Physiological Descriptions of Minute Bodies Made by Magnifying Glasses... », изданном в Лондоне в 1667 г.
Гук был родоначальником цитологии — он не только открыл клетку, но и указал на ее важнейшее свойство микрокомпартмента (а не просто обнаружил многокамерное строение некротизированной покровной ткани древесных растений, или пробки, как обычно утверждают историографы биологии).
Микроскопические полости на срезе пробки по виду напомнили Гуку ячейки пчелиных сот (в оригинале лат. cellula — ячейка). При этом он отметил, что камеры пробки имеют тонкие стенки (в оригинале лат. interstitium — промежуток) и что в сухой древесине они заполнены воздухом, а в зеленой клетке растительным соком.
Выбранное Гуком многозначное слово cellula (уменьш. лат. celia — помещение) вбирает в себя много нюансов — ячейка, каморка, келейка, карцер, клетка для животного, кладовка, кабинка и т. д. К словам celia или cellula восходят термины, обозначающие биологическую клетку в большинстве романских языков (англ. cell, исп. céllula, итал. céllula, нем. Zelle, франц. cellule и т. д.). Особо важно то обстоятельство, что Гук интуитивно уловил суть клетки как микрокомпартмента.
Роль компартментализации отражена в образном названии науки о клетке — цитологии (греч. kitos — сосуд, урна).
В понятие «клетка» постепенно стали вкладывать расширенное содержание, называя так не только периметр биологического микрокомпартмента вместе с цитоплазмой, но и совокупность внутренних деталей этого микрокомпартмента и ассоциированных с ним наружных структур. Хотя во многих случаях клетки имеют ригидные стенки, которые могут сохраняться и в постмортальном состоянии, незаменимую интегрирующую роль играет СМ.
Мембранные структуры выполняют разнообразные функции. Они отвечают за функционирование генофора и аппарата трансляции, а также за собственное сохранение и воспроизведение. Генетические, метаболические, транспортные, регуляторные и коммуникативные процессы в той или иной степени связаны с мембранами. Поэтому в клеточной биологии мембранологии всегда отводится центральное место.
Мембранологический подход использован при классификации глобальных морфотипов (см. разделы 8.1.1 и 8.1.2). Вкратце напомним, что все представители домена Archaeaобладают «минимальной» мембранной системой, которая состоит только из СМ. В то же время большинство представителей домена Bacteria имеют две мембранные структуры (СМ и ОМ), а некоторые — три мембранные структуры (СМ, ОМ и ICM).
8.3.1. Биомембраны
Мембранология как раздел клеточной биологии возникла в начале 1960-х годов. Поскольку методы ультраструктурного анализа вначале были малоинформативными, первые представления об архитектуре мембран сложились на основе анализа их биохимического состава.
Термином «мембрана» (лат. membrana — перепонка) пользовались еще классики биологии. В частности, швейцарский натуралист Альбрехт фон Галлер (А. von Haller) в своем учебнике по физиологии человека «Primae lineae Physiologiae usum Praelectiones Academicum aucrae et emenlate», изданном в 1747 г., называл мембранами соединительнотканные оболочки внутренних органов (фасции). В цитологическом смысле термин «мембрана» стали употреблять в конце XIX в., когда обратили внимание на явления, происходящие на внешней границе клетки.
Общее представление о мембранах унаследовано цитологией от органической и коллоидной химии. К началу XX в. выяснилось, что, если жирные кислоты, жирные спирты или липиды нанести на водную поверхность, они растекаются в монослойные пленки, а при эмульгировании в воде образуют мицеллы и двухслойные везикулы.
Молекулы липидов — в случае прокариотов это в основном фосфолипиды — ориентируются таким образом, что их полярный конец, или гидрофильная «головка» взаимодействует с гидрофильной фазой (в природных условиях это обычно водный раствор), а неполярный конец, или гидрофобный «хвост» разворачивается в противоположную сторону.
Билипидная мембрана — это ультратонкая и изнутри гидрофобная пленка, разделяющая два гидрофильных компартмента. Способность липидов к самосборке и упорядоченной ориентации приводит к образованию жесткой и одновременно с этим эластичной квази-кристаллической структуры. Мембраны при ничтожно малой (по сравнению с протяженностью) толщине механически прочны, поскольку их матрикс образован двумя слоями сильно взаимодействующих гидрофобных молекул. Неслучайно для солюбилизации мембранных структур используются поверхностноактивные вещества (детергенты).
При выборе физико-химического прототипа клеточной мембраны, или биомембраны учитывалось главное условие — окружающая цитоплазму пограничная пленка должна быть не растворимой в воде и физически прочной. Применительно к липидным мембранам максимально устойчивому состоянию, или минимальному уровню свободной энергии отвечает двухслойная мембрана (англ. bilayer membrane).
Первую модель строения мембраны, или «химическую» билипидную модель предложили в 1925 г. Гортер (Е. Gorter) и Грендель (F. Grendel). Согласно ей, мембрана состоит из двух зеркально ориентированных липидных монослоев. Гидрофобные взаимодействия между углеводородными «хвостами» осуществляются как в плоскостях полу мембранных листков, так и между полумембранными листками, а взаимодействие заряженных головок с молекулами воды или другого гидрофильного растворителя происходит на противоположных поверхностях мембраны (рис. 7, А).
Рис. 7. Модели билипидной мембраны. А — «химическая» мембрана (по Гортеру и Гренделю); Б — «трехслойная» биомембрана (по Дэниэлли и Дэвсону); В — «элементарная» биомембрана (по Робертсону); Г — «жидкостно-мозаичная» биомембрана (по Сингеру и Николсону); 1 — интегральный белок; 2 — периферический белок; 3 — амфипатический белок; in — внутренняя поверхность; out — внешняя поверхность.

В середине 1930-х годов, благодаря прогрессу аналитической и препаративной биохимии, модель биомембраны Гортера-Гренделя уступила место модели «трехслойной» биомембраны (англ, triple layer membrane), которую предложили Дэниэлли (J.Danielli) и Дэвсон (Н. Davson). В соответствии с ней, по обеим сторонам двойного липидного слоя лежат белковые слои. Внутренняя область, или матрикс такой мембраны проявляет амфипатические свойства (греч. amphi — двойной и pathos — страсть; «сочетание внутренней гидрофобности с внешней гидрофильностью»). Модель Дэниэлли-Дэвсона (рис. 7, Б) объясняла не только механические и электроизолирующие свойства биомембран, но и их физиологические свойства, в том числе ферментативную активность.
Следует напомнить, что из-за своей ультрамикроскопической толщины биомембраны были в то время недоступны для наблюдения in situ.
В конце 1950-х годов, когда в цитологии стали использоваться электронно-микроскопические методы, выяснилось, что животные и растительные клетки окружены мембраной, которая имеет толщину ~8 нм и состоит из примерно одинакового количества липидов и белков. Одновременно с доказательством мембранной природы по-
граничного слоя были получены данные, что ядерная оболочка, аппарат Гольджи и вакуоли, в свою очередь, представляют собой мембранные компартменты и что клетка пронизана сетью мембран — эндоплазматическим ретикулумом (лат. reticulum — сеточка; англ. endoplasmic reticulum, ER). Общий вывод был таков, что мембранные структуры универсальны для клеток животных и растений, имеют стандартную толщину и характерный трехконтурный профиль на срезе (два электроноплотных слоя, разделенные электронопрозрачным слоем).
Все это позволило Робертсону (D. Robertson) распространить физико-химическую модель трехслойной мембраны на живые объекты и предложить модель «элементарной» биомембраны (англ. elementary membrane). Согласно этой модели, биомембрана состоит из липидного матрикса с асимметричными антиподными поверхностными слоями. Внутренний слой представлен негликозилированным белком, а наружный слой — гликопротеином (рис. 7, В). Модель «элементарной» биомембраны стала важным шагом к объяснению морфофункциональных свойств мембранных структур у животных и растений.
Позднее модель «элементарной» мембраны распространили на клетки других ядерных организмов — грибов и протистов, а также на прокариотов-бактерий. Общими усилиями биохимиков и цитологов была создана концепция универсально распространенной, или «унитарной» биомембраны (англ. unit membrane).
Вернуться к модели биомембраны Дэниэлли-Дэвсона, но уже на новом качественном уровне и максимально близко к реальным физиологическим условиям позволила «жидкостно-мозаичная» модель биомембраны (англ. liquid mosaic membrane) (рис. 7, Г). Ее разработали в начале 1970-х годов Сингер (S. Singer) и Николсон (G. Nicholson) на основе следующих положений:
— билипидный слой способен к обратимым фазовым переходам из кристаллического состояния в жидкое, причем во втором случае белки и надмолекулярные комплексы обладают горизонтальной плавучестью, т. е. они диффундируют в липидной фазе с образованием квази-стационарных физиологических ансамблей;
— мембранные белки обладают вертикальной плавучестью, т. е. они интегрированы в мембрану в меру своей гидрофобности, а не в соответствии со своим размером;
— физиологическая роль белка зависит не только от его принадлежности к определенному функциональному классу, но и от ориентации по отношению к мембране.
Таким образом, под унитарными жидкостно-мозаичными мембранами подразумеваются структуры, состоящие из двух зеркально ориентированных липидных монослоев, в которые интегрированы или с поверхностью которых связаны полипептиды. Общее расположение полипептида и ориентация его доменов по отношению к липидному матриксу определяются профилем гидропатичности (распределением гидрофильных и гидрофобных радикалов вдоль цепи аминокислотных остатков). В соответствии с этим мембранные белки подразделяются на три группы — интегральные, периферические и амфипатические (рис. 7, Г 1-3).
Авторы модели унитарной биомембраны понимали, что архитектура мембран зависит от характера взаимодействия липидов с полипептидами, а также с полисахаридами. Эти компоненты мембраны не только не уменьшают, но, напротив, повышают прочность липидного матрикса. В то же время они делают строение мембран менее регулярным, а их состав — более лабильным как в онтогенетическом, так и в филогенетическом плане. Все это дает основание рассматривать мембраны как квази-упорядоченные надмолекулярные структуры.
Особая заслуга в разработке функциональной модели биомембраны принадлежит Питеру Митчелу (Р. Mitchell, Нобелевская премия по химии, 1978 г.). В начале 1970-х годов он добавил к двум основным свойствам протеолипидной мембраны, амфипатичности и механической прочности, третье свойство — активного полупроницаемого барьера, разделяющего соседние компартменты. При затрате энергии между этими компартментами перераспределяются молекулы и ионы, благодаря чему они находятся в осмотическом неравновесии.
Мембранные полипептиды выполняют архитектурные, транспортные, метаболические, генетические и регуляторные функции. От особенностей полипептидного состава мембран зависит их физиологическая специализация. В то же время мембраны имеют универсальное свойство, связанное с гидрофобной природой липидного матрикса. Оно выражается в низкой проницаемости для гидрофильных молекул, прежде всего гидрофильных ионов. Поэтому все унитарные мембраны потенциально энергогенны — они могут запасать энергию в форме трансмембранной разности электрохимических потенциалов катионов Н+ или Na+. Естественно, что диэлектрические свойства унитарных мембран служат лишь необходимым условием ассимиляции энергии. Условием достаточным является то, что в мембране должен содержаться белковый трансформатор энергии, а из окружающей среды должен поступать энергоноситель в биологически доступной форме (в виде квантов электромагнитного поля или в виде окисляемого субстрата).
Помимо фосфолипидов, к широко распространенным, но не универсальным липидам прокариотной клетки относятся моно- и полициклические производные бесцветных полиизопреноидов (хиноны, стероиды и гопаноиды), а также окрашенные полиизопреноиды (каротиноиды). Последние существуют как в свободной форме, так и в составе пигмент-белковых комплексов, которые могут быть бинарными (каротинопротеины) или тройственными (хлорофилл-каротин-протеины).
Мембрана рассматривается в первую очередь как разделительный барьер и площадка для иммобилизации ферментов, а также транспортных, регуляторных и других белков. Билипидный матрикс гораздо реже рассматривается как компартмент, поскольку поперечник мембраны ничтожно мал по сравнению с ее протяженностью. И тем не менее это компартмент — внутри мембраны в латеральном направлении мигрируют белки и белковые комплексы; в свою очередь, молекулы липида могут перемещаться в продольном (латеральном) или в поперечном (трансверсальном) направлениях. Во втором случае говорят о том, что они «кувыркаются», или делают сальто-мортале (англ. flip-flap). В компартменте билипидного матрикса и на его поверхностях сосредоточены субстраты ферментов, а также молекулы, играющие роль сигнализаторов, регуляторов или биологически инертные.
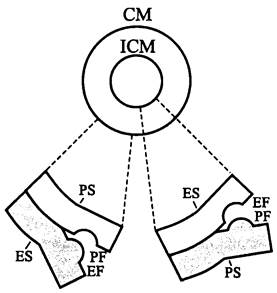
Для физиологической роли, которую выполняют мембраны in situ, важное значение имеет как латеральная, так и трансверсальная анизотропия. Методы для анализа локализации мембранных липидов еще мало разработаны, и немногочисленные данные относятся только к ОМ бактерий.
Напротив, уже давно научились выявлять мультиполипептидные комплексы и крупные белки на поверхности реплик, полученных с помощью вакуумного напыления углерода или тяжелого металла на «криосколы» — поверхности разлома замороженных клеток и субклеточных препаратов. Они имеют форму глобулярных выступов или комплементарных им углублений диаметром 5-15 нм. Большинство унитарных мембран при криоскалывании образуют два полумембранных листка, соответствующих двум липидным монослоям. По терминологии, предложенной в конце 1960-х годов Брайтоном (D. Branton), полумембранный листок, ориентированный в сторону цитоплазмы, называется протоплазматическим (англ. protoplasmic, Р). Реплика внутреннего разлома для протоплазматического листка обозначается как протоплазматическая лицевая сторона (англ. protoplasmic face, PF), а реплика внутреннего скола — как протоплазматическая поверхность (англ, protoplasmic surface, PS). Аналогичные реплики для обращенного в сторону экзоплазмы, или экзоплазматического (англ. exoplasmic, Е) полумембранного листка соответственно обозначаются как EF и ES (рис. 8).
Рис. 8. Профили криосколов унитарной мембраны. СМ — цитоплазматическая мембрана; 1СМ — интрацитоплазматические мембраны; РS — протоплазматическая поверхность; ЕS— экзоплазматическая поверхность, РF — протоплазматическая лицевая сторона; ЕF — экзоплазматическая лицевая сторона.
8.3.2. Разнообразие мембранных структур у прокариотов
Морфобиохимические и молекулярно-биологические данные, полученные при изучении бактерий и архей, привели к открытию ранее неизвестных типов биомембран. Некоторые из них имеют уникальный состав и выполняют специфические функции. Однако можно назвать ряд общих свойств, которыми должна обладать любая биомембрана. Независимо от своего конкретного строения она:
— имеет гидрофобный матрикс, состоящий из монослоя или двойного слоя молекул;
— гидрофильна по крайней мере с одной поверхности;
— выполняет функцию барьера между цитоплазмой и экзоплазмой или между периплазмой и экзоплазмой;
— образует замкнутый компартмент.
Перечисленными свойствами может обладать не только билипидный слой, но и монослой белка или липида. В первом случае речь идет об унитарной мембране, а во втором — об «неунитарной» мембране (англ. non-unit membrane). Различие между ними, в частности, морфологическое.
Срез унитарной мембраны, контрастированный солями тяжелых металлов, при просвечивающей электронной микроскопии выглядит как два темных контура, раз-
деленных светлым промежутком около 8 нм, что равно сумме длин двух липидных «хвостов». В свою очередь, поперечный срез неунитарных мембран одноконтурный и по ширине составляет 3-5 нм. На криосколах поверхностей неунитарных мембран не выявляются глобулярные частицы диаметром 5-15 нм, которые соответствовали бы интегральным и амфипатическим белкам, характерным для унитарных мембран.
Между унитарными и неунитарными мембранами существуют и функциональные различия — неунитарные мембраны не содержат транспортных систем, в том числе для создания протонного градиента. Это ограничивает их участие в физиологических процессах и сводит его к роли оболочки специализированных компартментов, а также докинг-площадки для отдельных ферментов конструктивного метаболизма. В отличие от универсально распространенных унитарных мембран, неунитарные мембраны встречаются только у бактерий и архей.
Чтобы выполнять свои функции, мембраны бактерий и архей не обязательно должны иметь протеолипидное строение и состоять из двух полумембранных листков (см. табл. 7).
Таблица 7. Мембранные структуры прокариотов
Мембранная структура |
Объект |
Функция |
Примечание |
Образованы унитарной мембраной |
|||
СМ |
Все бактерии и археи |
Полифункциональна |
У представителей филы AI Crenarchaeota липиды образуют монослой (гидрофобные «хвосты» соединены ковалентно) |
ОМ |
Бактерии грамотрицательного морфотипа |
Вместе с ригидным слоем стенки поддерживает структурную целостность клетки; участвует в транспорте и сигналинге |
У трихомных форм поддерживает структуру трихома; создает общий периплазматический компартмент |
ICM |
Бактерии грамотри- цательногоморфотипа с автотрофным метаболизмом |
Преимущественно энергоассимилирующая структура |
У представителей фил ВХ Cyanobacteria , ВХIIProteobacteria и BXV Planctomycetes могут быть изолированными от СМ |
Образованы неунитарной белковой мембраной |
|||
Оболочка интрацитоплазматических газовых везикул |
Незакономерно представлена у бактерий и архей |
Увеличивает плавучесть клетки |
Образуется адаптивно путем самосборки |
Оболочка интрацито-плазматических гранул полигидроксиалканоатов; оболочка карбоксисом |
Незакономерно представлена у бактерий |
Участвует в запасании и использовании запасного материала; участвует в иммобилизации RuBisСО и концентрировании СО2 |
Образуется адаптивно путем самосборки; содержит интегральные полипептиды, а также амфипатические и периферические ферменты |
Оболочка экстрацеллюлярных газовых баллонов |
Некоторые бактерии грамотрицате- льного (Alcaligenes sp.) и грамположительного (Bacillusvesiculiferous) морфотипа |
- |
Образуется адаптивно путем самосборки, участвует в накоплении кислорода |
Поверхностный Э-слой |
Незакономерно представлен у бактерий и архей |
Наряду с ригидным слоем (или вместо него) поддерживает целостность клетки; участвует в транспорте и сигналинге |
Участвует в образовании квази-периплазматического компартмента |
Образованы неунитарной липидной мембраной |
|||
Оболочка хлоросом |
Фототрофные представители филы BVIChloroflexi] все представители филы BXI Chlorobi |
Участвует в упаковке и биогенезе периферической светособирающей антенны |
Образуется адаптивно путем самосборки или конститутивно; содержит интегральные полипептиды |
Примечание: (-) — данные отсутствуют.
Установлено, что мембраны прокариотов более разнообразны, чем мембраны эукариотов, причем это касается не только молекулярного состава, но и функций этих структур, а также их локализации.
К сказанному можно добавить, что термин «мембрана» используется для обозначения прямых или спирализованных белковых ламелл, которые встречаются в цитоплазме некоторых бактерий и имеют толщину несколько десятков нанометров. Примером служат «R-тела» (сокр. англ. refractive — светопреломляющий) хемогетеротрофной бактерии Holospora sp. — симбионта парамеций, а также свободноживущих фототрофных бактерий, в частности пурпурной бактерии Rhodospirillum centenum. В некоторых случаях показано, что они кодируются внехромосомными генетическими элементами — плазмидами. Структура R-тел и их функциональное значение задают много загадок. Установлено, что они играют роль киллер-фактора в смешанной популяции протистов, состоящей из особей-носителей бактериальных симбионтов и незараженных особей. Возможно, что образование R-тел связано с патологическими состояниями (например, вирусной инфекцией или прионизацией); не исключено также, что некоторые R-тела имеют артефактную природу. Несмотря на внешнее сходство таких ламелл с мембранами, они к ним не относятся. Топологическое свойство истинных мембран — это везикулизация, т. е. образование замкнутых сфер и гомеоморфных фигур.
Функциональная активность мембран определяется спецификой их молекулярных компонентов, причем для белков, помимо молекулярного состава, принципиальное значение имеет распределение в трансверсальном и латеральном направлениях, а также их взаимодействие друг с другом и небелковыми молекулами.
Состав (но не физическое состояние) липидного матрикса влияет на свойства мембран в меньшей степени. Так, мембраны бактерий и эукариотов содержат структурно сходные липиды, хотя в функциональном плане сильно различаются. Напротив, мембраны бактерий и архей имеют альтернативный липидный состав, однако спектр физиологических функций у них одинаков.
8.3.3. Мембранные липиды
Липидами в широком смысле слова (греч. lipos — жир и eidos вид; «нечто жироподобное») считаются любые гидрофобные органические соединения. При расширенном толковании этого термина липидами называют даже некоторые белки, например, интегральная b-субъединица F0F1-АТФазы известна как «протеолипид». Однако собственно липиды —это углеводороды, а также азотсодержащие, кислород-, серу- и фосфорсодержащие производные углеводородов.
Липиды можно условно разделить на структурные, функциональные и запасные.
Структурные липиды. Они входят в состав биомембран, реже — в состав покровов, например, образуют ламинарный слой чехла гетероцисты цианобактерий или чехол «везикул» актиномицетов (см. раздел 8.5.4).
Функциональные липиды. Они входят в состав биомембран, где участвуют в преобразовании световой или химической энергии (хиноны и каротиноиды). Кроме того, они универсально предохраняют клетку от окислительного стресса (каротиноиды). В большинстве случаев такие липиды образуют унитарные мембраны, хотя бывают исключения — например, у зеленых фототрофных бактерий мембрана хлоросомы состоит из монослоя галактолипида.
Запасные липиды. У прокариотов они накапливаются в цитоплазме. Примером служат полигидроксиалканоатные гранулы бактерий (см. раздел 8.3.11.7). В свою очередь, ядерные клетки откладывают триацилглицериды в цитоплазме или в строме элайопластов — запасающих органелл, которые представляют собой один из вариантов дифференцированных пластид.
В настоящем разделе мы рассмотрим только структурные липиды, входящие в состав бактериальных СМ и 1СМ, а также структурные липиды археотной СМ.
На структурных липидах из ОМ грамотрицательных бактерий, и на структурных липидах квази-ОМ бактерий из группы «mycolata», мы остановимся в разделах 8.4.1.1 и 8.5.2.2.
8.3.3.1. Мембранные липиды бактерий
По химическому строению мембранные липиды прокариотов подразделяются на три основные группы:
— ацилсодержащие и/или алкилсодержащие глицеролипиды;
— полиизопреноиды;
— полиизопреноид-содержащие глицеролипиды.
Бактерии могут образовывать липиды первой и второй групп. Образование липидов третьей группы является уникальным свойством архей.
Ацилсодержащие и/или алкилсодержащие глицеролипиды имеют 1-3 гидрофобных «хвоста», которые чаще всего связаны с полиолом — глицеролом. Вместо глицерола в качестве несущего полиола могут использоваться глюкоза, дисахарид трегалоза или остатки арабинозы (в миколовых кислотах, см. раздел 8.5.2.2), а также сдвоенные остатки глюкозамина (в липиде А; см. раздел 8.4.1.1). В этом случае суммарное число гидрофобных хвостов может быть очень большим — до 50, и такая биохимическая стратегия не только увеличивает гидрофобность липидных молекул, но и дает бактериям возможность широко варьировать их структуру. Кроме того, к глицеролу обычно присоединяется полярная «головка», что позволяет глицеридам образовывать билипидный слой.
В составе молекулы полиизопреноидных липидов (гопаноидов, каротиноидов, стероидов и хинонов) нет полиола. Наличие у многих из них полярной «головки» не делает их основой биипидного слоя, хотя они корректируют его структуру (стероиды и гопаноиды) или выступают в роли функциональных липидов (каротиноиды и хиноны).
В полиизопреноидсодержащих липидах архей гидрофобные «хвосты» связаны с полиолом — глицеролом или кальдитолом (см. ниже). По аналогии с ацилсодержащими и/или алкилсодержащими липидами бактерий и эукариотов, они обычно содержат полярные головки, что позволяет им образовывать гидрофобное ядро СМ. Строение полиизопреноидной цепи в таких липидах исключительно вариабельно.
Мембранные липиды бактерий изучены в основном на примере Е. coli и В. subtilis. Собраны фундаментальные сведения о фракционном составе и биосинтезе мембранных липидов. Однако до сих пор неизвестно, существует ли латеральная и трансверсальная анизотропия липидного матрикса, т. е. насколько однородно они распределены в плоскостях и поперек мембраны.
Большинство бактериальных глицеролипидов представляет собой двойные сложные эфиры глицерола и жирных кислот — 1,2-диацил-sn-глицериды (рис.9, Б).
Напомним, что согласно стереоспецифической нумерации (англ, stereospecific numeration, sn) в проекции Эмиля Фишера (E.Fischer) верхний атом углерода идет под №1, а вторичная гидроксильная группа направлена влево от вертикально ориентированной оси молекулы. Из-за присутствия в глицероле асимметричного центрального атома углерода большинство бактериальных глицеролипидов являются левовращающими D-изомерами.
Жирные кислоты. В мембранах бактерий свободные жирные кислоты встречаются редко и в малом количестве. Обычно они образуют сложные эфиры с глицеролом. Исключением служит сложный эфир ω-алициклической С20-жирной кислоты с метанолом, относящийся к недавно открытому классу липидов — ледеранам (см. раздел 8.3.11.1).
Цепи жирных кислот в мембранных липидах различаются по длине. Они бывают разветвленными, а также могут содержать кольца и двойные связи (рис. 9, А).
Рис. 9. Мембранные липиды бактерий.
РА — фосфатидная кислота; R — остаток жирной кислоты; РG — фосфатидилглицерол; DРDG — дифосфатидилдиглицерол (кардиолипин); РЕ — фосфатидилэтаноламин и РS — фосфатидилсерин (кефалины); РС — фосфатидилхолин (лецитин); РI — фосфатидилинозитол; X — полярная группа
(этаноламин и др.; см. «Б»).
Жирные кислоты в мембранных липидах относятся к двум основным группам.
В первую, широко распространенную группу входят:
— неразветвленные насыщенные жирные кислоты (рис. 9, А, а): додекановая (C12:0; лауриновая), тетрадекановая (С14:0; миристиновая), гексадекановая (С16:0; пальмитиновая) и октадекановая (C18:0; стеариновая);
— моно-cis-ненасыщенные жирные кислоты (рис. 9, А, б): гексадеканоевая (C16:1, ∆9; пальмитолеиновая) и октадеканоевая (C18:1, ∆11; цис-вакценовая);
— жирные кислоты с циклопропановыми кольцами (рис. 9, А, е): 9,10-cis-метилен-гексадекановая и 11,12-cis-метилен-нонадекановая (лактобацилловая);
— жирные кислоты с одним метильным или одним гидроксильным заместителем (10-метилгексадеканоевая и 2- или 3-гидроксигексадеканоевая).
Все они синтезируются из ацетил-СоА в качестве праймера и малонил-СоА в качестве удлинителя цепи. Примером служит пальмитиновая кислота:
СН3СО ~ SCoA + 7-ООС - СН2 — СО ~ SCoA + 14НАДФН —> СН3 - (CH2)14 - СООН + 7СO2 + 14НАДФ + 6Н2O + 8CoASH.
Во вторую, редко встречающуюся группу входят:
— терминально разветвленные жирные кислоты в изо-конфигурации (рис. 9, А, г) или в антеизо- конфигурации (рис. 9, А, д);
— ω- алициклические жирные кислоты (рис. 9, А, ж).
Они также синтезируются из соответствующего праймера и малонил-СоА в качестве удлинителя цепи. В случае необходимости они подвергаются модификации (десатурации или гидроксилированию). Грамположительные бактерии, в частности представители рода Bacillus обычно содержат больше терминально разветвленных жирных кислот, чем грамотрицательные бактерии.
Одна из старых догм липидологии гласила, что у бактерий, в отличие от растений и животных, отсутствуют полиненасыщенные жирные кислоты. Однако в настоящее время у морских бактерий Cowliella psychrerythraea, Flexibacer polymorphus и Shewanella gelidimarina, а также у ряда цианобактерий обнаружены поли-cis-ненасыщенные жирные кислоты (рис. 9, А, б), в частности эйкозопентаеноевая кислота (С20 : 5, ∆5, 8, 11, 14, 17).
Согласно другой догме, у бактерий отсутствуют trans-мононенасыщенные жирные кислоты. Однако в настоящее время они найдены у Pseudomonas putida, которая образует их путем прямой изомеризации (рис. 9, А, в).
В отличие от эукариотов, у которых изменение физических свойств липидного ядра мембраны происходит за счет изменения соотношения между фосфолипидами
и холестеролом, бактерии используют другую стратегию — варьируют жирнокислотный состав липидов.
Способность бактерий варьировать жирнокислотный состав мембранных липидов в зависимости от изменений условий окружающей среды для того, чтобы мембраны находились в жидкостно-кристаллическом состоянии, называется «гомеовязкостной адаптацией» (англ. homeoviscous adaptation). Благодаря этому бактерии поддерживают вязкость своих мембран на относительно постоянном уровне.
У бактерий, которые самостоятельно синтезируют жирные кислоты (прототрофов по жирным кислотам), это происходит путем изменения набора соответствующих ферментов. В то же время для бактерий, нуждающихся в экзогенных жирных кислотах (ауксотрофов по жирным кислотам, например, Acholeplasma laidlawii) температура, при котоpой возможен рост, зависит от типа добавленной жирной кислоты.
Первая задача мембраны — быть барьером для гидрофильных молекул, что не только не противоречит статичности этой структуры, но напрямую зависит от нее. Вторая задача мембраны — служить матриксом для функциональных белков — требует внутренней динамичности, чтобы они могли свободно плавать или хотя бы смещаться в латеральном, а также трансверсальном направлениях. Для физиологической активности мембраны одинаково губительны как «замороженное» кристаллическое состояние, так и «расплавленное» жидкое состояние.
В случае фазового перехода при понижении температуры функциональные белки «вымораживаются», т. е. в результате латеральной диффузии вытесняются из липидного слоя и случайно группируются в участках, свободных от липидов. В свою очередь, липиды с более низкой температурой фазового перехода образуют «незамерзшие» домены, которые остаются в жидком состоянии.
В случае фазового перехода при повышении температуры мембрана разжижается, и скорость диффузии ее компонентов возрастает. В конечном счете, контакты между функциональными белками становятся слишком кратковременными и хаотичными.
При температуре, когда происходит рост, мембрана обязательно должна содержать участки в промежуточном жидкостно-кристаллическом состоянии, хотя в ней могут быть относительно большие кристаллические зоны.
Обратимый переход мембранных липидов из кристаллического в жидкое состояние зависит от температуры плавления жирных кислот. Однако в физиологическом отношении важнее температура фазового перехода липидного матрикса. В зависимости от преобладающего типа жирной кислоты она ниже температуры ее плавления на 10-45° С.
Липидный слой с плотно упакованными длинными и «прямыми» молекулами характеризуется более высокой упорядоченностью и повышенной жесткостью, и поэтому он плавится при более высокой температуре.
Более высокую температуру фазового перехода имеют липиды, образованные:
— длинноцепочечными жирными кислотами (по сравнению с короткоцепочечными);
— насыщенными жирными кислотами (по сравнению с ненасыщенными);
— trans-мононенасыщенными жирными кислотами (по сравнению с моно-cis-ненасыщенными и поли-cis-ненасыщенными);
— разветвленными жирными кислотами (по сравнению с неразветвленными);
— ациклическими жирными кислотами (по сравнению с циклическими).
Поэтому при более низкой температуре прежде всего увеличивается относительное содержание ненасыщенных жирных кислот. В свою очередь, длина жирнокислотной цепи прогрессивно возрастает у мезофильных, термофильных и экстремально термофильных бактерий (С15) —» (С15-С17) —» (С17-С19).
Текучесть мембраны уменьшается не только при снижении температуры, но и при других воздействиях, которые сокращают расстояние между фосфолипидными молекулами — например, при повышении гидростатического давления или при добавлении стероидов к фосфатидам. У барофильных бактерий при повышении гидростатического давления увеличивается относительное содержание ненасыщенных жирных кислот.
Ферменты биосинтеза гицеролипидов используют растворимые субстраты, в частности 3-фосфоглицерол. Поскольку эти ферменты находятся на PS-поверхности СМ, возникает проблема, как перенести готовый продукт в наружный полумембранный листок. Спонтанно это происходит довольно медленно, и для ускорения используются «переворачивающие» ферменты — флиппазы.
В связи с ключевой ролью мембран в жизни бактериальной клетки, большое фармакологическое значение имеют антибиотики, мишенью для которых служит биосинтез липидов (тиолакто- мицин, церуленин и др.).
Фосфатиды. Основой полярных диацилглицеридов является фосфатидная кислота (англ. phosphatidic acid, РА), которая содержит жирнокислотные остатки в положении 1 и 2, а остаток фосфорила в положении 3. В свободном виде фосфатидной кислоты обычно мало, в основном она образует замещенные по фосфорилу производные — фосфатиды (рис. 9, Б).
Три самых распространенных у бактерий фосфатида — это фосфатидилглицерол (англ. phosphatydyl glycerol, PG), дифосфатидилдиглицерол (англ. diphosphatydyl diglycerol, DPDG; кардиолипин) и фосфатидилэтаноламин (англ. phosphatydyl ethanolamine, РЕ; кефалин). Фосфатидилглицерол преимущественно синтезируют неспорообразующие грамположительные бактерии, а фосфатидилэтаноламин — грамотрицательные бактерии.
Менее распространен фосфатидилсерин (англ, phosphatydyl srerine, PS) — например, он известен у представителей рода Rhizobium, а также фосфатидилхолин (англ. phosphatydyl choline, PC; лецитин).
Еще реже встречается фосфатидилинозитол (англ. phosphatydyl inosite, PI; инозитид), который играет важную роль во внутриклеточном сигналинге у эукариотов. Тем не менее, у некоторых бактерий, например, из рода Mycobacterium, это главный мембранный липид.
От количественного соотношения между анионными полярными головками фосфатидилглицерола и дифосфатидилглицерола и катионными полярными головками фосфатидилэтаноламина и фосфатидилхолина зависит величина избыточного отрицательного заряда на обеих поверхностях СМ.
Плазмалогены. Многие анаэробные бактерии образуют 1-алк-енил-2-ацилгли- цериды, или плазмалогены (рис. 9, В), которые представляют собой внутримолекулярную комбинацию винилового эфира глицерола с алифатическим альдегидом и сложного эфира глицерола с жирной кислотой. Существуют и неацилированные варианты, а также варианты с двумя ацильными остатками в положении 2 и положении 3.
Плазмалогены, впервые обнаруженные у живущей в рубце жвачных животных анаэробной бактерии Ruminococcus flavefaciens, бывают и у грамотрицательных, и у грамположительных бактерий.
Моноалкилфосфоглицериды. Еще реже у анаэробных бактерий встречаются 1-алкил-2-ацил-фосфоглицериды — обычные компоненты рыбьего жира (рис. 9, Г). В данном случае глицерол в положении 1 образует простую эфирную связь с неразветвленным алифатическим спиртом, а в положении 2 находится остаток жирной кислоты. В качестве этерифицирущего спирта могут выступать химиловый (гексадециловый), батиловый (октадециловый) и селахиловый (октадек-cis-енил) спирты. Ацильные заместители находятся в положении 2 или в положении 2 и 3.
Уникальную группу моноалкилглицеридов образуют некоторые варианты недавно открытых уникальных липидов — ледеранов (см. раздел 8.3.11.1).
Диалкилфосфоглицериды. Исключительно редко бактерии образуют 1,2-ди- алкилглицериды (рис. 9, Д). В этом случае к глицеролу в положении 1 и положении 2 с помощью простой эфирной связи присоединяются изо- или антеизо-разветвленные алифатические спирты. Обладателями таких липидов являются термофильные сульфатредуцирующие бактерии из рода Thermodesulfobacterium (фила ВIII Thermodesulfobacteria).
Нейтральные ацилглицериды и ацилглюкозиды. Такие липиды являются полярными, но электронейтральными.
Примерами нейтральных ацилглицеридов служат полностью замещенный 1,2,3- триацил-sn-глицерид (рис. 9, Е), частично замещенный диацил-sn-глицерид, 1-моно- ацил-sn-глицерид и 3-моноацил-sn-глицерид, а также полностью замещенный диацилгликозилглицерид (рис. 9, Ж).
В свою очередь, нейтральные ацилглюкозиды могут содержать от одного до трех ацильных заместителей (рис. 9,3). Такие липиды у бактерий встречаются довольно редко.
Стероиды (циклические полиизопреноидные липиды). В отличие от ацилглицеридов и ацилглюкозидов, гидрофобный домен полиизопреноидных липидов представлен не ацильными остатками, а состоит из циклических или из регулярно разветвленных ациклических структур. Для их биосинтеза используются изопреновые C5-единицы (-СН2-С(СНз)=СН-СН2-).
Циклические полиизопреноиды из группы стероидов типичны для эукариотов. Напротив, прокариоты синтезируют их исключительно редко. Известны всего три достоверных примера — метилотрофная бактерия Methylococcus capsulatus, а также миксобактерии Nannocystis exedens и Polyangium sp., которые самостоятельно синтезируют стероиды, причем в таких же количествах, как и эукариоты. Способность некоторых штаммов Azotobacter sp. и Synechococcus sp. синтезировать стероиды требует подтверждения.
Некоторым бактериям необходимы экзогенные стероиды, которые спонтанно включаются в состав СМ. Редкими примерами этого служат представители родов Helicobacter, Mycoplasma и Treponema.
Стероиды, в частности С27-холестерол представляют собой тетрациклические соединения с боковой полиизопреноидной цепью в положении 17 (рис. 10, А). Благодаря тому, что холестерол содержит полярный гидроксильный радикал в положении 3, он ориентируется в липидной фазе мембраны точно так же, как это делают фосфатиды, причем тетрациклическая часть молекулы по размеру соответствует гидрофобному «хвосту» фосфолипида.
Рис. 10. Циклические полиизопреноидные липиды. А — стероиды; Б — гопаноиды.
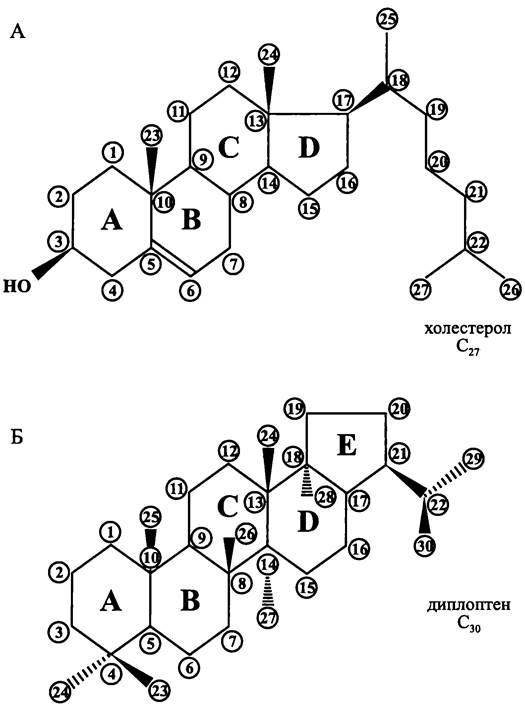
Стероиды играют важную роль в гомеовязкостной адаптации. Два гидрофобных «хвоста» в молекуле фосфатида обладают гибкостью, которая увеличивается с повышением температуры. В итоге мембрана разжижается и, в конечном счете, теряет свои физиологические функции. Плоские молекулы стероидов, напротив, обладают жесткостью. Они могут компенсировать гибкость фосфатидов и их значительную диффузию.
Гопаноиды (циклические полиизопреноидные липиды). Хотя бактерии не содержат стероидов, у них широко распространены суррогаты стероидов — гопаноиды (англ. hopanoid). Название этих пентациклических углеводородов происходит от того, что их первый представитель, гидроксигопанон был выделен из живицы тропического дерева Нореа (сем. Dipterocarpaceae), в свою очередь, названного в честь английского ботаника XVIII в. Джона Хоупа (J.Hope). Источником гопаноида у этого представителя криптогамных служат мембраны хлоропластов. Поскольку хлоропласты произошли от цианобактерий, это не эукариотный, а бактериальный липид.
Гопаноиды широко распространены у грамположительных и грамотрицательных бактерий; исключением являются энтеробактерии, пурпурные бактерии и некоторые облигатные анаэробы. У архей гопаноиды не обнаружены. Накопление гопаноидов коррелирует с высоким содержанием ГЦ-пар (> 50%). Поскольку такой состав ДНК свойствен бактериям в «стрессовых» экологических нишах, гопаноиды можно рассматривать как своего рода «стрессовые» молекулы.
Простейшим представителем гопаноидов является С30-гопаноид диплоптен (рис. 10, Б), обнаруженный, в частности, у цианобактерий, а также у Methylococcus capsulatus и В. subtilis.Структура С30-гопаноидов может усложняться путем введения С5-алкильного заместителя в положение 30. Кроме того, может изменяться число и позиция гидроксилов полярной боковой цепи в положении 29, а также может появляться метильная группа в положении 2 или положении 3 или двойная связь в положении б и положении 11.
Аналогично стероидам, гопаноиды участвуют в гомеовязкостной адаптации. Они ориентируются вдоль молекул фосфолипидов (их полярная «головка» находится на поверхности мембраны) и повышают упорядоченность мембраны при физиологической температуре, стабилизируя двойной липидный слой в квази-кристаллическом состоянии.
Сквален и другие ациклические каротиноиды. Мембраны всех бактерий, за исключением актиномицетов, а также представителей родов Staphylococcus и Streptococcus, содержат очень мало ациклического сквалена (рис. 11, В, а). Этим они принципиально отличаются от архей, у которых сквален и его гомологи являются характерными неполярными липидами (см. ниже).
В то же время сквален используется в качестве предшественника для биосинтеза стероидов и гопаноидов. Они образуются путем его циклизации или циклизации его эпоксида.
Сквален представляет собой С30-каротиноид, или «тритерпен» (греч. terebinthos — скипидарное дерево). Он собирается из двух, остатков фарнезилпирофосфата (С15) на боковой ветви генерального пути биосинтеза С20- и С40-каротиноидов:
СНзСО ~ SСоА —> —> —> С5 (изопентенилпирофосфат СН2=С(СНз)-СН2-СН2 ~ РР) —> С10 (геранилпирофосфат, или «монотерпен») —>
С15 (фарнезилпирофосфат, или «сесквитерпен») —> С20 (геранилгеранилпирофосфат, или «дитерпен») —> С40 (фитоин, или «тетратерпен»).
Терпеноиды могут модифицироваться путем циклизации, гидроксилирования и эпоксидирова- ния, что приводит к очень большому разнообразию этих соединений (>2 • 104вариантов).
Наиболее важное превращение терпеноидов связано с одним из альтернативных механизмов — десатурацией (уменьшением степени насыщенности) и сатурацией (повыщением степени насыщенности). В результате десатурации синтезируются универсально распространенные окрашенные каротиноиды. В свою очередь, полная сатурация приводит к образованию регулярно разветвленных алканов — фитанов, которые образуют боковые цепи уникальных липидов архей (см. раздел 8.3.3.2).
Помимо сквалена, в состав двойного липидного слоя могут входить и другие ациклические каротиноиды. Они изучены слабо; возможно, что вместе со скваленом они принимают участие в гомеовязкостной адаптации.
8.3.3.2. Мембранные липиды архей
Вспомогательные мембранные липиды архей сходны с мембранными липидами бактерий и эукариотов, причем во всех трех случаях это свободные полиизопреноиды. Подобно бактериям, архей могут включать в состав своих мембран линейные полиизопреноиды — сквален и его гомологи. Однако они никогда не образуют полициклических полиизопреноидов — стероидов и гопаноидов.
В то же время основная часть мембранных липидов архей уникальна. Это простые эфиры, содержащие регулярно разветвленные, полностью насыщенные боковые цепи. Такие липиды обладают повышенной рекальцитрантностью, поскольку простые эфиры устойчивее к гидролизу, чем сложные, а полностью насыщенные и
регулярно разветвленные углеводородные цепи противостоят расщеплению и окислению при высокой темературе.
Археолы и кальдархеолы. Напомним, что полярные липиды бактерий и эукариотов образуются на основе фосфатидной кислоты. В ее состав входят сложные эфиры жирных кислот или, в очень редких случаях, простые эфиры глицерола с алкановыми спиртами (см. раздел 8.3.3.1).
В отличие от этого, полярные липиды архей образуются на основе особых глицеролипидов — «археолов» и «кальдархеолов», которые представляют собой простые эфиры глицерола или простые эфиры 9-атомного спирта нонитола с полиизопреноидными спиртами. В данном случае гидрофобные «хвосты» регулярно разветвлены и состоят из 20, 25 или 40 атомов углерода (рис. 11).
Археолы (англ. archaeol; «спирт архей») представляют собой простые диэфиры глицерола и полиизопреноидных спиртов, прототипом которых служит С20-спирт фитанол. Они содержат два коротких «хвоста», каждый из которых состоит из 20-25 атомов углерода. В редких случаях два С20-«хвоста» соединяются концами. Наиболее распространенным археолом является 2,3-ди-О-фитанил-sn-глицерол (англ. diphytanyl glyceride diether, DGD). Его боковые цепи состоят из 20 атомов углерода (рис. 11 А, а). Важно подчеркнуть, что, в отличие от фосфатидов, гидрофобные хвосты связаны с глицеролом простой эфирной связью не в положении 1 и 2, а в положении 3 и 2 (еще раз напомним, что по стереоспецифической нумерации в проекции Эмиля Фишера верхний атом углерода идет под № 1, а вторичная гидроксильная группа обращена влево от вертикальной оси молекулы). Поэтому археолы являются не левовращающими D-изомерами, а правовращающими L-изомерами.
Чаще всего обе боковые цепи археолов состоят из четырех восстановленных изопреновых (метил-бутадиеновых) субъединиц со структурой -СН2-СН(СНз)-СН2-СН2-. Однако бывают и такие варианты, когда в одной или обеих цепях число изопреновых единиц возрастает на одну (С2О, С25- археолы и С25, С25-археолы; рис. 11, А, а). Иногда в одной или в другой С20-цепи при третьем атоме углерода (если считать от эфирной связи) появляется гидроксильный радикал. Наконец, две С20-цепи могут соединяться концами, и тогда образуется моноциклический диглицерид (англ. monocyclic glyceride diether, MGD; рис. 11, А, б).
Рис. 11. Мембранные липиды архей. DGD — дифитанилдиглицерид; МGD — моноциклический диглицерид; DGT — дибифитанилтетраглицерид; СDТ — циклизированный дибифитанилтетраглицерид. Пояснение для структур а — е («Б») и а — г («В») в тексте.
Археолы составляют до 100% изопреноидных эфиров у экстремально галофильных архей, а также у метаногенных кокков (фила АII Euryarchaeota). Напротив, у представителей рода Thermoplasma (фила All Euryarchaeota), а также у серазависимых гипертермофилов (фила AI Crenarchaeota) их количество не превышает 5% общего содержания глицероэфиров.
Кальдархеолы (лат. caldus — горячий; англ. caldarchaeol; «спирт термофильных архей») представляют собой простые тетраэфиры глицерола и полиизопреноидных спиртов. Они содержат длинные боковые цепи, которые образуются путем ковалентного соединения «голова к голове» боковых цепей двух диэфиров. При этом первичные гидроксилы находятся в frans-конфигурации.
Чаще всего происходит попарное соединение, и тогда образуется тетраэфир — 2,3,2,3-тетра- О-дибифитанил-ди-sn-глицерол (англ. diphytanyl glyceride tetraether, DGT), боковые цепи которого состоят из 40 атомов углерода. Иногда связь образуется только в одной паре (рис. 11, А, в). В результате усложнения структуры кальдархеолов внутри ациклической полиизопреноидной цепи появляются циклопентановые кольца, и в этом случае образуются циклизированные дифитанильные тетраэфиры (англ. cyclic diphytanyl glyceride tetraether, CGT). Число колец в каждой цепи варьирует от одного до четырех, причем они могут располагаться как симметрично, так и асимметрично. Другим усложнением является замена одного из глицеролов на уникальный 9-атомный спирт нонитол, или кальдитол (рис. 11, А, г).
Кальдархеолы составляют до 90% изопреноидных эфиров у представителей родов Thermoplasma и Sulfolobus, а также у термофильных метаногенов. Циклизированные тетраэфиры встречаются только у Thermoplasma и гипертермофильных архей, а смешанные тетраэфиры глицерола и кальдитола — только у гипертермофилов из филы AI Crenarchaeota.
Мембрана, состоящая из кальдархеолов, характеризуется уникальным строением — в ее основе лежит не двойной липидный слой, а амфипатический монослой, в котором различают гидрофобную сердцевину и две гидрофильные полярные поверхности. В отличие от обычных унитарных мембран (см. рис. 8), такая мембрана не образует внутренних поверхностей разлома.
Тетраэфиры более упорядочены, чем диэфиры, и поэтому при совместном существовании в мембране они сегрегируются в различных ее участках, т. е. мембрана становится латерально анизотропной. Монослой значительно стабильнее двойного слоя и менее проницаем для ионов, в частности протонов.
Полярные липиды архей. Полярные «головки» археотных липидов могут быть заряженными или электронейтральными. В целом они аналогичны полярным головкам липидов бактерий, но более разнообразны. Они присоединяются к археолу или кальдархеолу в положении 1 через фосфорильный остаток (рис. 11, Б, а-д) или напрямую (рис. 11, Б, е).
В качестве фосфорзамещенной полярной головки могут выступать серин (рис. 11, Б, а), глицерол (рис. 11, Б, б), глицерофосфат или глицеросульфат (рис. 11, Б, в) и этаноламин (рис. 11, Б, г), а также разнообразные полиолы и сахара — инозитол (рис. 11, Б, д), аминопентатриол, галактоза, глюкоза и манноза.
Напрямую присоединяются к археолу или к кальдархеолу моно-, ди- и тригликозильные группировки. Они могут быть дополнительно фосфорилированы или сульфатированы, а также могут содержать вторичные гликозильные остатки или вторичные остатки этаноламина (рис. 11, Б, е).
Thermoplasma, у которой отсутствует клеточная стенка, содержит липополисахариды с длинными и разветвленными полярными цепями (см. раздел 8.5.1.2).
Неполярные липиды архей. Неполярные липиды составляют не более 20% общего количества липидов в СМ архей. Еще раз напоминаем, что архей не синтезируют стероидов и гопаноидов, и их неполярные липиды представлены ациклическим скваленом (рис. 11, В, а).
Наряду со скваленом, существуют его ди-, тетра-, гекса-, окта- и декагидропроизводные, в частности сквалан (рис. 11, В, б). Кроме того, встречаются гомологи сквалена с цепью из 15-30 атомов углерода, например, сестертерпанан (рис. 11, В, в) и фарнезан (рис. 11, В, г).
Гомеовязкостная адаптация у архей. Поскольку архей не образуют фосфатидов и не могут синтезировать стероидов и гопаноидов, им приходится осуществлять гомеовязкостную адаптацию иным способом, чем это делают бактерии или эукариоты.
При относительно низкой температуре в СМ увеличивается количество «разжижающих» компонентов — сквалена и его частично восстановленных производных. В свою очередь, при высоких температурах возрастает содержание тетраэфиров, что делает мембрану более упорядоченной и повышает температуру фазового перехода.
За счет образующихся циклопентановых колец изменяется толщина СМ, а также ограничивается вращение бифитанильных цепей. Вязкость и ригидность липидного монослоя повышаются. Это способствует не только выживанию архей при экстремальных температурах, но и обеспечивает их оптимальный рост в условиях, где не могут существовать другие живые организмы.
8.3.3.3. Значение мембранных липидов для палеомикробиологии
В разделе 1.8 уже шла речь о биомолекулах, которые обладают высокой устойчивостью к разрушению и, в отличие от легко уязвимых нуклеиновых кислот и белков, неограниченно долго сохраняются в постмортальном состоянии.
К числу наиболее рекальцитрантных биомолекул относятся мембранные липиды (или их компоненты). Благодаря тому, что они имеют разветвленную и/или циклическую структуру и не содержат или содержат очень мало двойных связей, они не разрушились и почти без изменений просуществовали до наших дней.
Именно из таких липидов состоит нерастворимый органический углерод осадочных пород — известняков, сланцев и глин — кероген (лат. сего — натирать мазью и греч. genea — происхождение; «источник маслянистого вещества»). Кроме того, такие липиды входят в состав фракции твердых углеводородов угля и фракции жидких углеводородов нефти.
Возраст этих «молекулярных ископаемых» достигает 1,5 млрд лет, а их общее количество оценивается в 1012 т (по углероду), что равно содержанию углерода в биомассе современных организмов.
Ископаемые липиды имеют специфическое строение, и их аналоги содержатся в биомембранах. Более того, иногда тот или иной тип «молекулярного ископаемого» находят раньше, чем его аналога у живых организмов. Применительно к этим случаям используется образный термин «молекулярный целакант» (англ, molecular coelacanth; представитель этой группы кистеперых рыб — Latimeria — до 1938 г. был известен только по ископаемым остаткам).
Поскольку бактерии, археи и эукариоты различаются по составу мембранных липидов, для каждого из этих доменов существует свой тип «молекулярного ископаемого». Для бактерий это гопаны (продукты спонтанного восстановления гопаноидов), для архей — фитаны (продукты спонтанного восстановления спиртовых «хвостов» у археолов и кальдархеолов), а для эукариотов — стераны (продукты спонтанного восстановления стероидов).
Таким образом, определяя геологический возраст, места залегания и состав осадочных пород, мы можем судить об архаичности, географическом распределении и разнообразии когда-то существовавших микроорганизмов.
8.3.4. Цитоплазматическая мембрана
Среди мембран прокариотной клетки важнейшее место принадлежит цитоплазматической мембране (СМ). Она не только обеспечивает структурную целостность организма, но и представляет собой полифункциональную органеллу.
СМ поддерживает осмотический баланс цитоплазмы с окружающей средой, осуществляет разные виды транспорта (в том числе, секрецию белка; см. раздел 8.6.1), участвует в биогенезе клеточной стенки и биосинтезе внеклеточных полимеров (см. раздел 8.5), а также рецептирует внешние регуляторные сигналы.
Помимо всех перечисленных функций, которые у прокариотов и эукариотов одинаковы, СМ прокариотной клетки может ассимилировать энергию в форме трансмембранного электрохимического потенциала, или протондвижущей силы (англ. proton motive force, Pmf). Наконец, CM принимает участие в генетических процессах — репликации и сегрегации дочерних хромосом, а также в векторном транспорте ДНК при вирусной инфекции или половом процессе, протекающем по сценариям генетической трансформации и конъюгации. Таким образом, с ней ассоциированы все важнейшие функции прокариотной клетки.
Прокариотная СМ имеет черты специализации, по отдельности свойственные:
— СМ эукариотов (импорт субстратов, экспорт метаболитов, секреция);
— ICM эукариотов (участие в структурной организации, копировании и экспрессии генетического материала; участие в биосинтезе липидов, а также в процессинге и таргетинге белков);
— ICM митохондрий или пластид (ассимиляция энергии).
Прокариотная СМ выполняет сильнее выраженную «экстравертную» роль, чем СМ ядерных клеток. Действительно, у бактерий и архей важнейшие генетические, метаболические и транспортные процессы ассоциированы с клеточной оболочкой (а не с мембранами вакуома, как в случае протистов, грибов, животных и растений). Метафорически это можно выразить следующим образом — ядерные клетки «скрывают» от собственного цитоплазматического компартмента процессы, локализованные в нуклеоплазме, а биосинтетические и транспортные процессы на уровне ER и аппарата Гольджи протекают в состоянии «внутренней эмиграции».
Хорошо изучена СМ цианобактерий, хотя для этого пришлось преодолеть трудности методического характера, поскольку это минорный компонент мембранного материала (<1%клеточного белка); кроме того, фрагменты СМ нелегко отделить от фрагментов ICM — мембран тилакоидов.
В результате анализа, проведенного автором учебника в конце 1970-х годов, а позднее австрийским биохимиком Гюнтером Пешеком (G. Peschek) было показано, что СМ цианобактерий содержит до 40 полипетидов и характеризуется низким содержанием Mg-порфиринов и высоким содержанием каротиноидов (среди них больше всего, 60% зеаксантина и меньше всего, 3% β-каротина). Каротинопротеин с молекулярной массой 40-45 кДа выполняет фотопротекторную функцию.
Поскольку СМ не содержит хлорофилла, она непосредственно не участвует в фототрофии, однако способствует этому процессу (среди ее компонентов обнаружен предшественник хлорофилла а — хлорофиллид а). Кроме того, в составе СМ обнаружены полипептиды, функционирующие в полости тилакоида (в частности, это Mn-стабилизирующая субъединица кислородвыделяющего комплекса, а также минорные субъединицы ФСI — белки PsaD и PsaE). Этот факт можно объяснить либо «неразборчивым» таргетингом белков ICM, либо тем, что ICM непосредственно сообщается с СМ.
Установлено, что СМ содержит F0F1-АТФазу, хотя неясно, действует этот фермент обратимо или только гидролизует АТФ. Помимо этого, СМ содержит Р-АТФазу, которая не транслоцирует протоны, а катализирует АТФ-зависимый перенос катионов Са2+ (что весьма необычно для бактерий, хотя АТФазы данного типа широко распространены у высших растений и грибов).
8.3.5. Интрацитоплазматические мембраны
Напомним, что клетки усложненного бактериального морфотипа имеют бимембранную оболочку, а также интрацитоплазматические мембраны (ICM). Биогенез ICM осуществляется адаптивно, в частности, у пурпурных бактерий при низком парциальном давлении кислорода. В редких случаях ICM сохраняются в любых физиологических состояниях (примером служат цианобактерии).
В учебниках по микробиологии сведения о бактериальных ICM даются конспективно; кроме того, при описании строения, функционирования и биогенеза этих структур повторяются характерные ошибки. Прежде всего, тезис о «простом» строении бактериальной клетки, якобы выраженном в отсутствии ICM, противоречит
фактам (они закономерно представлены у многих бактерий). Другая ошибка связана с тем, что ICM рассматривают не как дифференцированные структуры, а как складки СМ. Наконец, утверждается, что ICM возникают путем инвагинаций СМ.
Самый распространенный дизайн ICM — это глубоко погруженные в цитоплазму мембранные цистерны. Реже это немногочисленные трубки или полусферы, связанные с СМ. Хотя в последнем случае ICM морфологически слабо выражены, все же это дифференцированные структуры, и их следует отличать от «ворсинок» СМ у гигантской бактерии Epulopisciumfishelsoni (см. раздел 8.1.3), за счет которых увеличивается площадь поверхности СМ.
Общие закономерности образования ICM. Подобно тому, как интенсивность транспортных процессов зависит от величины поверхностно-объемного коэффициента (см. раздел 8.1.3.1), фактором, критичным для ассимиляции энергии, является относительный размер поверхности СМ.
В качестве примера рассмотрим фототрофию. Ее эффективность зависит от площади мембраны, поглощающей свет и ассимилирующей энергию его квантов. При отсутствии ICMфототрофию можно было бы интенсифицировать за счет увеличения поверхностно-объемного коэффициента. Однако такая морфогенетическая стратегия почему-то не используется фототрофными бактериями, среди которых, за исключением Rhodomicrobium vannielii, нет мицелиальных форм. В онтогенезе эта стратегия также не получила распространения (единственное исключение — Rhodospirillum tenue, которая при слабом освещении образует длинные тонкие клетки с более высоким, а на ярком свету короткие толстые клетки с более низким поверхностно-объемным коэффициентом). В основном же используется стратегия «интернирования» фотосинтетических мембран, т. е. их размещение внутри клетки. Естественно, что ICM по причине внутренней локализации не выполняют пограничных функций — таких, как поглощение субстратов, экспорт, рецепция сигналов и т.д.
Генераторы Pmf занимают много места в мембране. Линейный размер каждого из них примерно равен толщине билипидного слоя, а у многих фототрофов он увеличивается за счет светособирающего комплекса. Поэтому фотосинтетический аппарат в ходе эволюции закономерно подвергался интернированию. В результате клетки бывают «набиты» ICM, площадь которых многократно (иногда на 2-3 порядка) превышает площадь поверхности СМ.
Однако фототрофные бактерии не обязательно образуют ICM. Примерами служат все зеленые серные бактерии (фила BXI Chlorobi), все гелиобактерии (фила ВХIII Firmicutes), большинство зеленых несерных бактерий (фила BVT Chloroflexi) и несколько видов пурпурных несерных бактерий (фила ВХII Proteobacteria). Среди цианобактерий (фила ВХ Cyanobacteria) не имеет тилакоидов только Gloeobacter violaceus.
Связано ли образование ICM с генеральными структурно-функциональными свойствами бактерий? Если такая связь существует, то насколько она закономерна?
Факты говорят о тройственной системе корреляций, связывающей способность к образованию ICM с морфотипом и энергетическим метаболизмом (рис. 12).
Первая корреляция связывает способность к образованию ICM с основным бактериальным, или грамотрицательным морфотипом. Бактерии, обладающие грамположительным морфотипом, ICM не образуют (табл. 8).
Рис. 12. Система корреляций, связывающая способность к образованию интрацитоплазматических мембран с морфотипом и энергетическим метаболизмом. ICМ — интрацитоплазматические мембраны.

Данную корреляцию можно объяснить тем, что открытые в периплазматическое пространство мембранные цистерны могут существовать лишь в том случае, если внутри них поддерживается осмотическое давление, противодействующее тургорному давлению цитоплазмы. Такой «контртургор» действительно существует. Он обеспечивается накоплением в периплазматическом пространстве осмотически активных соединений — уникальных полигликозидов, ковалентно связанных с остатками сукцинила, фосфоглицерола и фосфоэтаноламина (см. раздел 8.4.2). С помощью регуляторного каскада, который начинается с осмосенсора, концентрация этих осмолитиков регулируется таким образом, что обеспечиваемый ими тургор в сумме с осмотическим давлением окружающей среды уравновешивает тургорное давление цитоплазмы. Бактерии, обладающие грамположительным морфотипом, лишены периплазматического пространства и не образуют ICM, поскольку не в состоянии обеспечить их «осмотический комфорт».
Вторая корреляция связывает способность к образованию ICM с автотрофным энергетическим метаболизмом. В данном случае это означает ассимиляцию световой энергии или энергии окисления неорганического субстрата. Выше мы уже говорили о том, что для интенсификации этого способа питания необходимо увеличить количество систем, создающих Pmf. Когда для них уже не хватает места в СМ, проблема «перенаселения» решается путем интернирования и специализации мембран, т. е. путем образования ICM.
Третья корреляция связывает грамотрицательный морфотип с автотрофным энергетическим метаболизмом. Из-за маленького размера пор, имеющихся в ОМ (см. раздел 8.4.1.2), периплазматическое пространство закрыто для экзогенных органических молекул размером >0,6 кДа. По этой же причине ограничивается секреция гидролитических ферментов, что не позволяет питаться продуктами гидролиза экзогенных полимеров. Таким образом, из-за свойств оболочки грамотрицательных бактерий давление отбора направлено в сторону уменьшения гетеротрофного потенциала. Характерно, что грамположительные бактерии, не имеющие этого ограничения, редко проявляют способность к автотрофному энергетическому метаболизму. В качестве исключений назовем фототрофа Heliophilum fasciatum и таких хемолитотрофов, как Bacillus tusciae, Clostridium mayombei и Mycobacteriumgastri.
Мезосомы прокариотов — артефактные ICM. В связи со сказанным рассмотрим «странные» случаи, когда ICM обнаруживаются у бактерий, обладающих грамположительным морфотипом. В первую очередь упомянем мезосомы — полиморфные структуры, получившие свое название из-за преимущественнй локализации в экваториальной области клетки (греч. mesos — находящийся в середине и soma — тело).
Таблица 8. ICM бактерий; распространение и характеристики
Морфофизиологическая группа; фила; представитель |
Строение |
Функция |
Тип и регуляция биогенеза |
Фототрофные бактерии |
|||
Пурпурные; фила ВХII- Proteobacteria; Rhodomicrobium vannielii(класс «Alphaproteobacteria»), Rhodocyclus purpureus (класс «Betaproteobac- teria»), Ectothiorhodospira halochloris (класс «Gammaproteobacteria») |
(Не) агрегированные сферы; (не)агрегированные трубки; стопки дисков; стопки ламелл |
Фототрофия; аэробное дыхание |
«Перманентное развитие»; «ступенчатое развитие» в обратной зависимости от интенсивности освещения; «индукция» при низком парциальном давлении кислорода |
Зеленые нитчатые; систематическое положение неизвестно; «морские Chlo- roflexus-подробные организмы» |
Неагрегированные трубки |
Фототрофия; аэробное дыхание |
- |
Гелиобактерии; филаВХIII Firmicutes; Heliobacterium chlorum (класс «Clostridia») |
Неагрегированные трубки |
- |
- |
Тривиальные цианобактерии; фила ВХ Cyanobacteria; Synechococcus sp. (пор. « Chroococcales » ) |
Неагрегированные тилакоиды |
Фототрофия; аэробное дыхание; трансляция на «шероховатых» тилакоидах |
«Перманентное развитие» ;«ступенчатое развитие» в обратной зависимости от интенсивности освещения |
Прохлорофиты; фила ВХ Cyanobacteria; Prochlorococcus marinus(пор. «Chroococcales») |
(Не) агрегированные тилакоиды |
Фототрофия |
«Перманентное развитие» |
Квази-фототрофные бактерии |
|||
Аэробные аноксигенные; фила ВХII Proteobacteria Erythrobacterlongus (класс « Alphaproteobacteria») |
Неагрегированные сферы |
Фототрофия |
«Индукция» при умеренном парциальном давлении кислорода и прерывистом освещении или в темноте |
Хемолитотрофные бактерии |
|||
Нитрифицирующие; фила ВХII Proteobacteria; Nitrо- bacterwinogradskyi (класс «Alphaproteobacteria»), Nitrosomonas europaea(класс «Betaproteobacteria»), Nitrosococcus oceanus (класс «Gammaproteobacteria») |
Неагрегированные трубки; стопки дисков; стопки ламелл |
Аммиакмонооксигеназная реакция; создание Pmf |
«Перманентное развитие»; «ступенчатое развитие» в прямой зависимости от снабжения минеральными веществами |
Анаэрбно окисляющие аммиак бактерии; фила ВХVPlanctomycetes; Сфтвшвфегы «Brocadia anammoxidans » (класс «Planctomycetacea») |
Вакуоль (анаммоксосома) |
Компартментализация субстратов, участвующих в анаэробном окислении аммиака; создание Pmf |
|
Железобактерии; фила ВХII Рroteobacteria; Gallionellaferruginea(класс «Betaproteobacteria») |
Неагрегированные трубки или сферы |
- |
- |
Бесцветные серные; фила ВХII Proteobacteria; Thiomargaritanamibiense (класс III «Gammapro- teobacteria») |
Вакуоль (нитратная или кислородная) |
Запасание терминального акцептора электронов (анионов NO3-, О2 или элементной серы) |
«Перманентное развитие» ; «ступенчатое развитие» в прямой зависимости от снабжения сульфидом; «индукция» сульфидом и при уменьшении парциального давления кислорода |
Метанокисляющие типа I; фила BXII Proteobacteria; Methylophilusmethylotrophus (класс «Betaproteobac- teria»), Methylococcus cap- sulatus (класс «Gamma- proteobacteria») |
Неагрегированные трубки; стопки ламелл |
Метанмоноокси- геназная реакция; создание Pmf |
«Перманентное развитие»; «ступенчатое развитие» в прямой зависимости от парциального давления кислорода; «индукция» метаном |
Метанокисляющие типа II; Фила BXII Proteobacteria; Methylosinustrichosporium (класс «Alphaproteobac- teria») |
Неагрегированные сферы; неагрегированные ламеллы; стопки ламелл |
Метанмонооксигеназная реакция; создание Pmf |
«Перманентное» развитие; «ступенчатое развитие» в прямой зависимости от парциального давления кислорода; «индукция» метаном |
Хемоорганогетеротрофные бактерии |
|||
Магнитобактерии; фила BXII Proteobacteria; Magnetospirillum gryphiswal- dense (класс «Alphaproteo- bacteria») |
(Не) агрегированные сферы |
Участие в биогенезе магнитосом |
«Индукция» железом |
Уксуснокислые; фила BXII Proteobacteria; Gluconobac- ter oxydans(класс «Alpha- proteobacteria») |
Неагрегированные сферы или ламеллы |
- |
«Индукция» в стационарной фазе |
Планктомицеты; фила BXV Planctomycetes', Gemmataobscuriglobus (класс « Planctomycetacia ») |
(Не) агрегированные ламеллы |
Обеспечение квази-эукариотизма |
«Перманентное развитие» |
Фила BXX Bacteroidetes, Crenothrix polyspora (класс « Sphing obacteria ») |
Стопки ламелл |
- |
- |
Примечание: (-) — данные отсутствуют.
Пик интереса к мезосомам пришелся на 1960-1970 годы, когда их часто находили у бацилл, грамположительных кокков и актиномицетов, а также у метаногенных архей. Вначале мезосомы считали аутентичными структурами и приписывали им функции, аналогичные функциям специализированных органелл ядерных клеток. В частности, их рассматривали как аппарат для сегрегации хромосом (по аналогии с веретеном деления), место сосредоточения электрон-транспортной цепи (по аналогии с митохондрией или пластидой), а также депо гидролитических ферментов (по аналогии с лизосомой). В аналогичном контексте обсуждались функции «метано-
хондрий» (англ. methanochondrion) — мезосом метаногенных архей. Как следует из названия, их считали анаэробными аналогами митохондрий.
Мезосомы были «издержкой» при поисках компартментов в прокариотной клетке. Существовало мнение, что такие компартменты не должны отличаться от своих эукариотных аналогов. В настоящее время показано, что эти структуры имеют артефактную природу. Главной причиной образования мезосом служит грубая фиксация, при которой отдельные участки СМ меняют положение из-за нарушения осмотического барьера. В частности, ДНК-связывающий локус СМ увлекается коагулирующей хромосомой и смещается в центр клетки, в результате чего образуются экваториальные инвагинации тубуло-везикулярного, ламеллярного или смешанного типа. Неудивительно, что мезосомы перестали обнаруживать после перехода на щадящие методы приготовления электронно-микроскопических препаратов, в особенности при «сверхбыстром» (106 град • с-1) замораживании в струе пропана с криозамещением льда раствором осмия в спирте или ацетоне.
У некоторых хемоорганогетеротрофных грамотрицательных (рода Azotobacter и Gluconobacter) и грамположительных бактерий (р. Streptomyces) в 1970-е годы время от времени описывали ICM, которые по морфологии отличались от мезосом и не были расположены на экваторе клетки. Скорее всего, это были артефактные структуры, появление которых было также вызвано грубой фиксацией.
Морфология бактериальных ICM. Набор морфологических вариантов ICM очень беден, и каждый из них высококонсервативен. В частности, у пурпурных, нитрифицирующих и метилотрофных бактерий ICM представляют собой однообразные агрегаты везикулярных, тубулярных или дискоидных модулей. Уникальным исключением является пурпурная бактерия Rhodopseudomonas acidophila, которая одновременно образует ламеллярные и везикулярные хроматофоры.
Часто пишут, что ICM состоят из сферических (везикулярных), трубчатых (тубулярных) или пластинчатых (ламеллярных) модулей. К данному списку следует добавить дискоидные модули, которые встречаются у некоторых пурпурных бактерий (примером служит Ectothiorhodospira halochloris). С помощью трубчатых анастомозов везикулы соединяются в гирлянды, а диски — в стопки, напоминающие граны.
Вопреки своему названию, пластинчатые модули реально не существуют, поскольку термином «ламелла» обозначают на срезе фигуры сдвоенных мембран, и неизвестно, на каком протяжении они взаимодействуют друг с другом и как уложены в пространстве.
Способность образовывать ICM, их молекулярный состав и механизм биогенеза кодируются геномом. Однако нет таких генов, которые кодировали бы общую архитектуру ICM, т. е. она обеспечивается эпигенетически. В частности, конфигурация модулей ICM зависит от размера входящих в их состав белковых комплексов, их латерального распределения и физиологического состояния клетки.
Роль первого «формообразующего» фактора видна на примере пурпурных бактерий, у которых геометрия ICM зависит от состава фотосинтетической единицы. В частности, мутант pufХ Rhodobacter capsulatus содержит больше проксимальных антенных комплексов LHCI, чем клетки дикого типа, и, соответственно, образует более крупные везикулярные хроматофоры. Связь между составом и морфологией ICM отмечена для мутантов Rhodobacter sphaeroides с дефектной дистальной антенной LHCII (штамм дикого типа имеет везикулярные, а мутант — тубулярные хроматофоры). Пигмент-белковые комплексы Е. halochloris упакованы в паракристаллические пластинки с гексагональной симметрией, что придает дискоидным хроматофорам характерную уплощенность.
Роль второго фактора можно показать на примере цианобактерий. Сдвоенные мембраны ти- лакоидов могут сближаться или расходиться. Сближению мембран препятствует одноименный (отрицательный) заряд ES-поверхностей. Однако при образовании Pmf, которая у цианобактерий в основном поддерживается благодаря градиенту концентрации протонов (∆рН), он нейтрализуется протонами, поступающими из цитоплазмы. В результате мембраны сближаются, и за счет гидрофобных взаимодействий, сил Ван-дер-Ваальса и водородных связей происходит их адгезия. В состоянии стресса, а также при старении или отмирании клеток адгезия нарушается, мембраны расходятся, и интратилакоидное пространство расширяется с образованием керитомической вакуоли (см. раздел 8.3.11.6). В отличие от тилакоидов цианобактерий, сдвоенные мембраны пурпурных и метилотрофных бактерий находятся на удалении друг от друга. В данном случае Pmf поддерживается за счет разности электрического потенциала (∆ƛ), и поэтому ЕS-поверхности энергизированных мембран заряжены одноименно, что препятствует адгезии.
Пространственная архитектура ГСМ обеспечивается тремя способами.
Во-первых, модули могут соединяться анастомозами (рис. 13, А). Такой вывод вытекает из электронно-микроскопических наблюдений. Например, после мягкого лизиса сферопластов Chromatium vinosum через разрыв клеточной оболочки высвобождается быстро седиментирующая гирлянда везикул (в то время как для осаждения индивидуальных везикул, диаметр которых <50 нм, требуется ультрацентрифугирование). Благодаря анастомозам дискоидные модули пурпурной бактерии Е. halochloris сложены в стопки, а трубчатые модули нитрифицирующей бактерии Nitrosomonas sр. образуют гребневидные кассеты.
Рис. 13. Взаимодействие модулей внутри системы интрацитоплазматических мембран. А — гирлянда; Б — адгезия; В — связь через тилакоидный центр; v1-3 — везикулы; I1— 3 —ламеллы; t1 — 2 — тилакоиды; СМ — цитоплазматическая мембрана.
Во-вторых, модули вплотную сближаются благодаря адгезии PS-поверхностей (рис. 13, Б). Показано, что фактором адгезии у пурпурных бактерий и цианобактерий-прохлорофитов являются апопротеины светособирающего комплекса.
Втретьих, модули могут определенным образом ориентироваться по отношению к клеточной оболочке, что показано на примере цианобактерий, имеющих так называемые «тилакоидные центры» (рис. 13, В). Эти органеллы не только «подвешивают» тилакоиды к СМ, но и участвуют в биогенезе фотосинтетического аппарата (см. ниже).
Функции ICM. Споры о том, являются ли ICM специализированной органеллой или это просто инвагинации СМ, закончились: в пользу первого утверждения свидетельствуют данные, полученные на пурпурных бактериях и цианобактериях с использованием электронной цитохимии, физиологического тестирования интактных клеток и биохимического анализа мембранных препаратов.
Чаще всего роль ICM заключается в создании трансмембранного электрохимического потенциала (Pmf). Ассимилированная энергия используется энергопотребляющими, или эндергоническими системами непосредственно (например, жгутиковым мотором или пермеазами), либо предварительно преобразуется в энергию АТФ. Широкое распространение и большое количество ICM у фототрофных и хемолитотрофных бактерий (см. табл. 8) объясняется высокой энергоемкостью их метаболизма.
Функции ICM изучались в основном у фототрофных бактерий. Прежде всего это касается цианобактерий, с тилакоидами которых связаны фотосинтетические реакционные центры и антенные комплексы, а также электрон-транспортная цепь, создающая Pmf Сведения для хемолитотрофов более фрагментарны; по некоторым данным, ICM содержат дегидрогеназные системы, с помощью которых ассимилируется энергия, освобождающаяся при окислении неорганических субстратов.
В редких случаях ICM принимают участие в конструктивном метаболизме. Прежде всего это касается трансляции. В частности, на поверхности тилакоидов цианобактерий обнаружены круги эпимембранных частиц, напоминающие фреты (англ. fret — орнамент), или агрегаты полирибосом на «шероховатых» тилакоидах пластид. Кроме того, в тилакоидах находится ключевой фермент биосинтеза окрашенных каротиноидов — фитоиндесатураза. Наконец, на них иммобилизованы ферредоксинзависимые редуктазы, играющие ключевую роль в ассимиляции нитрата и нитрита.
Биогенез ICM. Унитарные мембраны растут и дифференцируются путем самосборки, причем в качестве предшественника используются уже существующие мембранные структуры. Такую биогенетическую стратегию можно охарактерзовать, перефразируя основной цитологический закон Рудольфа Вирхова (R. Virchow): «Каждая клетка происходит от другой клетки» (лат. omnis cellula е cellulae). А именно: «Каждая мембрана происходит от другой мембраны» (лат. omnis membrana е membranae). Однако это положение не распространяется на неунитарные мембраны, поскольку их биогенез осуществляется не на матрице, а путем самосборки de novo.
Биогенез унитарной мембраны можно рассматривать как квази-репликативную самосборку органеллы. Новый материал встраивается в мембрану по сродству к резидентным компонентам, благодаря чему воспроизводится ее структурно-функциональная специфичность. В результате мембрана разрастается, и это позволяет ей делиться или отпочковывать дочерние мембранные структуры. У бактерий, как правило, реализуется вариант деления. Исключения составляют отпочкование от ОМ мембранных везикул (см. раздел 8.4.1.3), а также отпочкование от СМ оболочки магнитосомы (см. раздел 8.3.11.6).
Биогенез ICM в основном изучен на примере пурпурной бактерии Rhodobacter sphaeroides. Это координированный процесс биосинтеза, сборки и обновления молекулярных компонентов фотосинтетической органеллы. Его конкретный характер зависит от физико-химических параметров среды, которые могут быть относительно постоянными или изменяются в количественном и качественном отношении. Точнее сказать, сценарии биогенеза ICM зависят от характера роста. Для их классификации можно использовать терминологию, которую предложил немецкий бактериолог Герхарт Древе (G. Drews).
Первый сценарий, «перманентное развитие» (англ, continuous development) представляет собой консервативную репликацию ICM. Она осуществляется при сопряженном росте, когда физико-химические условия внешней среды относительно постоянны. В данном случае уже существующие ICМ служат матрицей, в которую встраиваются «свежие» компоненты фотосинтетического аппарата — реакционные центры, светособирающие антенны и электрон-транспортная цепь. После удвоения мембранного материала он поровну распределяется между дочерними клетками, что обеспечивает постоянство качественного состава и количества ICM.
Второй сценарий, «ступенчатое развитие» (англ. stepwise development) связан с изменением состава и количества ICM. Он реализуется в нестационарных условиях среды, в частности, когда происходит трофический «сдвиг» (англ. nutritional shift), и клетки начинают ассимилировать больше или меньше энергии. В соответствии с данным сценарием бактерии проходят биоэнергетическую акклиматизацию.
При «фотоакклиматизации» (англ. photoacclimation), когда изменяется интенсивность света или его спектральный состав, имеет место характерная поведенческая реакция — приспособление к новому световому режиму на всех уровнях организации фотосинтетического аппарата (от отдельной фотосинтетической единицы до ICM в целом). Если изменяется интенсивность света, действует принцип светолюбивости-теневыносливости (размер антенны находится в обратной зависимости от интенсивности света). Если изменяется спектральный состав света, наблюдается «комплементарная хроматическая адаптация» (англ, complementary chromatic adaptaion), т. е. синтезируются пигменты, поглощающие доминирующий тип световых квантов.
У хемолитотрофных бактерий существует аналогичная стратегия. Состав и количество ICM регулируются так, чтобы оптимально использовать неорганический энергоноситель конкретного типа и определенной концентрации. В отличие от фотоакклиматизации, степень развития ICM тем выше, чем больше концентрация субстрата, хотя корреляция менее заметна.
Ступенчатое развитие по типу «репрессии» (англ. repression) выражается в подавлении пролиферации ICM и/или в уменьшении скорости встраивания специфических компонентов, например, апопротеинов светособирающего комплекса. В свою очередь, «дерепрессия» (англ. derepression) связана с противоположными изменениями и также регулируется на уровне транскрипции. Переход к новому состоянию ICM требует, чтобы репликация мембранного материала была разобщена с репликацией других клеточных структур, в результате чего рост становится несопряженным. При «дерепрессии» в дочерних клетках увеличивается пул ICM, тогда как при «репрессии» избыточный мембранный материал разбавляется по ходу клеточных делений.
Третий сценарий, «ингибирование/индукция» (англ. inhibition/induction) представляет собой образование ICM de novo или их исчезновение. В частности, у пурпурных бактерий биогенез ICM регулируется парциальным давлением кислорода (см. II том учебника). При «ингибировании» биогенез ICM прекращается из-за подавления кислородом транскрипции фотосинтетических генов, объем ICM уменьшается в геометрической прогрессии по ходу деления, и в итоге клетки депигментируются. При частичном ингибировании в составе ICMувеличивается концентрация дыхательных комплексов и уменьшается содержание пигментных систем. В случае «индукции» биогенез фотосинтетического аппарата дерепрессируется в ответ на снижение парциального давления кислорода. Реакционные центры и антенные комплексы равномерно встраиваются в протяженные участки СМ, которые разрастаются и оттесняются в цитоплазму, превращаясь в ICM. Такой механизм называется «разбавлением» (англ, in-synthesis; «синтез внутри»), и он описан для одних видов пурпурных бактерий. У других видов действует механизм «конденсации» (англ. on-synthesis; «синтез на поверхности»), когда точечные локусы СМ служат затравкой для сборки ICM.
Для большинства аноксигенных фототрофных бактерий кислород играет роль негативного регулятора биосинтеза бактериохлорофиллов a — g, что приводит к подавлению биогенеза фотосинтетического аппарата на всех уровнях — от блокирования трансляции апопротеинов и прекращения их встраивания в реакционные центры или антенные комплексы до уменьшения объема ICM.
Однако случается и так, что кислород не влияет на биогенез аноксигенного фотосинтетического аппарата. Иногда (у «регуляторных» мутантов и даже у штаммов дикого типа) биосинтез бактериохлорофилла не подавляется, а, наоборот, индуцируется при переходе от микроаэробных условий к аэробиозу. Такой тип регуляции, в частности, свойствен квази-фототрофным бактериям, которые при нормальном парциальном давлении кислорода образуют больше сферических хроматофоров, чем в микроаэробных условиях. Квази-фототрофных бактерий объединяет то, что они неспособны к анаэробному фототрофному росту. Они обладают портативным фотосинтетическим аппаратом, состав и механизм действия которого такие же, как у пурпурных бактерий. Место квази-фототрофии в энергетическом метаболизме до конца не выяснено. Скорее всего, при дефиците питательных субстратов она служит источником «энергии поддержания» (англ. maintenance energy), необходимой для репарационных процессов и сохранения структурной целостности клетки.
В отличие от факультативно фототрофных пурпурных бактерий или факультативно метилотрофных бактерий, у цианобактерий и облигатно хемолитотрофных бактерий отсутствует механизм ингибирования/индукции, поскольку они ассимилируют энергию только с помощью ICM и постоянно в них нуждаются.
Результаты генетического анализа пурпурных бактерий проливают свет на детали биогенеза ICM, касающиеся поведения интрамембранных комплексов и отдельных молекул.
В данном отношении подробно изучены штаммы Rhodobacter capsulatus. Показано, что латеральное распределение фотосинтетических единиц, или «cis-анизотропия» мембраны и адаптивное изменение состава хроматофоров (фотоакклиматизация) обеспечиваются согласованной транскрипцией двух оперонов. Первый из них, puf-оперон кодирует αβ-апопротеины проксимальной антенны LHCI, а также L/M-полипептиды реакционного центра. Второй, рис-оперон кодирует апопротеины дистальной антенны LHCII (см. II том учебника).
Биогенез ICM комплексно регулируется на уровне биосинтеза пигментов и апопротеинов. В частности, бактериохлорофилл встраивается в ICM на протяжении всего клеточного цикла в стехиометрическом соотношении с антенными полипептидами, в то время как липиды предварительно накапливаются в СМ, а затем, непосредственно перед клеточным делением, доставляются в ICM. В случае Rhodobacter sphaeroides ингибиторы биосинтеза бактериохлорофилла подавляют трансляцию антенных апопротеинов, после чего блокируется образование ICM. Интеграция антенных полипептидов требует специфического распознавания их N-терминальных доменов и зависит от энергизированного состояния ICM.
Механизмы биогенеза ICM у цианобактерий слабо изучены, хотя выяснено, что в доставке пигментов к тилакоидам участвуют тилакоидные центры (англ. thylakoid centres).
Эти цилиндрические образования расположены непосредственно под СМ. Они имеют размер 30 x 300 нм и состоят из глобулярных субъединиц. Тилакоиды расходятся от них лучами. Вероятно, тилакоидный центр не только фиксирует их положение, но и служит каналом, по которому предшественники хлорофилла а поступают из СМ на место своей постоянной дислокации. Доставка галактолипидов в тилакоиды осуществляется при помощи белка VIPP (сокр. англ. vesicle-inducing protein in plastids; 37 кДа), мутации по его гену нарушают способность образовывать ламеллярную систему. Таргетинг белков, местом назначения которых служит полость тилакоида, является эквивалентом секреции и система, обеспечивающая их доставку, сходна с генеральной Sec-системой (см. раздел 8.6.1). Однако эти белки не покидают клетку и функционируют в составе фотосинтетического аппарата. С помощью иммуноцитохимического метода установлено, что с тилакоидами Anabaena sp. и Synechococcus sp. связан шаперон GroEL; специфичные к нему иммунозонды (конъюгаты с коллоидными частицами золота) в основном выявляются на периферических тилакоидах, что свидетельствует об анизотропии процесса сборки фотосинтетических мембран.
Мембранный континуум. Одна из нерешенных проблем цитологии бактерий — это способ структурного взаимодействия между СМ и ICM. В разделе 8.1.1 мы уже говорили о том, что для бактерий характерна топологически непрерывная мембранная система, или «мембранный континуум».
В пользу существования континуума у фототрофных и хемолитотрофных бактерий говорят морфологические данные. В частности, на срезах и криосколах Chromatium sp., Nitrosomonas sp. и Methylocystis sp. часто выявляются анастомозы между СМ и ICM. В клетках пурпурной серной бактерии Ectothiorhodospira halochloris до 90% хроматофоров прямо связаны с СМ. Косвенным свидетельством существования мембранного континуума служат результаты центрифугирования мембранных фрагментов пурпурных бактерий и цианобактерий — помимо зон, соответствующих СМ и ICM, в градиенте образуется зона с промежуточным биохимическим составом.
Однако мембранный континуум не универсален, и пора отказаться от классического постулата Роджера Стэниера: «Бактерии не имеют вакуолей». В некоторых случаях вакуоли обнаружены, и они окружены унитарной мембраной (см. раздел 8.3.11.1).
Сложна ситуация с цианобактериями, где анастомозы между СМ и обширной ICM редко удается обнаружить. С одной стороны, это можно объяснить тем, что их диаметр меньше толщины ультратонкого среза, и поэтому большинство из них не попадает в плоскость электронно-микроскопического изображения. С другой стороны, эволюция ICM у цианобактерий могла привести к их изоляции от СМ, и тогда сохранившиеся анастомозы — рудиментарный признак. Наконец, ICM цианобактерий могли специализироваться настолько, что приобрели полную автономию от СМ.
Наследуют ли континуум дочерние клетки, или после деления он восстанавливается путем вторичного слияния СМ и ICM? Может ли континуум нарушаться в интервале между делениями?
На эти вопросы ответ еще не получен. Однако ясно, что мембранная система бактерий потенциально способна к формированию континуума, и этимони отличаются от эукариотов, у которых прямое взаимодействие между СМ и ICM совершенно исключено.
С учетом изложенного можно говорить о трех типах взаимоотношений между СМ и ICM:
1. Открытый континуум. Анастомозы между СМ и ICM существуют, и релаксированные цистерны ICM сообщаются с периплазматическим компартментом. В таком случае они содержат периплазму (рис. 14, А).
Рис. 14. Структурные взаимоотношения цитоплазматической мембраны и интрацитоплазматических мембран. А — «открытый» континуум; Б — «закрытый» континуум; В — «разорванный» континуум; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; IСМ — интрацитоплазматические мембраны.
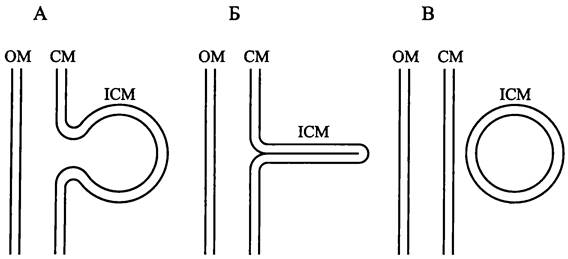
2. Закрытый континуум. Анастомозы между СМ и ЮМ существуют, однако цистерны ICМ стянуты или сплющены и поэтому не сообщаются с периплазматическим компартментом (рис. 14, Б).
3. Разорванный континуум. Между СМ и ICМ не существует анастомозов, цистерны ICМ не сообщаются с периплазматическим компартментом и образуют непрерывный эндоплазматический компартмент или систему изолированных компартментов, аналогичную вакуому эукариотной клетки. В таком случае внутреннее пространство этих цистерн заполняется «эндоплазмой» (рис. 14, В).
8.3.6. Хромосомы
Мембранная система прокариотной клетки устроена таким образом, что она не изолирует генетический материал от цитоплазмы. Хромосомы у бактерий и архей могут занимать различное положение и взаимодействуют с другими органеллами (оболочкой, рибосомами, цитоскелетом и включениями).
В данном разделе мы остановимся только на морфологии и молекулярном строении прокариотных хромосом. Поведение хромосом в онтогенезе (репликация, расхождение), инфраструктура прокариотного генома и другие вопросы цитогенетики прокариотов будут рассмотрены в III томе учебника.
8.3.6 1. Хромосомы бактерий
Большинство работ по бактериальным хромосомам выполнено на Е. coli, В. subtilis и Caulobacter crescentus. Результаты свидетельствуют о большом сходстве, которое проявляют эти виды в отношении пространственной организации и динамического поведения хромосом.
История изучения компартментализации генетического материала у бактерий: от «ядра» до свободной хромосомы. Проблема существования ядра у бактерий изучается с конца XIX в.
В 1890 г. зоолог Отто Бючли обнаружил у серных бактерий прижизненно окрашиваемое гематоксилином «центральное тело», которое он принял за истинное ядро. В 1891 г. его ассистент Варлих (Wahrlich) описал у зафиксированных бактерий «хроматин», окрашивающийся ферроцианидом калия. В 1900 г. Фейнберг (Feinberg) применил к бактериям эозиновую методику, предложенную Романовским (D. Romanowski) и Гимза (G.Giemsa) для выявления ядра у малярийного плазмодия. При этом он выявил у них некий аналог ядра, или «ядерное образование» (нем. Kerngebilde). В 1932 г. Причманн (К. Pritschmann) и Риппель (A. Rippel) выявили ДНК в клетках В. mycoides. Для этого они использовали цитохимический метод Фельгена (R. Feulgen), основанный на «кислотной депуринизации», или удалении с помощью кислоты пуриновых оснований из дезоксирибонуклеотидов и проведении цветной реакции между освобожденными альдегидными группами и анилиновым реактивом Шиффа. В 1940-х годах Карл Робиноу провел анализ бактериального хроматина in situ и пришел к ошибочному выводу о его сходстве с хромосомами эукариотов (в частности, он использовал такие термины, как «ядерная структура», «митоз», «телофаза» и т. д.). Все упомянутые исследователи считали, что бактерии имеют ядро, окруженное мембраной.
В начале 1950-х годов, когда при биологических исследованиях стали использовать электронный микроскоп и была разработана техника приготовления ультратонких срезов, Чэпмен и Хиллиэр (J. Hillier) показали, что у бактерий ДНК-содержащая зона не окружена мембраной. Благодаря этому уже в 1958 г. Эдуард Келленбергер сформулировал вывод о том, что бактерии не имеют ядра и митоза, а их «нуклеоид» представляет собой одну или несколько хромосом, свободно лежащих в цитоплазме.
С помощью радиоавтографического метода Кэрнс (J. Cairns) в 1963 г. доказал, что хромосома Е. coli представляет собой кольцевую молекулу ДНК с контурной длиной ~1 мкм, что соответствует молекулярной массе 3106 кДа (4600 т. п. н.).
Бактериальные хромосомы обычно представляют собой компактные тела, или «нуклеоиды» расположенные в центре клетки и ориентированные вдоль ее продольной оси. Единственное исключение составляет гигантская бактерия Е. fishelsoni (см. раздел 8.1.3.4), хроматин которой образует узкий ободок по периферии клетки.
При окрашивании живых клеток эпифлуоресцентными красителями (например, диамидинофенилиндолом; diamidine phenylindole, DAPI), а также при использовании метода замораживания- протравливания хромосомы напоминают по форме коралловую веточку. После химической фиксации они приобретают зернистый вид или состоят из электроноплотных фибрил, пучки которых часто имеют волнистую или перистую структуру (наблюдаемые картины могут быть артефактом, поскольку хромосома постоянно меняет свою конфигурацию в ходе репликации и транскрипции).
Бактериальную хромосому можно выделить в препаративном виде как частицу, называемую «компактной» хромосомой (англ/ folded chromosome). Ее диаметр составляет 0,5-1,5 мкм, что соответствует размеру нуклеоида in situ, и она имеет константу седиментации 1500-3000 S. Чтобы избежать декомпактизации хромосомы при лизисе клетки, используются химические реагенты, экранирующие силы электростатического отталкивания между фосфатными группами ДНК, в частности 1М NaCl или 5 мМ спермидин. В настоящее время установлено, что в таких условиях изменяются архитектура хромосомы и характер ее взаимодействия с цитоплазматическими структурами и клеточной оболочкой, т. е компактная хромосома — это препаративный артефакт.
Анализ состава хромосом показал, что прокариотный хроматин беднее белком, чем хроматин эукариотов (соответственно <10% и 50%), и поэтому он был назван «LP-хроматином» (от англ. low- protein). Хотя такие препараты обычно содержат РНК и фрагменты клеточной оболочки, это не структурные части хромосомы, а сопутствующие примеси. Истинными белковыми компонентами нереплицирующейся бактериальной хромосомы являются ДНК-связывающие белки (см. ниже), и в динамическом взаимодействии с ней находятся РНК-полимераза, а также топоизомеразы, т. е. ферменты, влияющие на ее компактизацию.
Общие сведения о бактериальных хромосомах. В большинстве случаев бактериальная хромосома имеет контурную длину ~1 мм, а это на три порядка больше диаметра клетки. Для того, чтобы разместить в цитоплазматическом компартменте такую макромолекулу, ей нужно придать компактную форму, что достигается с помощью специальных структурных белков и ферментов и требует затраты энергии (см. ниже).
Бактериальная хромосома обычно замкнута в кольцо, и две репликативные вилки движутся в противоположных направлениях от участка инициации (oriC), встречаясь в локусе терминации репликации (Тег), где находится сайт dif, отвечающий за декатенацию, т. е. размыкание сцепленных дочерних хромосом. Хотя способ инициации репликации и ее механизм одинаковы у всех трех филогенетических доменов, эукариотные хромосомы, в отличие от бактериальных, реплицируются не из одного, а из нескольких участков инициации.
Многие бактерии содержат внехромосомные генетические элементы — плазмиды, которые отличаются от хромосом не столько размером (мегаплазмиды приближаются по величине к небольшим хромосомам), сколько тем, что они не содержат информационных генов. Поэтому при их элиминации клетка сохраняет жизнеспособность.
Число хромосом в бактериальной клетке зависит и от природы организма, и от фазы развития популяции. Многокопийные хромосомы встречаются у представителей разных родов. Например, Deinococcus radiodurans (фила BIV «Deinococcus- Thermus») имеет от 4 до 10 копий, Borrelia hermsii (фила BXVII Spirochaetes) — от 8 до 16, Desulfovibrio gigas (класс «Deltaproteobacteria») — от 9 до 17, a Azotobacter vinelandii (класс «Gammaproteobacteria») — до 80 копий хромосомы на клетку.
В молодых клетках хромосом обычно больше, чем в старых (например, Е. coli в стационарной фазе содержит 1-2 хромосомы, а в экспоненциальной фазе 2-8 хромосом).
Кроме того, установлено, что бактериальные клетки могут обладать не только многокопийными хромосомами, но и в некоторых случаях сложным «кариотипом», когда геном распределяется между несколькими хромосомами и внехромосомными элементами.
Например, геномы Agrobacterium tumefaciens С58, Brucella melitensis и Rhodobacter sphaeroides (класс «Alphaproteobacteria») содержат по две разные хромосомы. В состав геномов В. cereus А0836 76 (фила ВХIII Firmicutes), Leptospira interrogans (фила BXVII Spirochaetes) и Rhizobium meliloti (класс «Alphaproteobacteria»), помимо хромосом входят 1-2 мегаплазмиды размером 100-500 т. п. н.
Наконец, в последнее время опровергнуто классическое представление о том, что бактериальная хромосома обязательно имеет кольцевую форму (в отличие от линейных хромосом у эукариотов). С помощью метода гельэлектрофореза в пульсирующем поле (англ. pulced-field gel electrophoresuis, PFGE), позволяющего разделять большие и маленькие линейные молекулы ДНК, было установлено, что некоторые бактерии содержат и кольцевые, и линейные хромосомы, а также могут иметь только линейные хромосомы.
Первая линейная хромосома размером 946 т. п. н. была выявлена в 1989 г. у возбудителя болезни Лайма — Borrelia burgdorferi В31 (фила BXVII Spirochaetes). Затем линейные хромосомы обнаружили у А. tumefaciens (класс «Alphaproteobacteria»), Rhodococcus fascians (фила ВХIII Firmicutes), Streptomyces coelicolor и S. lividans (фила BXIV Actinobacteria).
Линейные плазмиды были впервые найдены в 1977 г. у S. rochei. Потом их обнаружили у многих других стрептомицетов, а также у R. fascians, Nocardia ораса (фила BXIV Actinobacteria), многих видов Borrelia sp. и Thiobacillus versutus (класс «Betaproteobacteria»).
В связи с открытием у бактерий линейных хромосом и линейных плазмид встал вопрос о природе их теломеров (концевых участков ДНК, обеспечивающих стабильность и полную репликацию линейного репликона), а также о механизме репликации, поскольку ДНК-полимеразы не способны копировать ДНК с «открытого» конца. Как выяснилось, проблема решается одним из трех способов — с помощью одноцепочечных концевых шпилек ДНК, с помощью белка, ковалентно присоединенного к 5'-концу двухспирального теломера, а также благодаря временному переходу линейной хромосомы в кольцевую репликативную форму.
Участок инициации репликации (oriC) занимает центральное положение в линейной хромосоме и разделяет ее на две примерно равные по длине «реплихоры».
ДНК-связывающие белки. Бактериальная хромосома занимает до 20% цитоплазматического компартмента и имеет объем около 0,2 мкм3. Концентрация ДНК в этом участке клетки чрезвычайно велика и близка к концентрации ДНК в хроматине интерфазных ядер (20-50 мг • мл-1). При растворении такого количества ДНК в пробирке получился бы очень вязкий гель.
Чтобы упаковаться в крошечном объеме, не теряя способности к репликации, репарации, рекомбинации и транскрипции, прокариотная хромосома должна сочетать в себе противоречивые свойства — высокую компактность и динамичную структурированность. Объемный коэффициент упаковки ДНК в бактериальной клетке исключительно высок и достигает х 1 тыс. (хотя это уступает плотности упаковки ДНК в головке бактериофага, которая в 30-60 раз выше).
В эукариотных хромосомах существуют несколько уровней компактизации. Нижний из них представлен нуклеосомой — регулярно повторяющимся октамерным комплексом: по две копии гистонов Н2а, H2b, НЗ и Н4. При сборке нуклеосомы два димера гистонов НЗ/Н4 образуют тетрамер, или «тетрасому», с которой связывается ДНК. К противоположным сторонам тетрасомы присоединяются два димера гистонов Н2а/Н2b. Вокруг получившегося октамера суперспирализуется участок ДНК протяженностью 146 п. н., образуя 1,65 «отрицательных», т. е. левозакрученных, супервитка (напомним, что закручивание одной нити ДНК вокруг другой называется витком двойной спирали, а закручивание двойной спирали вокруг себя — суперспирализацией). Помимо гистонов Н2а, Н2b, НЗ и Н4 существуют не гомологичные им «линкерные» гистоны Н1/Н5, которые присоединяются к тетрасоме, обеспечивая суперспирализацию еще 20 п. н., и вместе они образуют «хроматосому».
Гистоны относятся к катионным, или положительно заряженным белкам, и их взаимодействие с ДНК неспецифично.
Нуклеосомная организация хроматина стабилизирует ДНК, одновременно препятствуя транскрипции. Для «разрыхления» хроматина N-концевые участки молекул гистонов, особенно линкеров Н1/Н5 подвергаются посттрансляционной ковалентной модификации путем ацетилирования или метилирования по остаткам Lys.
В бактериальной хромосоме отсутствует нижний уровень упаковки в виде стабильной, регулярно повторяющейся нуклеосомы, и поэтому определить участие белков в ее строении достаточно трудно. Кроме того, оказалось, что некоторые из них играют не только структурную, но и регуляторную роль.
Компактизация бактериальной хромосомы зависит от небольшой гетерогенной группы белков, важнейшими из которых являются HU, H-NS и SMC. Гистоноподобные белки HU и H-NS, или HLP-белки (от англ. histone-like protein) синтезируются в наибольшем числе копий. Их взаимодействие с ДНК зависит от ее первичной последовательности, а также от присутствия в ней шпилек и изгибов.
HU и H-NS, а также минорные гистоноподобные белки FIS и IHF не только участвуют в организации бактериальной хромосомы, подобно эукариотным гистонам, но и влияют на экспрессию генов, а также на репликацию и рекомбинацию ДНК.
Белок HU. Этот гистоноподобный белок (от англ. histone U; «гистоноподобный белок штамма U-93 Е. coli») представлен гетеродимером субъединиц Huα и Huβ и образуется в количестве 3 • 104 копий на клетку. Субъединицы белка HU имеют молекулярную массу ~10 кДа и обладают катионными свойствами, благодаря чему они образуют солевые мостики с фосфатными группами ДНК.
Белок HU выполняет функцию, сходную с функцией гистонов, хотя гомология между ними отсутствует. Совместно с ДНК-гиразой (топоизомеразой I) он обеспечивает отрицательную суперспирализацию ДНК. Предполагается, что 10 димеров HUαβ образуют изгиб на участке ДНК длиной 102 п. н. (рис. 15, А).
Рис. 15. Три уровня компактизации бактериальной хромосомы. А — нижний (изгиб с помощью 10 димеров белка НU); Б — средний (суперспирализация с помощью димера белка SМС и вспомогательных белков); В — верхний (образование доменов).
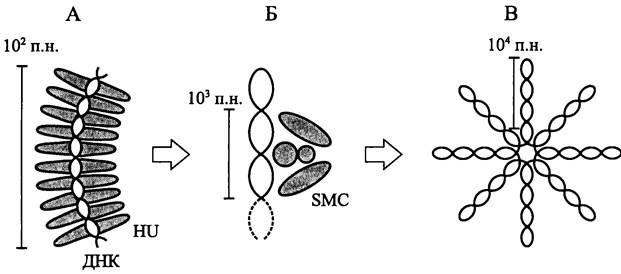
В отличие от гистонов, белок НU оставляет ДНК более доступной для транскрипции, и поэтому нет необходимости в его посттрансляционной модификации путем ацетилирования или метилирования. Регуляция транскрипции осуществляется в результате сайт-специфического связывания положительных и отрицательных регуляторов, а также путем смены σ-факторов.
Помимо роли, которую белок HU играет в общей упаковке ДНК, он влияет на ее локальную конфигурацию. Благодаря этому он косвенно участвует в таких процессах, как репликация, сайт-специфическая рекомбинация и транскрипция. Такую же роль выполняют небольшие катионные белки FIS (от англ. factor of internai stabilization) и IHF (от англ. intégration host factor).
Белок H-NS. Этот гистоноподобный белок (от англ. histone-like nucleoid structuring protein) образуется в количестве 2 • 104 копий на клетку. Он имеет молекулярную массу 15,6 кДа и не несет на себе избыточного электростатического заряда. Его С-концевой домен взаимодействует с ДНК, а N-концевой домен отвечает за белок-белковые взаимодействия и гомоолигомеризацию. Этот белок компактизует ДНК, изменяет ее суперспирализацию и индуцирует ее изгибы. Однако его основной ролью является не упаковка ДНК, а участие в глобальной регуляции, поскольку под его контролем находятся до 40 генов (ген антисмысловой РНК micF, ген порина оmрС, ген β-глюкозидазы bgl и т. д.).
Белки из семейства SMC. Эти белки (от англ. structural maintenance of chromosomes) играют универсальную роль в организации хромосом у представителей всех трех филогенетических доменов. Они обнаруживают гомологию с миозином и у эукариотов играют роль моторов при конденсации хроматина.
SMC — это крупные (> 50 кДа) белки, образующие V-образные гомо- или гетеродимеры. Плечи гетеродимера состоят из взаимно перекрученных антипараллельных цепей длиной ~50 нм и вместе со вспомогательными белками создают повторяющуюся стабильную структуру в виде отрицательных супервитков размером 103 п. н. (рис. 15, Б).
В зависимости от специфики вспомогательных белков, димеры SMC могут играть не только роль «конденсинов», т. е. суперспирализовать бактериальную ДНК, но и участвовать в ее репарации, рекомбинации, сегрегации дочерних хромосом и других процессах.
Уровни компактизации хромосомы. Таким образом, нижний уровень компактизации бактериальной ДНК обеспечивается гистоноподобными белками HU и H-NS. Хотя образующиеся при этом комплексы отдаленно напоминают нуклеосомы, они не имеют такой же стабильной структуры и, кроме того, не повторяются регулярно в cis-положении (т. е. вдоль нити ДНК).
Однако главная роль в компактизации бактериальной ДНК принадлежит не гистонам и не гистоноподобным белкам (в отличие от эукариотов, а также от многих архей), а отрицательной суперспирализации, которая осуществляется с помощью SMC-белков и топоизомераз.
Ферменты из суперсемейства топоизомераз локально действуют на одну или две взаимодействующие молекулы ДНК. Они создают или релаксируют торсионное напряжение, образуют или завязывают узлы, а также вызывают (де)катенацию, т. е. размыкают или, наоборот, переплетают кольца ДНК.
Топоизомеразы I производят одноцепочечные, а топоизомеразы II — двухцепочечные разрывы в молекуле ДНК. Типичным примером топоизомеразы I является ДНК-гираза, обеспечивающая суперспирализацию прокариотных хромосом.
Суперспирализованная молекула обладает избыточной свободной энергией, которая служит движущей силой для многих процессов, происходящих на уровне хромосомы. Именно поэтому полная потеря отрицательной суперспирализации вызывает летальный эффект.
В незащищенной молекуле, как в скрученной пружине, для релаксации достаточно единственного одноцепочечного разрыва. Однако in situ такой эффект предотвращается благодаря сегрегации ДНК на независимо суперспирализованные петли, или хромосомные «домены». В случае разрыва они релаксируются поодиночке, и суперспиральная структура хромосомы в целом не нарушается.
Число и размер этих доменов точно не установлены. Считается, что бактериальная хромосома состоит из 50-200 доменов, и их средний размер, по последним данным, составляет ~104 п. н. (рис. 15, В). Однако неясно, где находятся внутриге- номные границы этих доменов, а также — способны ли они перемещаться.
8.3.6.2. Хромосомы архей
Хромосомы архей имеют признаки, свойственные, с одной стороны, бактериям, с другой стороны — эукариотам.
Как и у бактерий, компактные нуклеоиды у архей расположены в центре клетки и ориентированы вдоль ее продольной оси. При окрашивании живых клеток эпифлуоресцентными красителями, а также на криосколах хромосомы имеют форму коралловой веточки, а после химической фиксации приобретают зернистое или фибриллярное строение.
Как и у бактерий, число хромосом у архей зависит от природы организма и фазы развития популяции. Например, Sulfolobus acidocaldarius в экспоненциальной фазе содержит 1-2 хромосомы, а М. voltae 3-10 хромосом. В молодых клетках обычно больше хромосом — в частности, Methanococcus jannashii в стационарной фазе имеет 1-5 хромосом, а в экспоненциальной фазе 3-15 хромосом.
Как и бактерии, архей могут содержать плазмиды. Например, у экстремального галофила Halofetrax mediterránea обнаружены отрицательно суперспирализованные кольцевые мегаплазмиды размером >500 т.п.н.
Вопрос о присутствии у архей набора разных хромосом, а также о существовании у них линейных хромосом и линейных плазмид остается открытым.
Как и в случае бактерий, хромосомы у архей имеют кольцевую форму. Число участков инициации репликации (oriC) окончательно не установлено, хотя получены данные о том, что их несколько.
ДНК-связывающие белки: гистоны. В начале 1990-х годов у термофильного метаногенного археота Methanothermus férvidas были обнаружены небольшие (~7 кДа) катионные белки HMfA и HMfB. Оказалось, что они на 30% гомологичны эукариотным гистонам и также обеспечивают нижний уровень компактизации ДНК. Истинные гистоны есть и у других членов филы АII Euryarchaeota (например, гистон HMk Methanopyrus kandleri, имеющий молекулярную массу 7 кДа), однако они отсутствуют у представителей филы AI Crenarchaeota.
Археотные гистоны по размеру меньше эукариотных и содержат укороченный N- концевой участок, подвергающийся посттрансляционной модификации, и поэтому характер их взаимодействия с ДНК несколько иной.
Нуклеосомы архей не обладают октамерной структурой, как у эукариотов, поскольку гомологи гистонов Н2а/Н2b у них отсутствуют. Однако они обнаруживают большое сходство с тетрасомой (H3/H4)2. Вокруг тетрамера суперспирализуется участок ДНК протяженностью ~70 п. н., образуя один правозакрученный (положительный) виток. Предполагаются, что гомологичные нуклеосомы архей и эукариотов в ходе эволюции дивергировали от тетрамерного прототипа.
Негистоновые ДНК-связывающие белки. Термофильные и гипертермофильные представители филы AI Crenarchaeota (в частности, Sulfolobus solfatarius
и S. tokodaii) не содержат гистонов, однако у них есть другие катионные ДНК- связывающие белки.
Первый из них, Sul7d (от англ. Sulfolobus 7-kDa DNА-binding protein), укладывается вдоль малой борозды ДНК, защищая хроматин от температурной денатурации. Кроме того, он компактизирует положительно суперепирал изованную ДНК (см. ниже). Показано, что он метилируется по остаткам Lys, хотя значение этой посттрансляционной модификации еще не выяснено.
Второй ДНК-связывающий белок, Alba (от англ. acetylation lowers binding affinity) имеет молекулярную массу 10 кДа и встречается не только у разных видов рода Sulfolobus, но и у представителей филы АII Euryarchaeota, а также у эукариотов, например, трипаносомид, высших растений и даже некоторых позвоночных. У архей он составляет до 5% растворимого белка и образует димеры, полностью укрывающие ДНК, что придает ей устойчивость к нуклеазам. Однако в одиночку он не обеспечивает нижнего уровня компактизации; требуется второй катионный белок — Sul7d или гомолог бактериального белка HU. Подобно гистонам, белок Alba подавляет транскрипцию, что зависит от его посттрансляционной модификации (ацетилаза еще не идентифицирована, а роль деацетилазы выполняет гомолог «гистоновой» деацетилазы Sir2).
Третий ДНК-связывающий белок описан у термофильного археота Thermoplasma acidophilum (фила АII Euryarchaeota). Он гомологичен бактериальному белку HU, компактизирует ДНК и защищает ее от тепловой денатурации, повышая температуру ее плавления примерно на 40°С.
Более высокие уровни компактизации ДНК, по-видимому, обеспечиваются SMC- белками. Их гомолог обнаружен, в частности, у метаногена М. jannaschii.
«Обратная» гираза. Как уже отмечалось, ДНК эукариотов и бактерий отрицательно суперспирализована. У первых это достигается главным образом путем закручивания ДНК на нуклеосомном октамере, у вторых — в основном с помощью ДНК-гиразы.
Уникальным архитектурным свойством хромосом архей является положительная (правозакрученная) суперспирализация ДНК, которая осуществляется при помощи «обратной» гиразы. Предполагается, что положительная суперспирализация стабилизирует ДНК у экстремофильных архей, особенно у таких гипертермофилов, как Sulfolobus.
8.3.7. Квази-эукариотизм
В настоящем разделе мы рассмотрим «квази-эукариотизм» — очень редкое явление, когда у прокариотов хромосомы компартментализуются при помощи 1СМ.
Компартментализация «информационных» структур и процессов у прокариотов. Как мы уже отмечали, эукариоты принципиально отличаются от прокариотов тем, что у них сайты процессинга генетического материала изолированы ядерной оболочкой от сайтов его экспрессии. Кроме того, в эукариотной клетке трансляция не начнется до тех пор, пока не завершится раунд репликации ДНК, а также пока не закончится синтез и процессинг мРНК (см. раздел 5.1).
В прокариотной клетке процессы репликации, трансляции и транскрипции менее изолированы — как в пространстве, так и во времени. Транскрипция мРНК может начаться еще до завершения полного раунда репликации ДНК. В свою очередь, трансляция может инициироваться на частично синтезированной молекуле мРНК, а транслирующие рибосомы могут регулировать действие РНК-полимеразы (см. раздел 8.3.8).
Однако это не означает, что у прокариотов транскрипция и трансляция не обособляются в пространстве. На рубеже XX-XXI вв. Эррингтон (J. Errington), а следом за ним и другие исследователи сконструировали молекулярные гибриды (англ. fusion protein) между зеленым флуоресцирующим белком и белковыми субъединицами РНК-полимеразы В. subtilis, а также между зеленым флуоресцирующим белком и белковыми субъединицами рибосомы. В результате с помощью светового микроскопирования живых клеток было показано, что РНК-полимераза находится в центральной части клетки, тогда как рибосомные белки расположены на периферии. Безусловно, из этого еще не следует, что у бактерий, как и у эукариотов, транскрипция и трансляция полностью изолируются друг от друга, поскольку эти процессы могут «перекрываться» на границе между флуоресцирующими зонами.
Аналогичным способом показано, что репликативный комплекс В. subtilis не рассредоточен по клетке, а расположен в ее центре, хотя в отсутствии ядерной оболочки и нуклеоплазмы хромосомы не компартментализованы, а лежат непосредственно в цитоплазме.
Еще раз напомним, что прокариотная клетка по определению не содержит ядра, маркером которого на ультраструктурном уровне служит ядерная оболочка. Строго говоря, по отношению к генофору прокариотов не следовало бы употреблять термины, намекающие на сходство с ядром, в частности, такой широко распространенный термин, как «нуклеоид» (см. разделы 2.3 и 2.4).
Квази-эукариотизм у планктомицетов. Планктомицеты (фила BXV Planctomycetes) являются одной из важнейших, в плане биоразнообразия и экологии, групп прокариотов. Особого внимания заслуживают две цитологические особенности этих бактерий. Первая — это отсутствие у них муреинового саккулуса (по аналогии с филой ВVII Thermomicrobia и микоплазмами из филы BXIII Firmicutes). Вторая — это квази-эукариотизм.
В 1995-2001 гг. австралийский микробиолог Ферст (J. A. Fuerst) обнаружил, что у пресноводного планктомицета Gemmata obscuriglobus ДНК окаймлена структурой, морфологически сходной с ядерной оболочкой. Она состоит из двух параллельных мембран толщиной 5-8 нм, разделенных промежутком в 14-21 нм, и изолирует квази-нуклеоплазму с расположенными в ней фибриллами ДНК от цитоплазмы. Однако если внимательно проанализировать опубликованные микрофотографии, выясняется, что квази-ядерная оболочка представляет собой бокаловидную инвагинацию СМ (рис. 16, А). Хотя такая органелла была названа «ядерным телом» (англ. nuclear body), это аналог, а не гомолог ядра. При внешнем сходстве с ядром мембранная граница ядерного тела не имеет характерных признаков ядерной оболочки и не изолирует процессы транскрипции и трансляции (поскольку в «квази-нуклеоплазме» содержатся рибосомы). Кроме того, истинная ядерная оболочка сращена с ICM, но никогда не образует анастомозов с СМ (см. раздел 5.1).
Рис. 16. Квази-эукариотизм. А — у планктомицетов; Б — у цианобактерий; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; ICM —интрацитоплазматические мембраны; М — муреиновый саккулус; S — поверхностный белковый слой; Т — тилакоид; МТ — модифицированный тилакоид.
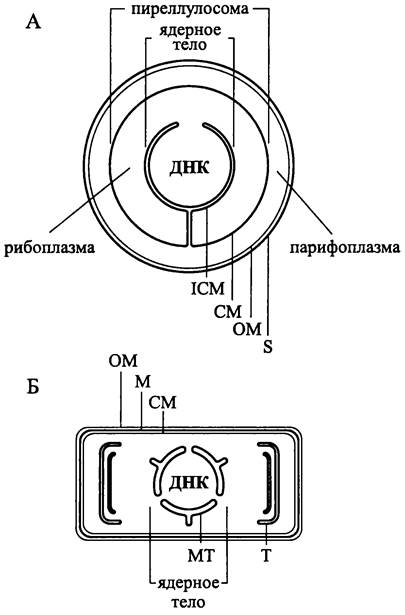
G. obscuriglobus и другие планктомицеты (в частности, Candidatus «Brocadia anammoxidans», Isosphaera pallida, Pirellula marina и P. staleyi) характеризуются еще одной нетривиальной особенностью — у них сильно гипертрофирован периплазмати- ческий компартмент. Его ошибочно принимают его за свободную от рибосом цитоплазму и предлагают для ее обозначения термин «парифоплазма» (греч. pariphe — кайма). В свою очередь, цитоплазму планктомицетов называют «рибоплазмой» (англ. ribosome — рибосома), а совокупность ядерного тела и рибоплазмы— «пиреллулосомой» (от родового имени планктомицета Pirellula).
Функциональное значение изоляции хромосомы G. obscuriglobus внутри напоминающей ядерную оболочку бокаловидной инвагинации СМ остается неясным.
Квази-эукариотизм у цианобактерий. Второй пример квази-эукариотизма связан с цианобактериями. В конце 1980-х годов американский бактериолог Йенсен (Т. Е. Jensen) опубликовал электронные микрофотографии Oscillatoria sp., на которых ДНК окружена двойной мембранной оболочкой с «ядерными порами» и соединенными с ней «цистернами эндоплазматического ретикулума» (рис. 16, Б). По его мнению, это показывает, что в ходе эволюции ядерная оболочка образовалась в результате видоизменения тилакоидов. Однако он не учел, что эукариоты произошли не от бактерий — это разные филогенетические домены. Кроме того, он анализировал старую (145-дневную) культуру, где ламеллярная система претерпела дегенеративные изменения, и ее морфология случайно напоминает ядерную оболочку.
8.3.8. Рибосомы
У прокариотов генетический материал не изолирован от аппарата трансляции, и прокариотные рибосомы занимают почти весь цитоплазматический компартмент. Относительное (по сравнению с другими органеллами) количество рибосом у прокариотов выше, чем у эукариотов, и это обеспечивает более высокую активность их метаболизма, а также более высокую скорость их роста и размножения.
Рибосомы — это множественные генеральные микрокомпартменты, которые находятся в цитоплазматическом компартменте и выполняют роль универсальной белоксинтезирующей органеллы. За исключением редких случаев, когда полипептиды синтезируются нерибосомным путем (см. разделы 8.3.11.4, 8.4.3.1 и 8.5.3.2), аминокислоты связываются в линейную цепь только благодаря ферментативной активности рибосом.
Биосинтетический процесс трансляции уникален, поскольку информация о порядке расположения нуклеотидных триплетов (код мРНК) переводится в информацию о порядке расположения аминокислот (код полипептида). Посредником между этими кодами, или «адаптером» (англ. adapter — звукосниматель аудиосистемы) является тРНК. Она доставляет аминокислоту в пептидилтрансферазный центр и одновременно с этим распознает ее кодон в молекуле мРНК.
Рибосомы служат универсальным аппаратом для синтеза белка, без которого невозможно существование клеточных форм жизни. Собственным аппаратом трансляции обладают не только облигатные внутриклеточные паразиты, но и полуавтономные органеллы бактериального происхождения — митохондрии и пластиды. Центральную роль в организации рибосомы играют рибосомные РНК, нуклеотидная последовательность которых содержит и высоко консервативные, и вариабельные участки. С учетом этого именно структура рРНК была избрана в качестве универсального критерия для реконструкции филогении клеточных форм жизни (см. главы 2, 4 и 7).
Типы рибосом. Рибосома — это мультимолекулярный комплекс, состоящий из рРНК и рибосомных белков в массовом соотношении 2:1.
В рабочем состоянии рибосома, или «моносома» представляет собой частицу диаметром 25 нм, которая состоит из двух субъединиц — большой L-субъединицы (от англ, large) и малой S-субъединицы (от англ, small). Они имеют разный состав, разную морфологию и выполняют разные функции.
По количественным признакам все рибосомы подразделяются на два типа — прокариотный и эукариотный. Прокариотная рибосома имеет коэффициент седиментации 70S(субъединицы 50S и 30S), а эукариотная рибосома — 80S (субъединицы 60S и 40S). Прокариотная рибосома содержит три молекулы рРНК — 23S (~3000 нуклеотидов), 16S (~1500 нуклеотидов) и 5S (~120 нуклеотидов), а также 53-65 однокопийных белков. Эукариотная рибосома устроена сложнее прокариотной. Она содержит не три, а четыре молекулы рРНК — 28S (4000-6000 нуклеотидов), 18S (1750-1850 нуклеотидов), 5S (~120 нуклеотидов) и 5,8S (~150 нуклеотидов), а также более богатый набор однокопийных белков (70-84).
Механизм трансляции. Хотя современные представления об архитектуре рибосом и процессе трансляции сложились на основе данных, полученных на бактериях, доказано, что работа рибосомы универсальна у всех трех глобальных доменов (рис. 17).
Рис. 17. Рабочий цикл рибосомы. А — исходное состояние; Б — претранслокационное состояние; В — гибридное транслокационное состояние; Г — однородное транслокационное состояние; Р, А и Е — сайты связывания пептидил-тРНК, аминоацил-тРНК и деацилированной тРНК; ЕF-Тu и ЕF-G — факторы элонгации.

Результаты рентген-структурного анализа с уровнем разрешения 5,5Ä и криоэлектронной микроскопии дали для рибосомы Е. coli картину геометрического тела сложнейшей конфигурации, состоящего из взаимно переплетенных молекул рРНК и белка. Внутри него, а также на его поверхности имеются каналы, борозды, углубления, площадки, выступы и мостики.
Субстратами для биосинтеза полипептидной цепи служат аминоацил-тРНК, причем для каждой аминокислоты существует своя тРНК и своя аминоацил-тРНК-синтетаза. Специфические тРНК (~75 нуклеотидов) различаются по первичной структуре, однако все они имеют стандартную Г-образную третичную структуру. На дистальном конце длинного «локтя» находится антикодон, комплементарный триплету мРНК, который кодирует специфическую аминокислоту. На дистальном конце короткого «локтя» всех тРНК находится 3'-концевая последовательность ССА. К аденозину (по его 2'- или З'-гидроксильному радикалу) присоединяется а-карбоксильная группа специфической аминокислоты. В ходе трансляции антикодон длинного «локтя» распознает на S-субъединице кодон мРНК, а короткий «локоть» с аминокислотой взаимодействует на L-субъединице с пептидилтрансферазным центром, который катализирует образование пептидной связи.
Долгое время считали, что трансляцию обеспечивают рибосомные белки, а рРНК служит лишь каркасом для их сборки. Однако в настоящее время доказано, что роль главного катализатора трансляции играет рРНК, а белки выполняют структурную функцию.
Между субъединицами рибосомы существует разделение труда. Малая субъединица содержит декодирующий центр, который обеспечивает взаимодействие между мРНК и тРНК. Большая субъединица содержит пептидилтрансферазный центр. В организации декодирующего центра участвуют 16S рРНК и рибосомные белки, в то время как пептидилтрансферазный центр образован только 23S рРНК. Последовательность Шайна-Дальгарно (J. Shine, L. Dalgarno), предшествующая стартовому кодону мРНК, спаривается с комплементарной последовательностью на З'-конце 16S рРНК. Антикодоновый конец тРНК также взаимодействует с 16S рРНК, тогда как акцепторный конец тРНК взаимодействует с 23S рРНК.
Рибосома шаг за шагом образует пептидные связи в направлении от N-конца к С-концу. Для инициации полипептидной цепи у бактерий используется особая аминокислота — формилметионин, которая доставляется в рибосому с помощью специфической тРНКРmet. Происходит спонтанная пептидилтрансферазная реакция: нуклеофильная α-аминогруппа аминоацил-тРНК атакует электрофильную карбонильную группу (*) в сложной эфирной связи между пептидом (или формил- метиониновой затравкой) и другой тРНК. В результате этого аминокислотная цепь удлиняется на один остаток, а молекула тРНК деацилируется:
![]()
Молекулы 16S рРНК и 23S рРНК образуют три сайта — Р, А и Е, каждый из которых представлен субсайтами в обеих субъединицах рибосомы. P-сайт (от англ. peptide) связывает пептидил- тРНК, A-сайт (от англ. amino acid) связывает аминоацил-тРНК, а Е-сайт (от англ. exit) связывает деацилированную тРНК.
Рабочий цикл рибосомы состоит из четырех этапов, или состояний (рис. 17, А-Г).
1. В исходном состоянии Р/Р-А/А (рис. 17, А) пептидил-тРНК находится в P-сайте, аминоацил- тРНК в A-сайте, Е-сайт свободен. Специфическая аминоацил-тРНК связывается с A-сайтом при помощи фактора элонгации EF-Tu. Для этого используется энергия гидролиза ГТФ. Тройственный комплекс аминоацил-тРНК/(ЕЕ-Тu)-ГТФ прочно связывается с рибосомой только если антикодон комплементарен кодону в декодирующем субсайте А.
2. В «претранслокационном» состоянии Р/Р-А/А (рис. 17, Б) происходит пептидилтрансферазная реакция. При этом аминокислота, находящаяся в A-сайте, образует связь с пептидом (или формилметионином, если цепь инициируется), который находится в P-сайте. В обоих случаях дипептид или полипептидная цепь, удлиннившаяся на один аминокислотный остаток, переносятся в А-сайт.
Чтобы следующая молекула аминоацил-тРНК могла попасть в A-сайт, пептидил-тРНК должна освободить его и перейти в P-сайт. Этот процесс называется «транслокацией». При транслокации взаимодействующие друг с другом тРНК и мРНК перемещаются внутри рибосомы на расстояние до 50А.
3. В «гибридном транслокационном» состоянии Е/Р-Р/А (рис. 17, В) связанный с пептидом конец тРНК перемещается на большой субъединице из А-субсайта в Р-субсайт, а акцепторный ССА- конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. Этот этап транслокации напоминает «эффект домино» и зависит от рРНК. Его механизм еще неизвестен.
4. В «однородном транслокационном» состоянии Е/Е-Р/Р (рис. 17, Г) антикодоновый конец тРНК, связанной с пептидом, перемещается на малой субъединице из А-субсайта в ее же Р-субсайт, а антикодоновый конец деацилированной тРНК перемещается из Р-субсайта в Е-субсайт. В результате этого мРНК сдвигается в малой субъединице на один кодон. Теперь A-сайт может принять следующую молекулу аминоацил-тРНК, а деацилированная молекула тРНК может покинуть рибосому. Хотя этот этап транслокации зависит от рРНК, он ускоряется фактором элонгации EF-G, который использует энергию гидролиза ГТФ.
Действие многих антибиотиков (канамицина, неомицина, олеандомицина, стрептомицина, тетрациклина, хлорамфеникола и др.) основано на их связывании с факторами элонгации и сайтами, которые образует рРНК.
Различие между прокариотным и эукариотным сценариями трансляции. В разделе 5.1 мы уже говорили о том, что эукариотная транскрипция разделена с трансляцией в пространстве и времени. Транскрипция вместе с процессингом РНК происходят в нуклеоплазме, а трансляция, в зависимости от типа клеток, осуществляется преимущественно в цитозоле или на шероховатом эндоплазматическом ретикулуме (англ. rough endoplasmic reticulun, RER). Интегральные белки встраиваются в мембрану RER котрансляционно, а секретируемые белки выделяются в полость цистерны RER через тороидальный переходник между выходным порталом рибосомы и мембранным транслоконом (его образует белок Sec61).
У прокариотов не существует пространственно-временной изоляции процессов транскрипции и трансляции. Цитоплазматические рибосомы присоединяются к 5'- концу мРНК еще до завершения образования короткоживущего транскрипта (см. раздел 8.3.10.1). Котрансляционная инсерция интегральных белков известна только на примере «шероховатых тилакоидов» цианобактерий (см. раздел 8.3.5). Гидрофобные белки при помощи SRP-частиц презентируются транс локону — компоненту генеральной системы секреции Sec (см. раздел 8.6.1).
8.3.8.1. Рибосомы бактерий
Состав бактериальной рибосомы и ее биогенез изучены в основном на примере Е. coli.
Состав. Большая и малая субъединицы бактериальной рибосомы имеют коэффициенты седиментации 50S и 30S (молекулярная масса соответственно 1,5 • 103 кДа и 750 кДа).
Большая субъединица состоит из двух молекул рРНК (23S и 5S) и 34 молекул белка (L1-L34). Малая субъединица образована одной молекулой рРНК (16 S рРНК) и 21 молекулой белка (S1-S21). Молекула 23S рРНК имеет массу 103 кДа, молекула 16S рРНК — 500 кДа, а молекула 5S рРНК — 40 кДа. За исключением двух случаев (L7 и L12), все рибосомные белки представлены только одной копией, имеют молекулярную массу 5-60 кДа и состоят из 45-555 аминокислотных остатков.
Биогенез. Гены, кодирующие рРНК, образуют один или несколько оперонов на бактериальной хромосоме и считываются в виде полицистронной мРНК. Структурные гены (цистроны) рРНК универсально расположены в последовательности 16S-23S-5S. Иногда между ними находятся «внутренние транскрибируемые спей- серы», в состав которых могут входить последовательности TPHKSer, tPHKGlu и тРНКIlе. Процессинг первичного транскрипта осуществляется путем расщепления при помощи РНКаз III и Е, а также путем метилирования.
Гены, кодирующие рибосомные белки, образуют на бактериальной хромосоме несколько оперонов. Наряду с цистронами рибосомных белков, в них входят гены факторов трансляции и субъединиц РНК-полимеразы.
Биогенез рибосомы начинается с транскрипции оперона рРНК, после чего первичный транскрипт расщепляется на 16S рРНК, 23S и 5S рРНК. Сборка субъединицы рибосомы занимает 0,5-1,5 мин и представляет собой строго координированный многоступенчатый процесс, или «карту сборки» (англ. assembly map). Естественно, что в случае 50S-субъединицы она более сложна. Молекулярным каркасом служит рРНК, которая в строгой последовательности одевается рибосомными белками.
Рибосомы постоянно диссоциируют на большую и малую субъединицы. Моносома образуется, только когда с малой субъединицей связывается мРНК. Затем при участии факторов инициации мРНК связывает тРНК, и только в этом случае к малой субъединице прикрепляется большая. Рибосома считывает мРНК до стоп- кодона и снова диссоциирует на субъединицы.
Скорость роста бактерий ограничивается скоростью синтеза белка, которая при 37°С составляет ~20 аминокислотных остатков • в секунду. Поэтому чтобы рост ускорился, бактерия должна содержать больше рибосом. В медленно растущих клетках рибосомы составляют 15% клеточной массы, а в быстро растущих клетках — до 50% (у эукариотов — не больше 5%); во втором случае скорость образования рибосом возрастает почти в 100 раз. При быстром росте бактерия (например, Е. coli) делится каждые 20 мин. В этих условиях клетка содержит до 105 рибосом, число которых удваивается к началу каждого клеточного цикла. Неудивительно, что на биосинтез рибосом направлено до 50% энергетических ресурсов, 70% суммарной РНК составляет рРНК, а на рибосомные белки приходится 30% всех синтезируемых белков.
При попадании бактерий в более благоприятную для роста среду скорость транскрипции увеличивается за несколько секунд. В число активируемых оперонов входят, в частности, опероны рРНК и рибосомных белков.
8.3.8.2. Рибосомы архей
К началу 1980-х годов стало известно, что ряд представителей порядков Halobacteriales и Methanococcales имеет рибосомы бактериального 70S-типа, состоящие из субъединиц 52S/31S и содержащие по одной молекуле 23S рРНК, 16S рРНК и 5S рРНК.
Позднее было установлено, что у других членов филы АН Euryarchaeota (пор. Methanomicrobiales, Thermoplasmatales и Thermococcales), а также у некоторых членов филы AI Crenarchaeota (пор. Desulfurococcales, Thermoproteales и Sulfolobales) рибосомы имеют константу седиментации 80S. Более крупный размер этих рибосом объясняется не увеличением суммарной массы рРНК (они содержат 23S рРНК, 16S и 5S рРНК, однако у них нет «эукариотной» 5,8S рРНК), а присутствием дополнительных рибосомных белков. Если в рибосомах бактерий их число составляет 53-54, то здесь их 60-65. Эти дополнительные белки относятся главным образом к
S-субъединице.
По морфологии археотные рибосомы второго типа отличаются от бактериальных и похожи на эукариотные (в частности, для S-субъединицы характерно сочетание выступов в форме «клювика» и двух «лапок»).
Рибосомные белки. Рибосомы экстремальных галофилов и некоторых галофильных метаногенов состоят преимущественно из кислых белков, что, видимо, является приспособительной чертой в среде обитания с высокой соленостью. В то же время рибосомные белки серазависимых термоацидофилов, как и рибосомные белки бактерий и эукариотов, проявляют щелочные свойства.
Археи и бактерии имеют взаимно гомологичные рибосомные белки Ll, Lll, S8 и S17. К числу взаимно гомологичных рибосомных белков архей и эукариотов относятся L2, L3, L5, L6, L10, L12, L14, L15, L22, L23, L30, S5, S7, S9, S10-12, S15 и S19. Некоторые рибосомные белки представлены как у архей, так и у бактерий (например, L1), однако отсутствуют у эукариотов. В свою очередь, некоторые рибосомные белки, одинаковые у архей и у эукариотов (например, L12), отсутствуют у бактерий.
Археотные рРНК. Молекулы 23S и 16S рРНК у архей не больше, чем на 10% гомологичны своим эквивалентам в составе бактериальной рибосомы (23S и 16S) и эукариотной рибосомы (28S и 16S). Вторичная структура рРНК у всех трех доменов имеет специфические особенности, связанные с появлением вставок или делеций. Однако рРНК архей в этом отношении мозаична. В одних участках вторичной структуры она сходна с рРНК бактерий, а в других — эукариотов.
«Мозаичность» археотных рибосом. Если мы сравним такие качественные признаки рибосомы, как нуклеотидная последовательность рРНК, аминокислотная последовательность рибосомных белков и детали процесса трансляции, то увидим, что так называемые «прокариотные» рибосомы образуют неоднородную группу — бактериальные рибосомы не гомологичны археотным рибосомам. Гетерогенность проявляется и в пределах группы археотных рибосом, которые мозаично сочетают в себе бактериальные и эукариотные признаки.
С одной стороны, рибосомы архей по первичной структуре рРНК четко дифференцируются как от рибосом бактерий, так и от рибосом эукариотов. С другой стороны, рибосомы архей по первичной структуре белков принадлежат либо к бактериальному типу, либо имеют черты рибосом эукариотного типа.
В итоге, по деталям процесса трансляции археи сходны либо с бактериями, либо с эукариотами. Как и у бактерий, археотная мРНК содержит последовательность
Шайна-Дальгарно, которая связывается со специфическим сайтом на S-субъединице. В то же время археи, как и эукариоты, используют для инициации полипептидной цепи не формилметионин, а метионин, который доставляется в рибосому специфической тРНКМet.
Рибосомы архей также проявляют мозаичную чувствительность к ингибиторам трансляции. В отличие от бактерий, археи устойчивы к рифампицину и хлорамфениколу, однако чувствительны к дифтерийному токсину.
Дифтерийный токсин переносит остаток АДФ-рибозы от НАД на эукариотный фактор элонгации ЕF-2 (аналог бактериального фактора элонгации ЕF-G). Мишенью для ковалентного присоединения АДФ-рибозы к консервативному С-концу фактора элонгации ЕF-2 служит необычная аминокислота дифтамид, которая представляет собой посттрансляционно модифицированный гистидин.
Бактериальный фактор элонгации ЕF-G не поддается АДФ-рибозилированию, в то время как археотный фактор элонгации ЕF-2 содержит дифтамид или его дезаминированное производное — дифтин.
Фактор элонгации, связывающий аминоацил-тРНК, сходен у всех трех глобальных доменов. Однако по чувствительности к антибиотику кирромицину он у архей ближе к эукариотному ЕF- Iα, чем к бактериальному ЕF-Тu.
8.3.9. Цитоскелет
Представления о цитоскелете сложились в цитологии позже, чем о других органеллах, и этот термин использовался только по отношению к эукариотам.
В 1917 г. английский эмбриолог Эдвин Конклин (Е. Conclin) пришел к выводу, что ригидность клеточной оболочки яйцеклетки, а также фиксированное положение ее органелл обеспечиваются эластичным цитоплазматическим матриксом. В 1930 г. английский цитолог Рудольф Питерс (R. Peters) предположил, что метаболические процессы координируются сложно организованной белковой сетью, и что часть ее образуют ферменты. На следующий год французский эмбриолог Поль Вэнтребер (P. Wintrebert) назвал эту сеть цитоскелетом (франц. cytosquelette). В общепринятой английской транскрипции (cytoskeleton) термин «цитоскелет» окончательно ввел в научный обиход в 1936 г. известный английский эмбриолог Джозеф Нидэм (J. Needham).
Цитоскелет эукариотов состоит из микротрубочек, микрофиламентов и промежуточных филаментов. Он отвечает за форму клетки и взаимное расположение органелл, а также обеспечивает везикулярный транспорт, поляризацию, деление, работу локомоторных систем, сигналинг, прикрепление к субстрату и т. д.
Центральным компонентом цитоскелета эукариотов является актин. В определенных условиях мономерный актин с молекулярной массой 43 кДа (G-актин) полимеризуется в протофиламенты (F-актин), отвечающие за форму многих эукариотных клеток.
Еще недавно считалось, что прокариоты лишены цитоскелета, и что у них нет актиноподобных белков. В основе этих представлений лежали классические опыты на Е. coli, где муреиновый слой разрушали лизоцимом (см. раздел 8.4.3.1), в результате чего палочковидные клетки приобретали сферическую форму. Мутации по генам, отвечающим за биосинтез и обновление мурерина, также приводили к изменению формы клетки. Поскольку при этом в цитоплазме не удавалось выявить филаментозных структур, напрашивался вывод, что у прокариотов нет цитоскелета, гомологичного или аналогичного цитоскелету эукариотов. Однако в настоящее показано, что цитоскелет является неотъемлемой частью прокариотной клетки.
В биологии прокариотов термином «цитоскелет» обозначаются три типа клеточных структур:
1. Морфоскелет (англ. morphology — морфология), или морфогенетический цитоскелет;
2. Дивискелет (англ. division — деление), или цитокинетический цитоскелет;
3. Энзоскелет (англ. enzyme — фермент), или метаболический цитоскелет.
Они принимают участие соответственно в мофогенезе, делении и конструктивном метаболизме (рис. 18).
Рис. 18. Классификация цитоскелетных структур у прокариотов.
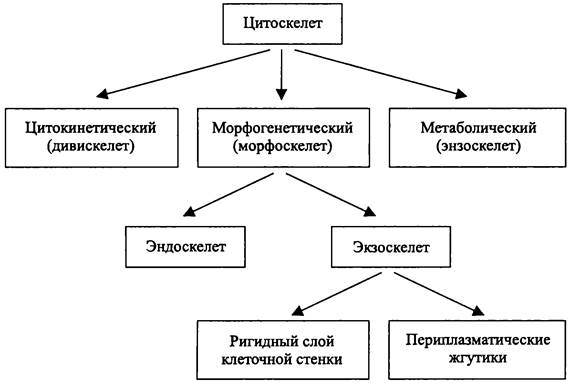
8.3.9.1. Морфоскелет
Под морфоскелетом, или собственно цитоскелетом понимают структуры, отвечающие за целостность клеточной оболочки, а также за индивидуальную форму бактерии или археота.
Морфоскелет универсально представлен эндоскелетом (англ. endoskeleton) — системой цитоплазматических микрофиламентов.
Наряду с эндоскелетом прокариотная клетка обычно имеет экзоскелет (англ, exoskeleton) — клеточную стенку, основу которой составляют ригидные слои, расположенные в периплазматическом и/или экзоплазматическом компартменте.
В морфогенетическом плане прокариоты занимают уникальное положение в мире живых организмов, поскольку в большинстве случаев индивидуальная форма клетки у них обеспечивается совместно эндоскелетом и экзоскелетом.
Форма прокариотов. Мир прокариотных организмов состоит из множества видоспецифичных клеточных форм. Кроме того, некоторые бактерии и археи представлены многоликими формами — имеются в виду бактериальные (Mycoplasma sp.) и археотные (Thermoplasma sp.) микоплазмы. Они лишены клеточной стенки и образуют «ассорти» из сферических, лопастных, булавовидных, гантелевидных и нитчатых клеток. Иногда индивидуальное морфологическое разнообразие проявляют и прокариоты с клеточной стенкой. Например, популяции археота Halogeometricum borinquense состоят из палочковидных, «квадратных», «треугольных» и «овальных»
особей. В обоих случаях такое цитологическое явление называется плеоморфизмом (греч. pleion — большой по числу и morphe — форма).
Форма прокариотной клетки консервативна, и это широко учитывается в феносистематике. Иногда морфологические особенности находят отражение в родовом названии или в видовом эпитете. Примерами служат бактерия рода Stella (лат. Stella — звезда) и археот Haloarcula quadrata (греч. alos — соль, лат. arcula — коробочка и лат. quadratus — квадратный, «галофил в форме квадратной коробочки»).
Клетки бактерий чаще всего имеют форму фигур вращения. Самые простые из них:
1. Сфера, или кокк (рис. 19,1). Пример — Streptococcus mutans (фила ВХIII Firmicutes).
2. Эллипсоид (рис. 19, 2). Пример — Chromatium vinosum (класс «Gammaproteo- bacteria»).
3. Цилиндр с полусферическими концами, или бацилла (рис. 19,3). Пример — Bacillus thuringiensis (фила ВХШ Firmicutes).
4. Цилиндр с коническими концами, или веретено (рис. 19,4). Пример — Cystobacter fuscus (класс «Deltaproteobacteria»).
5. Удлиненная палочка, или мицелиальная нить (рис. 19,5). Пример — Thermothrix thiopara (класс «Betaproteobacteria»).
При сжатии сферической клетки с противоположных сторон она превращается в «диск» (рис. 19,6). Пример — Crenothrix polyspora (фила ВХХ Bacteroidetes).
В результате радиальной деформации диска его поперечный срез становится овальным (рис. 19,7). Пример — Crinalium epipsammum (фила ВХ Cyanobacteria). Поперечный срез диска также бывает полукруглым (рис. 19,8). Пример — Simonsiella steedae (класс «Betaproteobacteria»). Наконец, поперечный срез диска бывает лопастным (рис. 19,9). Пример — Starria zimbabweënsis (фила ВХ Cyanobacteria).
Изгиб палочки в одной плоскости придает ей форму половины тора, или полумесяца (рис. 19,10). Пример — Rhodocyclus purpureus (класс «Betaproteobacteria»).
При закручивании палочки одновременно по трем осям бактерия приобретает форму вибриона, т. е. отрезка спирали (рис. 19,11). Пример — Vibrio cholerae (класс «Gammaproteobacteria»). Она также принимает форму спириллы, т.е. спирали с несколькими полными витками — растянутой или плотно сжатой (рис. 19,12). Пример — Spirillumvolutans (класс «Betaproteobacteria»).
Палочки могут за счет ветвления принимать Y-образную форму (рис. 19,13). Пример — Mycobacterium smegmatis (фила BXIV Actinobacteria).
В свою очередь, мицелиальные нити бывают слабо или сильно разветвленными (рис. 19,14). Пример — Streptomyces carpinensis (фила BXIV Actinobacteria).
Встречаются бактерии бугристой формы (рис. 19,15). Пример — Nitrosolobus multiformis (филогенетическое положение не установлено).
Наконец, бактерии могут образовывать выросты нитевидной и конической формы, или простеки (рис. 19,16). Пример — Ancalomicrobium adetum (класс «Alphaproteobacteria»).
Что касается архей, то среди них можно найти все «бактериальные» варианты формы (рис.19,1-16).
Некоторые экстремально галофильные археи имеют уникальную форму «квадратов» (рис. 19,17) или «треугольников» (рис. 19,18). Примеры — Haloarcula quadrata и Halogeometricum borinquense (фила АН Euryarchaeota).
Рис. 19. Основные варианты морфологии прокариотной клетки.

Главный формообразующий элемент прокариотной клетки — это ригидный слой клеточной стенки (см. разделы 8.4.3 и 8.5.2). Второй функцией этого слоя служит противодействие тургору, который составляет 1-5 бар (0,1-0,5 мПа), а в отдельных случаях даже достигает 20 бар.
Для снижения тургорного давления (или в противовес ему) клетки могут использовать четыре стратегии.
Первая стратегия — это экскреция воды, поступающей в клетку по градиенту водного потенциала. Прокариоты, в отличие от эукариотов, не имеют сократительных вакуолей, и только немногие из них могут выделять излишки воды через аквапорины (см. II том учебника).
Вторая стратегия — это выравнивание давления по обе стороны СМ путем накопления в цитоплазме осмотически активных веществ, или осмопротекторов. Наиболее эффективно, хотя и уникальным способом это делают экстремально галофильные археи, в цитоплазме которых снакапливаются катионы К+ в концентрации, равной концентрации катионов в окружающей среде.
Третья стратегия — это образование ригидного экзоскелета, и подобным образом поступают многие протисты, все грибы и растения, а также большинство бактерий и архей.
Четвертая стратегия — это использование эндоскелета.
Для обеспечения осмотического комфорта прокариоты больше полагаются на экзоскелет, в то время как эндоскелет у них играет преимущественно морфогенетическую роль.
Эндоскелет. Первые данные о том, что клетки бактерий и архей (Е. coli, В. subtilis, Methanococcus voltae и др.) содержат сеть белковых фибрилл, лежащую под СМ, были получены в конце 1990-х годов с помощью метода иммуноэлектронной микроскопии. В качестве зондов использовались моноклональные антитела к С-концевому участку актина, содержащему высококонсервативный мотив Ser-Gly- Pro-Ser-Ile-Val-His-Arg-Lys-Cys-Phe.
В дальнейшем выяснилось, что за образование белкового эндоскелета отвечает оперон mre (от англ. murein duster е), обеспечивающий образование муреинового саккулуса. Одним из его продуктов является актиноподобный белок МгеВ, который полимеризуется в протофиламенты, сходные с F-актином. Показано, что у В. subtilis он образует спираль, подстилающую СМ (рис. 20, А).
Рис. 20. Молекулярная подоплека «несферической» формы бактерий. А — палочковидная форма, приобретенная с помощью цитоскелетного белка МгеВ; Б — спиральная форма, приобретенная с помощью цитоскелетного белка СгеS.

Белок МrеВ и сходные с ним актиноподобные белки, например, мы, широко распространены у палочковидных, нитевидных и спиралевидных видов. Из них состоит эндоскелет, который отвечает за «несферическую» форму бактерий. Молекулярная гомология между МrеВ и актином говорит об общем и очень древнем происхождении этих цитоскелетных белков.
Более сложную форму клетке придают протофиламенты других белков. В частности, вибриоидная или спиральная форма стебельковой бактерии Caulobacter crescentus (лат. caulis — стебель и crescentus — полумесяц) обеспечивается цитоске- летным белком кресцентином CreS. Он гомологичен цитокератинам — белкам промежуточных филаментов (intermediaryfilament, IF) животных, и имеет сходную с ними мультидоменную структуру. Кресцентиновая спираль расположена вдоль вогнутой стороны клетки. Предполагается, что белок МгеВ придает клетке форму палочки, в то время как белок CreS закручивает ее в спираль (рис. 20, Б).
Форма бактерий, не имеющих клеточной стенки, полностью зависит от эндоскелета. Например, у Mycoplasma pneumoniae существует так называемая «терминальная органелла» (англ. therminal organelle). Это бульбовидный вырост клетки, который окружен СМ и содержит филаментозный цитоскелет, состоящий из
белков семейства HMW и вспомогательных белков. Окончательные выводы об их сходстве (или отличии) с известными цитоскелетными белками еще не сделаны. Терминальная органелла полифункциональна. Она принимает участие в сегрегации дочерних хромосом, цитокинезе и скользящей подвижности (см. раздел 8.6.2.1), а также содержит адгезины, прежде всего белок Р1, которые связываются с гликолипидами и сиалолипидами на поверхности эпителиальных клеток хозяина.
Интересно, что за форму пластид, которые произошли от бактерий и в ходе внутриклеточной эволюции потеряли ригидную стенку, отвечает не актиноподобный белок, а тубулиноподобный белок FtsZ (см. III том учебника). От какого цитоскелетного белка зависит форма митохондрий, еще не ясно.
В отличие от бактерий, сведений об эндоскелете у архей очень мало. По некоторым данным, клетки Thermoplasma sp. содержат микрофиламенты, состоящие из актиноподобного белка. На примере гипертермофила Sulfolobus shibatae показано, что шаперонины (раздел 8.3.10.2) образуют филаментозные структуры диаметром 10 нм, одной из функций которых, возможно, является участие в формировании эндоскелета.
Экзоскелет. Термином «экзоскелет» в цитологии прокариотов обозначаются разнообразные ригидные структуры, расположенные вне СМ. За счет (не)ковалентных внутренних взаимодействий они приобретают жесткую конфигурацию, которая навязывается протопласту. В изолированном виде эти структуры, как скорлупа ореха, продолжают сохранять характерную форму клетки.
Наиболее распространенным типом экзоскелета служат ригидные слои, которые имеют разную локализацию и различаются по химическому составу.
Бактерии, обладающие грамотрицательным морфотипом, образуют периплазматический экзоскелет в виде муреинового саккулуса (см. раздел 8.4.3.1) или белкового саккулуса (см. раздел 8.4.3.2).
Бактерии, обладающие грамположительным морфотипом, образуют экзоплазматический экзоскелет в виде муреин-тейхоатного саккулуса (см. раздел 8.5.2.1), муреин-миколил-арабиногалактанового саккулуса (см. раздел 8.5.2.2) или паракристаллического поверхностного слоя (см. раздел 8.5.2.3).
В свою очередь, археи образуют экзоплазматический экзоскелет в виде паракристаллического поверхностного слоя (см. раздел 8.5.3.1), псевдомуреинового саккулуса (см. раздел 8.5.3.2) или гетерополисахаридного слоя (см. раздел 8.5.3.3).
Совершенно другой тип экзоскелета имеют спирохеты (фила BXVII Spirochaetes). В данном случае ведущая роль принадлежит не сплошному ригидному слою, а осевой структуре — совокупности периплазматических жгутиков (см. раздел 8.6.2). Характерным примером является возбудитель болезни Лайма Borrelia burgdorferi, которая по размеру превышает большинство патогенных бактерий (0,3x20 мкм). На обоих концах клетки расположены по 7-11 периплазматических жгутиков, которые перекрываются в экваториальной зоне, образуя периплазматический пучок, или аксиальную нить. Ее спиральная форма передается гибкому цитоплазматическому цилиндру и гибкой ОМ. Как показал результат инсерционного мутагенеза, при блокаде гена flaB, кодирующего главный флагеллин нити жгутика, спирохеты перестают образовывать периплазматические жгутики. Одновременно они теряют свою спирализацию и приобретают палочковидную форму.
8.3.9.2. Дивискелет
Под названием «дивискелет», или «септосома» (англ. septosome) известна система микрофиламентов, обеспечивающая цитокинез. Благодаря ему сестринские хромосомы обособляются внутри своих цитоплазматических компартментов.
В состав септосомы у бактерий и архей входят белки FtsZ, FtsA и МuкВ, которые по ряду свойств (молекулярная масса, третичная структура, способность связывать и гидролизовать нуклеозидтрифосфаты) напоминают белки цитоскелета у прокариотов — тубулин и актин, хотя не гомологичны им. Эти белки будут подробно рассмотрены в III томе учебника.
8.3.9.3. Энзоскелет
По характеру локализации клеточные ферменты подразделяются на мембраносвязанные и растворимые (цитозольные, периплазматические и экзоплазматические).
Растворимые цитозольные ферменты, вопреки своему названию, образуют филаментозные структуры и нередко функционируют как часть мультиэнзимного комплекса. По ним, как по конвейеру, передвигаются субстраты и интермедиаты метаболических путей. Для обозначения данного варианта цитоскелета в середине 1990-х годов был предложен термин «энзоскелет» (англ. ensoskeleton).
Энзоскелет не только создает функциональный ансамбль ферментов, но и взаимодействует с нуклеоидом, рибосомами и мембранным аппаратом.
Характерным примером такого мультиэнзимного комплекса является триптофансинтаза Salmonella typhimurium. Это своего рода молекулярный туннель, по которому происходит облегченная диффузия индола.
В свою очередь, из клеток Е. coli выделен компартментализованный суперкомплекс, содержащий стехиометрический набор ферментов, участвующих в гликолизе. Молекулярная масса суперкомплекса составляет 1650 кДа, и он образует частицы диаметром более 30 нм. С помощью ферментов-л инке ров отдельные ферменты объединяются в цепочки. Те, в свою очередь, формируют цитоплазматическую сеть, в состав которой дополнительно входит комплекс пируватдегидрогеназы, состоящий из 8 тримеров липоамидредуктазы/трансацетилазы, 12 молекул дигидролипоил-дегидрогеназы и 24 молекул пируватдекарбоксилазы. Наконец, с суперкомплексом связана регуляторная киназа и фосфатаза. Совместно они образуют частицу, по размеру превосходящую рибосому (~ 25 нм).
Бактериальные рибосомы и другие компоненты аппарата трансляции также образуют энзоскелет. Он взаимодействует с хромосомой и мембранами. Возможно, в его отдельных участках образуются структуры, аналогичные эукариотному ядрышку.
8.3.10. Генеральные компартменты для посттранскрипционного и посттрансляционного процессинга
Среди регуляторных механизмов прокариотной клетки первое место занимает контроль репликации и транскрипции. Следующим по важности является посттранскрипционный и посттрансляционный процессинг. Благодаря ему поддерживается гомеостаз, исправляются ошибки транскрипции и трансляции, достигается адаптация к условиям окружающей среды (которые часто являются экстремальными), а также обеспечиваются условия, выгодные для взаимодействия внутри популяции и с другими компонентами биоты.
8.3.10.1. Деградосомы
Посттранскрипционный процессинг мРНК, в частности, выражается в ее деградации путем расщепления на поли- и мононуклеотиды. Этот процесс, изученный в основном на примере Е. coli, компартментализован в деградосоме — мультиферментном комплексе молекулярной массой 160-460 кДа и константой седиментации 8-16S.
В отличие от эукариотов, у которых время жизни мРНК измеряется многими часами, бактерии синтезируют нестабильную мРНК, которая живет не дольше нескольких минут. «Короткий век» этих макромолекул связан главным образом с тем, что при вегетативном существовании, а также при клеточной (де)дифференцировке бактерии быстро и адекватно реагируют на изменения в окружающей среде. В обоих случаях они должны репрограммировать биосинтез белка путем изменения количественного и качественного состава пула мРНК.
Наряду с контролем промоторов, центральным способом регулирования уровня экспрессии генов является деградация мРНК. С одной стороны, независимо от стабильности мРНК, соответствующие клеточные механизмы рано или поздно выводят ее из оборота. С другой стороны, возможность обновления пула мРНК позволяет перенастраивать белоксинтезирующий аппарат в соответствии с конкретными особенностями адаптации к изменяющимся параметрам окружающей среды. Наконец, поскольку особенностью инфраструктуры бактериального генома служит объединение структурных генов в опероны, с которых считывается полицистронная мРНК, при ее частичной деградации образуется набор мРНК с разным временем жизни, что дополнительно влияет на паттерн трансляции.
Деградация мРНК у прокариотов осуществляется с помощью экзорибонуклеаз и эндорибонуклеаз. В отличие от эукариотных мРНК, прокариотные мРНК легче поддаются деградации, поскольку для этого не требуется предварительное деаденилирование, т. е. ферментативное удаление защитных полиадениловых кэпов на 3'-конце.
Каркас деградосомы составляет эндорибонуклеаза Е. Этот фермент имеет молекулярную массу 118 кДа и гидролизует мРНК по 3'-концу на относительно крупные полинуклеотидные фрагменты. Его N-участок содержит активный центр, а С-участок служит платформой для сборки остальных деталей деградосомы (рис. 21).
Рис. 21. Деградосома. RhlB — геликаза; PNP — полинуклеотидфосфорилаза (экзорибонуклеаза); РРК — полифосфаткиназа; (Ф)n—полифосфат; Ф — фосфатная группа.

Вторым ферментом деградосомы является экзорибонуклеаза PNP (от англ. polynucleotide phosphorylase; гомотример субъединиц 78 кДа). Она осуществляет фосфоролиз мРНК, т. е. с помощью неорганического фосфата отщепляет от 3'-конца по одному нуклеозиддифосфатному остатку.
Третьим ферментом деградосомы является геликаза RhlB (от англ. RNA helicase type В; димер субъединиц 50 кДа). За счет энергии гидролиза АТФ этот фермент расплетает дуплексы, или двухцепочечные участки в молекуле мРНК, тем самым подготавливая ее к расщеплению нуклеазами.
Четвертый фермент деградосомы — энолаза, или фосфоэнолпируват-гидратаза, которая катализирует обратимую реакцию гликолиза: 2-фосфоглицерат + Н2О <-> фосфоэнолпируват. Какую роль в данном случае играет этот фермент, неясно.
Роль пятого фермента деградосомы, возможно, выполняет РНКаза III, которая расщепляет двухцепочечные участки РНК.
Наконец, дополнительными компонентами деградосомы служат молекулярные шапероны GroEL и DnaK.
С деградосомой тесно ассоциирована, хотя непосредственно не входит в число ее структурных компонентов, полифосфаткиназа РРК (от англ, polyphosphate kinase; гомотетрамер субъединиц 80 кДа). Этот фермент регенерирует АТФ для геликазы согласно уравнению реакции (Ф)n+ АДФ —> (Ф)n-1 + АТФ (см. раздел 8.3.11.4).
8.3.10.2. Шаперонины
Пострансляционный процессинг белка происходит в двух генеральных микрокомпартментах — шаперонинах и протеасомах. Они представляют собой гомогенные популяции ультрамикроскопических частиц, расположенных в цитоплазматическом компартменте.
Молекулярные шапероны. Термин «молекулярные шапероны» (англ. chaperone — лицо, обеспечивающее нормальные взаимоотношенения между людьми) служит для обозначения гетерогенного молекулярного суперсемейства, в которое входят более 50 белков, различающихся по первичной последовательности и третичной структуре. Эти белки имеются у представителей всех трех филогенетических доменов и выполняют такие функции, как:
— фолдинг (англ. fold — складывать), или правильная укладка белка при его синтезе de novo;
— сборка правильно уложенных белков в надмолекулярные комплексы;
— таргетинг (англ. target — мишень), или доставка белков в определенные области клетки и перенос белков через мембраны;
— изменение конформации регуляторных белков;
— предотвращение образования патологических белковых агрегатов, а также их разборка;
— ренатурация белка, поврежденного при стрессе;
— придание обновляемому белку такой конформации, которая благоприятна для его протеолитического расщепления.
Широкий спектр действия молекулярных шаперонов определяется их свойством взаимодействовать с участками полипептидной цепи, играющими критическую роль при внутримолекулярной или межмолекулярной самосборке, а также их способностью стабилизировать эти участки. Тем самым молекулярные шапероны отслеживают конформационное состояние белка на всем протяжении жизни его молекулы, начиная с трансляции.
Хотя информация о третичной структуре белка заложена в его первичной структуре, спонтанный и термодинамически выгодный фолдинг возможен только в разбавленном растворе. При высокой концентрации белков в цитоплазме и особенно при супероптимальной температуре происходят сильные гидрофобные взаимодействия, которые приводят к неправильному складыванию белка и его агрегации.
Кроме того, скорость укладки белка намного выше скорости трансляции. Для того, чтобы лидирующий конец полипептидной цепи не вступал в ошибочное взаимодействие сам с собою или с другими белками, т. е. чтобы его конформация в конечном счете оказалась правильной, требуется коррекция. Ее осуществляют молекулярные шапероны, а также содержащие их микрокомпартменты — шаперонины (см. ниже).
Молекулярные шапероны прежде всего участвуют в укладке белка при его синтезе de novo. Значительная часть полипептидной цепи, появляющейся из выходного туннеля рибосомы, взаимодействует со связанным молекулярным шапероном. В случае бактерий это «триггер-фактор» (англ. trigger factor, TF), а в случае архей и эукариотов — «комплекс, ассоциированный с нативным полипептидом» (англ, nascent polypeptide-associated complex, NAC). Со связанным шапероном функционально взаимодействуют растворимые молекулярные шапероны — DnaK (в случае бактерий), комплекс DnaK с префолдином (в случае архей) или комлекс Hsp70 с префолдином (в случае эукариотов).
В результате взаимодействия с этими молекулярными шаперонами новая полипептидная цепь, как правило, принимает правильную конформацию. При наличии отклонений последняя корректируется путем процессинга белка внутри шаперонина.
Шаперонины также корректируют конформацию ранее образованных полипептидных цепей, которая по той или иной причине стала ненативной.
Глобальное значение шаперонинов подтверждается тем, что дефектные по ним мутанты нежизнеспособны.
В случае бактерий шаперонин представлен комплексом GroES/GroEL, в случае архей — термосомой (см. ниже), а в случае эукариотов — комплексом ССТ.
Движущей силой конформационной перестройки белка служит его собственная свободная энергия, а также свободная энергия гидролиза АТФ.
Одни молекулярные шапероны присутствуют в клетке постоянно, и при нефизиологической температуре или при действии токсичных агентов уровень их экспрессии либо сохраняется прежним, либо повышается. Другие молекулярные шапероны являются стрессовыми белками, например, белки теплового шока (англ, heat-shock protein, Hsp); они индуцируются в неблагоприятных условиях и служат ключевыми факторами для выживания.
В соответствии со своим размером, молекулярные шапероны подразделяются на группу больших (~60 кДа) и малых (~10 кДа). Малые молекулярные шапероны называют «младшими» (англ, junior)— в основном из-за того, что механизм их действия менее изучен, чем у «старших» (англ. senior) шаперонов. Большинство младших шаперонов образует глобулярные полидисперсные олигомеры, суммарная молекулярная масса которых 200-800 кДа.
Шаперонины; общие сведения. Как уже указывалось, молекулярные шапероны вместе со своими кофакторами образуют шаперонины (англ. chaperonin) - микрокомпартменты, предназначенные для коррекции конформации неправильно сложенного белка.
Субъединицы шаперонинов принадлежат к группе больших молекулярных шаперонов (≈ 60 кДа) и собираются в тороидальную структуру, состоящую из двух колец.
Существуют две филогенетически обособленные группы шаперонинов.
К группе I относится шаперонин бактерий (содержащий шаперон GroEL), а также его гомолог в митохондриях (содержащий шаперон Hsp60) и пластидах (содержащий шаперон RuBP; от англ. RuBisCO subunit binding protein). Основу этих мик- рокомпартментов образуют два кольца, каждое из которых состоит из 7 субъединиц молекулярного шаперона GroEL.
К группе II относятся шаперонины архей и эукариотов. Эти микрокомпартменты состоят из двух колец, каждое из которых образовано 8-9 субъединицами молекулярного шаперона TF55 и молекулярного шаперона ССТ.
Шаперонины бактерий. Это олигомерный комплекс, состоящий из 14 идентичных субъединиц шаперона GroEL (57 кДа). Согласно результатам рентген-кристаллографии с уровнем разрешения 2,8А, он имеет форму полого цилиндра, высота которого составляет 14,5 нм, а диаметр 13,5 нм. Центральная полость цилиндра GroEL сообщается с окружающей средой через концевые отверстия диаметром 4,5 нм. На одном торце к цилиндру прикрепляется крышка в форме блюдца, состоящая из 7 субъединиц кошаперонина GroES (10 кДа).
Субъединица GroEL состоит из трех доменов — обладающего АТФазной активностью экваториального домена, промежуточного домена и апикального домена, который связывает субъединицу кошаперонина GroES и может изменять угол своего наклона. Комплекс 14GroEL/7GroES7/7ATФ претерпевает цикл конформационных превращений, в ходе которых белок связывается, попадает в центральную полость шаперонина, а затем выходит в цитозоль (рис. 22). В центральной полости шаперо- нина частично развернутая молекула белка защищается от нежелательных взаимодействий при фолдинге или разворачивается с помощью молекулярного шаперона, что дает ей возможность снова принять правильную конформацию.
Рис. 22. Шаперонин. А, Б, В, Г — цикл конформационных превращений комплекса 14GroEL/ 7GroES/ 7АТФ; GroEL — трехдоменный молекулярный шаперон; GroES — кошаперонин.

Шаперонины архей. Как мы уже говорили, шаперонины архей не гомологичны шаперонинам бактерий и отличаются от них по архитектуре. Вместо шаперона
GroEL (Hsp60) у них имеется шаперон, гомологичный эукариотному шаперону ССТ.
Шаперонины архей изучены преимущественно у термофилов, например, Thermoplasma acidophilum (фила АII Euryarchaeota) и экстремальных термофилов — например, Sulfolobusshibae (фила AI Crenarchaeota) и Thermococcus profundus (фила All Euryarchaeota). Это дало основание называть их «термосомами», хотя аналогичные компартменты обнаружены у мезофилов, например, у экстремальных галофилов и метаногенов.
Термосомома представляет собой тороидальный комплекс, образованный двумя кольцами и состоящий из 16-18 идентичных или неидентичных субъединиц α/β. В частности, термосома S. shibae состоит из 16 идентичных субъединиц шаперона TF55 с молекулярной массой ~60 кДа. Она имеет тороидальную форму (высота 15 нм, диаметр 16 нм), ее центральная полость сообщается с цитозолем через отверстия диаметром 5 нм.
Как и бактериальный шаперонин, комплекс 16TF55/8ATФ проходит через цикл конформационных превращений, в ходе которых белок принимает правильную конформацию, побывав в центральной полости тороида.
8.3.10.3. Протеасомы
Наряду с шаперонинами, или микрокомпартментами, предназначенными для коррекции третичной структуры белка, существуют протеасомы, или микрокомпартменты, которые служат для ликвидации белков путем протеолиза. Сочетание фолдинга с протеолизом является одним из важнейших условий адаптации и выживания клеток в изменяющихся, особенно — в экстремальных условиях окружающей среды.
Существует несколько категорий белков, подлежащих устранению. Во-первых, это склонные к агрегации белки, третичная структура которых нарушена вследствие генной мутации, ошибки трансляции или теплового шока и не исправляется с помощью шаперонинов. Во-вторых, это белки, для которых характерна высокая скорость обновления (например, белковые факторы транскрипции, репарации, рестрикции-модификации, а также компоненты регуляторных каскадов или специфические белки, отвечающие за деление, клеточную дифференциацию и коллективное поведение). Втретьих, это субъединицы белковых комплексов, стехиометрия которых оказалась измененной. Вчетвертых, это белки, мобилизуемые как источник азота при голодании по этому элементу.
Протеолиз является мощным оружием, и чтобы его жертвой не оказались нормальные и необходимые белки, он держится под контролем. Наиболее эффективным способом является компартмен- тализация. Эукариоты используют для компартментализации протеаз специализированные мембранные органеллы (лизосомы).
Шире встречающимся способом компартментализации протеаз является их самосборка в протеасомы — ультрамикроскопические бочковидные аргегаты, во внутренней полости которых содержатся активные центры. Туда разрешен доступ только полностью развернутым белкам.
Развертывание осуществляется с помощью конститутивных или временно образующихся вспомогательных комплексов, которые обладают АТФазной активностью и называются анфолдазами (англ. unfold — разворачивать), или «обратными шаперонинами».
После развертывания белки расщепляются энергозависимыми протеазами. Они относятся к четырем низкогомологичным суперсемействам — Clp, Lon, FtsH и 20S-протеасомам. СIр-протеазы ClpAP и ClpXP представляют собой комплекс АТФазы (СIрА или СIрХ) и «сериновой», т. е. имеющей каталитический домен с остатками Ser протеазы (СIрР). Lon-протеазы содержат АТФазный домен и сериново-протеазный домен внутри одного полипептида. FtsH-протеазы содержат АТФазный домен и Zn2+-зависимый протеазный домен также внутри одного и того же полипептида. Некоторые из них индуцируются при стрессе или тепловом шоке.
Перечисленные протеазы — это мультикаталитические комплексы, образующие путем самосборки компартмент, в котором протеолитические активные центры изолируются от содержимого клетки. Предварительно развернутые (не)нормальные белки входят в эти микрокомпартменты
сквозь узкие отверстия — их ширина примерно равна диаметру α-спирали. С помощью нескольких активных центров они гидролизуются до олигопептидов, причем до окончания процесса промежуточные продукты в цитозоль не выходят.
Важнейшими энергозависимыми протеазами являются 20S-протеасомы.
20S-протеасомы. Генеральные компартменты этого типа образуют гомогенную популяцию субмикроскопических частиц, имеющих размер 12 x 15 нм и молекулярную массу 700-750 кДа. В эукариотной клетке они локализуются в ядре или цитоплазме, где ассоциированы с эндоплазматическим ретикулумом и цитоскелетом. В прокариотной клетке они находятся в цитоплазме или непрочно связаны с внутренней поверхностью СМ.
20S-протеасомы распространены у всех эукариотов и архей. У бактерий они встречаются довольно редко — только у отдельных представителей филы BXIV Actinobacteria (рода Frankia, Mycobacterium, Rhodococcus и Streptomyces).
Основное различие между прокариотными и эукариотными 20S-протеасомами заключается в степени их сложности. Прокариотные протеасомы обычно содержат несколько идентичных копий субъединиц α и β. Последние взаимно гомологичны и состоят из пяти центральных складчатых β-цепей, фланкированных по обоим концам молекулы α-спиралями. По сравнению с субъединицей а, субъединица β имеет более короткий N-концевой фрагмент.
В свою очередь, эукариотные протеасомы содержат по 7 гомологичных вариантов субъединиц α и β (молекулярной массой 22-35 кДа). У позвоночных структура протеасомы усложняется тем, что несколько конститутивных субъединиц β замещаются индуцибельными гомологами.
Каталитическую активность проявляют только субъединицы β, и она определяется единственным аминокислотным остатком — N-концевым остатком Thr.
Из субъединиц α и β путем самосборки образуются гептамерные кольца α7 и β7 (у прокариотов) и α1-7 и β1-7 (у эукариотов). В обоих случаях два сложенных вместе β-кольца фланкированы двумя а-кольцами, в результате чего образуется бочковидный комплекс из четырех колец, высота которого 15 нм, а диаметр 12 нм (рис. 23). Через него проходит сквозной канал, расширяющийся в центральную полость, и два фланкирующих ее «преддверья» размером 4x5 нм. Центральная полость размером 3,8 х 5,5 нм содержит каталитические центры и образована двумя β-кольцами. Оба преддверья, функции которых неизвестны, образованы одним α- кольцом и одним β-кольцом. «Субстратные» белки проникают в преддверья через входные отверстия в α-кольцах диаметром 1,3 нм. Из преддверий они попадают в центральную полость через отверстия в β-кольцах, диаметр которых составляет 2,2 нм.
Рис. 23. Протеасома. α и β —субъединицы; А1 и А2 — «преддверья»; С — центральная полость. Черными кружками обозначены каталитические центры.

В результате неспецифического протеолиза субстратный белок расщепляется на олигопетиды из 4-10 аминокислотных остатков, которые освобождаются в цитозоль и там подвергаются дальнейшему расщеплению. В центральной полости может разместиться только одна молекула субстратного белка, и освободившееся после ее окончательного расщепления место занимает следующая молекула.
Ширина входного отверстия протеасомы приближается к диаметру полипептидной цепи. Поэтому через него могут пройти только полностью развернутые белки, что предварительно осуществляется с помощью вспомогательных комплексов.
Обязательным компонентом этих комплексов служит АТФ-аза. В случае эукариотов и бактерий это АТФаза из суперсемейства ААА (от англ. ATPases associated with various cellular activities), a в случае архей — АТФаза из суперсемейства PAN (от англ, protéasome activating nucleotidase). При временном (у прокариотов) или постоянном (у эукариотов) объединении с АТФазой 208-протеасома становится функционально завершенной структурой.
У эукариотов 20S-протеасомы становятся частью более сложного компартмен- та — 26S-протеасомы.
26S-протеасомы универсальны для эукариотов и необходимы для их жизнедеятельности. Они имеют молекулярную массу ~2500 кДа и содержат вспомогательные комплексы в виде двух асимметричных 19S-кэпов, фланкирующих бочковидную 208-структуру. Кэпы состоят из 15 разных субъединиц с молекулярной массой 25-110 кДа. Одни из них распознают субстратный белок, другие обладают АТФазной активностью и окончательно разворачивают субстратный белок, третьи транслоцируют его во внутреннюю полость протеасомы и презентируют его активным центрам.
Получение белком «черной метки» для протеолитической деградации обычно осуществляется путем убиквитирования, т. е. АТФ-зависимого ковалентного связывания с цитозольным белком убиквитином (англ. ubiquitin, Ub; 8,6 кДа).
Таким образом, у эукариотов АТФ-зависимый протеолиз включает в себя две стадии: а) распознавание неправильно свернутого белка убиквитином и его мечение; б) расщепление меченного убиквитином субстратного белка в 26S-протеасоме.
Между 20S-кором и кэпами расположены кольцеобразные переходники. Они состоят из шести субъединиц, принадлежащих к суперсемейству ААА-АТФаз.
Поскольку четыре из семи β-субъединиц не имеют N-концевого остатка Тhr, каталитической активностью обладают не все 14 β-субъединиц, а только шесть из них (β1, β1', β2, β2', β5 и β5').
В сборке эукариотной протеасомы принимают участие молекулярные шапероны, например, Нсs73. Напротив, прокариотные протеасомы образуются путем самосборки, и для их созревания молекулярные шапероны не требуются.
20S-протеасомы бактерий. Если эукариоты используют протеасомы в любых условиях, то для бактерий их присутствие необязательно (например, мутанты Mycobacteriumsmegmatis с делегированными генами протеасом не отличаются от штаммов дикого типа). Они есть не у всех бактерий и обычно экспрессируются в экстремальных условиях.
20S-протеасомы не содержат 19S-кэпов (в отличие от 26S-протеасом). У них также есть комплексы, распознающие и развертывающие субстратные белки, но стационарной структуры они не образуют.
Как уже указывалось, бактериальные протеасомы состоят из однотипных субъединиц оси β. Исключение представляет Rhodococcus erythropolis N186/21, у которого по два типа этих субъединиц.
Бочковидный комплекс из четырех колец содержит 14 активных протеолитических сайтов.
Интересно, что по строению бактериальные 20S-протеасомы напоминают шаперонины. Оба микрокомпартмента имеют цилиндрическую форму, сходные размеры и состоят из гептамерных колец, образующих центральную полость (см. раздел 8.3.10.2). Однако между шаперонинами и протеасомами есть существенное различие. Входное отверстие шаперонина имеет диаметр 4,5 нм, благодаря чему в центральную полость проникают частично развернутые белки, и там их структура корректируется. Протеасома имеет более узкое входное отверстие (1,3 нм), поэтому в центральную полость попадают только полностью развернутые белки, где подвергаются протеолитическому расщеплению.
В развертывании субстратных белков могут участвовать молекулярные шапероны DnaK/DnaJ/Hsp70 и GroEL/GroES.
«Упрощенные» протеасомы бактерий. Большинство бактерий содержит более просто устроенные протеолитические микрокомпартменты, примером которых является протеасома HslV (сокр. англ. heat-shock locus V). Она состоит из двух колец, по шесть субъединиц в каждом. Эти кольца взаимодействуют с АТФазой из семейства Clp (HslU). Отверстия на противоположных сторонах образующегося цилиндра имеют диаметр ~0,7 нм.
HslV-протеасомы и 20S-протеасомы вместе не встречаются. Некоторые бактерии протеасом вообще не образуют, например, они отсутствуют у Synechocystis sp. РСС 6803 (филаВХ Cyanobacteria), Mycoplasma genitalium, М. pneumoniae (фила ВХШ Firmicutes) и Chlamydia trachomatis (фила BXVI Chlamydiae).
20S-протеасомы архей. Если для эукариотов протеасомы «универсальны и обязательны», для бактерий «неуниверсальны и необязательны», то для архей они «универсальны, но необязательны».
Состав и архитектура протеасом у архей в настоящее время досконально изучены, в том числе с помощью рентгеноструктурного анализа с уровнем разрешения 3,5 À. Установлено, что они почти не отличаются от бактериальных 20S-протеасом (см. рис. 23).
Как и бактериальные протеасомы, протеасомы архей обычно состоят из субъединиц α и β только одного типа. Однако у нескольких видов Aeropyrum (фила AI Crenarchaeota), а также Haloferax и Halococcus (фила АII Euryarchaeota) они бывают 2-3 типов.
В отличие от бактерий, архей не имеют СIр-протеаз, Lon-протеаз и FtsH-протеаз, которые агрегируют с образованием «упрощенных» протеасом. Поэтому за энергозависимую деградацию белков у них отвечают только 20S-протеасомы.
Естественно, что протеасомы архей так же, как протеасомы эукариотов и бактерий, участвуют в клеточном ответе на неблагоприятные условия окружающей среды.
8.3.11. Специализированные компартменты
В цитоплазматическом компартменте прокариотной клетки может находиться несколько типов специализированных компартментов. Они различаются способом, которым обеспечивается их граница, своим содержимым и функциями. Одни из них встречаются у представителей обоих прокариотных доменов, другие бывают только у бактерий. Некоторые имеют очень широкое распространение; другие крайне редки, а иногда даже уникальны.
8.3.11.1. Вакуоли
В соответствии с определением, которое дал прокариотам Роджер Стэниер (см. раздел 2.3), одним из цитологических признаков, чем они принципиально отличаются от эукариотов, служит отсутствие вакуолей — микрокомпартментов, окруженных унитарной мембраной.
Классический термин «вакуоль» (от лат. vacuum — пустое пространство) используется в цитологии ядерных организмов, хотя им обозначают структуры, которые имеют разный состав и выполняют разные функции. Чаще всего вакуолями называют микрокомпартменты, которые находятся в цитоплазме, окружены унитарной мембраной, или тонопластом и заполнены гидрофильным жидким содержимым. У протистов и растений вакуоли служат для поддержания осмотического баланса с окружающей средой путем компартментализации воды с ее последующим удалением, а также для хранения раствора гидрофильных метаболитов, или клеточного сока.
Тонопласт происходит от irans-мембран аппарата Гольджи. Во многих отношениях он напоминает лизосомальную мембрану, хотя это две разные «стволовые линии» онтогенеза эукариотных ICM. В обоих случаях за счет ограничивающей мембраны обеспечивается градиент концентрации катионов, в том числе Н+, и внутри обоих компартментов изолируются гидролитические ферменты. Однако вакуоли служат преимущественно для хранения метаболитов, а лизосомы — для деструкции макромолекул или для процессинга секретируемого материала.
Таким образом, вакуоли представляют собой структурно обособленный и функционально специализированный участок ICM эукариотной клетки.
Мы уже говорили о том, что IСМ бактерий принципиально отличаются от IСМ эукариотов. Хотя вопрос о структурной обособленности IСМ от СМ до конца не выяснен, известно, что эти два участка мембранной системы различаются по молекулярному составу. Установлено также, что у бактерий «узкой специализацией» ГСМ служит ассимиляция энергии (см. раздел 8.3.5).
Модули, из которых состоят ГСМ, в некоторых случаях обнаруживают внешнее сходство с вакуолями, особенно когда они раздуваются, однако это не дает оснований считать их специализированными органеллами.
Истинные вакуоли у бактерий исключительно редки, и сведения о них носят в основном описательный характер.
Загадкой для цитологов стала нитратная вакуоль морских хемолитотрофов Beggiatoa, Thiomargarita и Thioploca, которая была впервые обнаружена в середине 1980-х годов (см. раздел 8.1.3.4). Внутри этого ограниченного унитарной мембраной компартмента, занимающего основную часть внутриклеточного объема, накапливаются анионы NO3- до конечной концентрации ~0,8 М. Среди представителей рода Thioploca есть штаммы с широкими и узкими клетками: в первом случае образуется одна крупная нитратная вакуоль, во втором — множество мелких нитратных вакуолей.
Упомянутые бактерии используют нитрат в качестве терминального акцептора электронов при анаэробном окислении сульфида, в результате чего NO3- восстанавливается до NH3или N2.
Недавно у Beggiatoa-подобных бактерий были обнаружены истинные вакуоли, внутри которых запасается не нитрат, а другой терминальный акцептор электронов — молекулярный кислород.
Механизм образования нитратных и кислородных вакуолей неизвестен; дело усугубляется тем, что бактерии с такими органеллами относятся к числу некультивируемых форм.
Еще одним примером вакуоли у бактерий является анаммоксосома, открытая в конце 1990-х годов. Эта уникальная органелла запасает энергию путем анаэробного окисления NH3, используя в качестве акцептора электронов анионы NO2- (см. II том учебника). Данным свойством обладает ряд представителей филы BXV Planctomycetes (Candidatus «Brocadiaanammoxidans», Candidatus «Scalindua sorokinii» и Candidatus «Kuenenia stuttgartiensis»).
Анаммоксосома занимает до 30% внутреннего объема клетки (рис. 24). Она окружена унитарной мембраной, состоящей из уникальных липидов — «ледеранов» (англ. ladder — лестница).
Рис. 24- Анаммоксосома в клетке бактерии-планктомицета. СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; S — поверхностный белковый слой; МА — мембрана анаммоксосомы; С — хромосома.
Жирные «хвосты» ледеранов (рис. 25) представляют собой С20-спирты или С20- кислоты. Они состоят из 8-углеродной линейной цепи, с которой связана «лестница» из пяти циклобутановых остатков или из трех циклобутановых остатков и одного циклогексанового остатка. Такие гидрофобные радикалы образуют простой эфир с метанолом или глицеролом, либо смешанный (простой и сложный) эфир с глицеролом.
Рис. 25. Липиды из группы ледеранов.
А — сложный эфир метанола и С20-жирной кислоты, содержащей пять агрегированных циклобутановых колец; Б — простой моноэфир глицерола и С20-жирной кислоты, содержащей три циклобутановых кольца, агрегированных с одним циклогексановым кольцом; В — эфир смешанного типа; Г — способ укладки ледеранов в липидном слое мембраны анаммоксосомы.
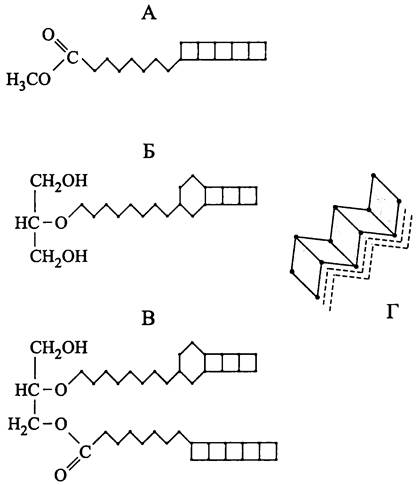
Напомним, что липиды на основе простых эфиров глицерина (универсальные для архей) у бактерий встречаются исключительно редко. Кроме рассматриваемого случая, они найдены только у некоторых термофилов и сульфатредукторов (см. раздел 8.3.3). У эукариотов их не бывает никогда.
Плотно упакованные ледераны создают непроницаемый барьер для токсичного гидразина (NH2-NH2J, образующегося внутри анаммоксосомы при окислении аммиака. При окислении гидразина анаэробной дыхательной цепью, состоящей из гидразиндегидрогеназы (англ. hydrazine oxidizing enzyme, HZO) и нитритредуктазы (англ. nitrite reducing enzyme, NR), на мембране анаммоксосомы создается протонный градиент.
В мембране анаммоксосомы содержится АТФ-синтаза, которая осуществляет фосфорилирование АДФ при движении протонов по градиенту их концентрации, т. е. из аммоксосомы в цитоплазму. Таким образом, анаммоксосома является уникальным дыхательным компартментом — функциональным аналогом симбиотической органеллы ядерной клетки, митохондрии.
Изучение анаммоксосомы только начинается, и механизм ее биогенеза пока еще неизвестен.
8.3.11.2. Газовые везикулы
Эти специализированные микрокомпартменты ограничены неунитарной мембраной. Они содержат газовую фазу, что придает прокариотной клетке повышенную плавучесть. Газовые везикулы широко распространены у планктонных форм и описаны примерно у 150 видов бактерий и архей. Особенно часто они встречаются у цианобактерий.
Газовые везикулы имеют стандартную морфологию и представляют собой пустотелые цилиндры с полыми конусами на обоих концах. У разных прокариотов они различаются по соотношению диаметра (50-200 нм) и длины (100-1200 нм). Стенка газовой везикулы имеет толщину ~2 нм и представляет собой однослойную мембрану, которая у разных видов состоит из взаимно гомологичных белков.
Газовые везикулы лежат в цитоплазме поодиночке или образуют сотовидные агрегаты (их неправильно называют газовыми «вакуолями»).
Газовая везикула — это инертная структура. Она заполнена атмосферным воздухом, который диффундирует в нее через клеточную оболочку, цитоплазму и белковую мембрану. В модельных экспериментах атмосферный воздух можно заменить различными газами (Аr, СН4, СО, СО2, Н2, N2, О2 или водяным паром). В предельном варианте внутри газовой везикулы может быть вакуум.
Мембрана газовой везикулы состоит из двух гидрофобных белков. Один из них уложен паракристаллическим монослоем вдоль кольцевых ребер, образующих цилиндрический баллон с коническими концами (рис. 26). Молекулы другого белка лежат поперек 4-5 ребер и стабилизируют органеллу снаружи.
Рис. 26. Газовая везикула. GvрА — главный белок неунитарной мембраны, образующей газовую везикулу.
Газы беспрепятственно диффундируют сквозь поры гидрофобной мембраны, которые имеют диаметр ~0,6 нм. В результате содержимое газовой везикулы по общему и парциальным давлениям не отличается от газовой фазы над жидкой средой, в которой находится бактерия. Вода из цитоплазмы удерживается силами поверхностного натяжения и вместе с гидрофильными молекулами внутрь газовой везикулы не проникает.
Мембрана газовой везикулы нерастяжима и механически прочна, но, когда давление на нее становится выше критического, газовая везикула сплющивается и расправиться уже не может.
В биогенезе газовой везикулы участвуют до 15 хромосомных или плазмидных gvp-генов (от англ. gas vesicle protein). Молекулярная масса их продуктов составляет 7-50 кДа. Большинство белков не идентифицировано; они могут соответствовать минорным компонентам газовой везикулы, а также участвовать в ее сборке или выполнять регуляторную роль (например, продукты генов gvpF-L).
Ген gvpA кодирует главный белок (7,5 кДа), на долю которого приходится до 90% твердого материала органеллы. Он состоит из двух антипараллельных (3-цепей; около 230 молекул образуют ребро шириной 4-5 нм (рис. 26). До сих пор неизвестно, являются ли ребра замкнутыми кольцами или это отдельные витки туго скрученной спирали. Исключительно высокая гидрофобность белка GvpA обеспечивает самосборку и гидрофобные свойства внутренней поверхности мембраны газовой везикулы.
Продукт гена gvpC имеет молекулярную массу 22 кДа и содержит повторяющийся 4-5 раз α-спиральный мотив из 33 аминокислотных остатков. Предполагается, что задача этого белка — стягивать ребра газовой везикулы, хотя неизвестно, как он ориентирован по отношению к ее продольной оси.
При биогенезе газовой везикулы вначале образуется биконическая структура, которая растет в длину и одновременно с этим в нее диффундирует газ из цитоплазмы.
Плотность бактерий обычно выше плотности воды, поскольку они богаты белками (1,33 г • см-3), углеводами (1,55 г • см-3) и нуклеиновыми кислотами (1,66 г • см-3). Благодаря образованию множества газовых везикул (плотностью 0,06-0,2 г • см-3) плотность бактерий снижается до 0,975 г • см-3 и становится ниже плотности воды (0,998 г • см-3 при 20°С). В результате они всплывают к поверхности водоема. Такой тип подвижности называется флотацией (см. раздел 8.6.2). Фототрофам он обеспечивает более выгодный световой режим, аэробам — лучшее снабжение кислородом. Кроме того, газовые везикулы отражают свет и поэтому защищают клетки от фотоповреждения.
Внешнее давление на мембрану газовой везикулы складывается из атмосферного давления или из давления экспериментальной газовой фазы над жидкой средой (PG),гидростатического давления (Рн), тургорногo давления (Рт) и силы поверхностного натяжения (Ps)- Газовая фаза внутри везикулы находится в равновесии с атмосферным воздухом или с экспериментальной газовой фазой над жидкой средой, поэтому контр-давление равно PG. В итоге ригидность мембраны газовой везикулы компенсирует Рн +Рт+ Ps (в зависимости от радиуса, она выдерживает критическую разность давлений в 0,1-3,5 мПа). Поскольку газ перераспределяется между полостью органеллы и цитоплазмой довольно медленно, увеличение гидростатического давления при быстром погружении бактерии или увеличение тургорного давления из-за накопления осмотически активных веществ вызывают коллапс газовых везикул, и клетка начинает тонуть. Тургорное давление, в частности, возрастает при биосинтезе углеводов. При высоком уровне автотрофной ассимиляции углерода в клетке накапливаются полигликозидные гранулы, играющие роль балласта. Напротив, при низком уровне автотрофной ассимиляции углерода образуются новые газовые везикулы, и клетка всплывают к поверхности.
Широта распространения газовых везикул у прокариотов и взаимная гомология Gvр-белков говорят в пользу их древнего происхождения. Трудно допустить, что прокариоты специально «изобрели» газовые везикулы для флотационной подвижности. Скорее всего, это свойство было приобретено вторично после мутационного
изменения белков, выполнявших иные функции. Уже давно подмечено сходство газовых везикул с белковой оболочкой, или капсидом вириона (интересно, что белок Gvр А на 50% гомологичен гликопротеину шипов капсида у вируса везикулярного стоматита). И все же «вирусное» происхождение этих специализированных компартментов маловероятно.
8.3.11.3. Полярные мембраноподобные органеллы
Под таким названием выступают загадочные микрокомпартменты, в разное время выявленные у разных прокариотов.
В случае метаногенных архей Methanococcus jannaschii, М. vannielii и М. voltae полярные мембраноподобные органеллы выглядят как сдвоенные ламеллы длиной до 0,5 мкм. Они расположены в основании жгутика, параллельно СМ на расстоянии от нее около 25 нм. Промежуток между дистальной ламеллой и СМ заполнен диффузным поперечно исчерченным материалом.
Поскольку срез такой ламеллы имеет двухслойный контур, их назвали «мембраноподобными». Это едва ли оправданно, поскольку настоящие мембраны всегда везикулизированы, т. е. замыкаются в сферу, а ламеллы мембраноподобных органелл свободны по концам. Аналогичные структуры описаны у экстремально галофильного археота Halobacteriumsalinarium, а также у грамотрицательных бактерий с полярным типом жгутикования — Campylobacter jejuni и Spirillum serpens.
Конкретные выводы о функциональном значении полярной органеллы делать еще рано, хотя, судя по ее локализации, она способствует работе жгутиков.
С одной стороны, полярная органелла может быть интегральным компонентом жгутикового аппарата. С помощью цитохимических методов показано, что она проявляет АТФазную активность, хотя непосредственным источником энергии для вращения жгутика служит не АТФ, a Pmf или Smf (см. разделы 8.6.2 и II том учебника).
С другой стороны, полярная органелла может быть частью эндоскелета или энзоскелета (см. разделы 8.3.9.1 и 8.3.9.3), и тогда ее функции должны быть связаны с поддержкой базального тела или с процессингом субъединиц жгутика и их векторным транспортом.
8.3.11.4. Полифосфатные, полиглюкозидные и цианофициновые гранулы
Специализированные микрокомпартменты, перечисленные в заглавии данного подраздела, принадлежат к числу классических включений (см. раздел 8.2).
Отдавая должное термину «включение», который используется и по отношению к эукариотам, нельзя не заметить, что он придает соответствующим структурам нежелательный смысловой оттенок — функционально инертного и даже чуждого клетке образования. Для включений, которые относительно редко встречаются в клетках эукариотов (примером служат кристаллы оксалата кальция у растений), это в чем-то справедливо. Напротив, когда мы говорим о включениях у прокариотов, такая интерпретация может принизить и даже исказить роль этих микрокомпартментов. Ведь им отводится значительная часть пространства клетки, они проявляют высокую физиологическую активность и чаще всего полифункциональны.
Включения хранят резервные вещества в осмотически инертной полимерной форме и представляют собой динамическое депо, где клетка запасает три метаболических ресурса — энергию, конструктивный субстрат или восстановитель (см. II том учебника). Объем этого депо зависит от стадии развития кульуры.
Рост можно определить, как координированную репликацию клеточных структур, которая, в конечном счете, приводит к увеличению биомассы. Часто, но далеко не всегда, рост предшествует делению. В общем случае рост бывает сопряженным и несопряженным.
«Сопряженный» рост (англ. balanced growth) является результатом одинакового темпа репликации всех субклеточных структур, в результате чего фракционный состав биомассы не изменяется. Например, за период ∆t количество ДНК, рибосом, мембранных липидов, материала ригидного слоя, включений и т. д. возрастает в одинаковое число раз (хn).
«Несопряженный» рост (англ. unbalanced growth) приводит к структурной диспропорции, так как разные компоненты клетки за период At реплицируются с разной скоростью (xn1, xn2, xn3 и т. д.). Обычно это связано с метаболическим стрессом при неравномерном истощении питательных субстратов — например, клетке не хватает экзогенного азота или фосфора на фоне относительного избытка экзогенного углерода. Несопряженный рост может также быть результатом попадания в новые трофические условия (англ. nutritional shift) при резкой смене ниши или при пересеве на другую питательную среду.
Полифосфатные гранулы. Это цитоплазматические микрокомпартменты, ограниченные твердой фазой. Они состоят из конденсированных неорганических фосфатов, или просто «полифосфатов» (Ф)n — неразветвленного полимера, состоящего из 100-1000 остатков неорганического фосфата (Ф), которые соединены фосфоэфирными связями (рис. 27).
Рис. 27. Полифосфат и воздействующие на него ферменты. 1 — эндополифосфатаза; 2 — экзополифосфатаза; 3-5 — АМФ-фосфотрансфераза, полифосфаткиназа и полифосфат-глюкокиназа; 6 — гуанозинтетрафосфат: пирофосфат-трансфераза.
Полифосфаты часто называют «молекулярными ископаемыми». По-видимому, они существовали еще на пребиотической стадии эволюции Земли, поскольку спонтанно образуются при дегидратации ортофосфата в вулканических конденсатах и глубоководных газовых выбросах. В современной биосфере способностью к биосинтезу полифосфатов обладает большинство прокариотов, а также некоторые протисты, грибы, водоросли и даже млекопитающие.
Будучи полианионами, полифосфаты связывают катионные красители, максимум поглощения которых при этом сдвигается в коротковолновую область. Поэтому полифосфатные гранулы часто называют «метахроматиновыми». Устаревший термин «волютиновые» гранулы происходит от видового эпитета бактерии Spirillum, volutans.
Под электронным микроскопом полифосфаты выглядят как резко контрастные кристаллоидные сферы диаметром от 50 нм до >1 мкм. Они расположены в центральной области клетки и быстро испаряются в электронном луче, оставляя после себя пустое место. Количество полифосфатных гранул при избытке фосфора в среде относительно постоянно и быстро уменьшается вплоть до полного исчезновения при фосфорном голодании.
Биосинтез полифосфатов. Полифосфаты в основном синтезируются с помощью полифосфаткиназы из концевой![]() фосфатной группы АТФ:
фосфатной группы АТФ:
nАТФ (Ф)n + nАДФ.
Сначала фосфорилируется His в активном центре, и он служит затравкой для образования полимерной цепи, которая у Е. coli имеет длину до 800 остатков. Фермент иммобилизован на СМ и представляет собой тетрамер из субъединиц молекулярной массой 80 кДа.
Функции полифосфатов. Полифосфаты полифункциональны. Они служат резервуаром энергии и фосфора, являются эндогенным регулятором метаболизма, а также хелатируют катионы металлов. В комплексе с катионами Са2+ полифосфаты локально изменяют фазовое состояние липидов СМ, что обеспечивает компетентность бактерий при генетической трансформации. Иногда из них состоит капсула (см. раздел 8.5.4.1).
При недостатке АТФ полифосфаты выполняют роль фосфагена (см. II том учебника). В этом случае концевая фосфорильная группа переносится на АДФ (с помощью полифосфаткиназы) или на АМФ (с помощью АМФ-фосфотрансферазы):
(Ф)n + АДФ —> (Ф)n-1 + АТФ,
(Ф)n-1 + АМФ —> (Ф)n-2 + АДФ, а затем АДФ диспропорционируется аденилаткиназой:
2АДФ —> АТФ + АМФ.
Помимо этого, концевая фосфорильная группа используется для активации глюкозы с помощью полифосфат-глюкокиназы:
(Ф)n + глюкоза —> (Ф)n-1 + глюкозо-6-фосфат.
Полифосфаты подвергаются «холостому» расщеплению под воздействием экзо- и эндополифосфатаз (рис. 27). Ортофосфат, образующийся с помощью иммобилизованных на СМ димерных экзополифосфатаз (58 кДа), служит источником фосфора для биосинтеза нуклеиновых кислот и фосфолипидов.
Участие полифосфатов в регуляторных процессах заключается прежде всего в том, что с опероном ррк, который кодирует полифосфаткиназу и экзополифосфатазу Е. coli, взаимодействует pho-регулон, регулирующий активность более 30 генов фосфорного метаболизма. Кроме того, с помощью гуанозинтетрафосфат-пирофосфат- трансферазы от полифосфата на ГДФ переносится концевой фрагмент, состоящий из двух фосфорильных групп, в результате чего образуется гуанозинтетрафосфат (англ. guanosine tetraphosphate, ppGpp):
(Ф)n + ГДФ —> (Ф)n-2 + ppGpp.
Это соединение играет роль сигнала тревоги, или «алармона» (англ. alarm — тревога). В условиях стресса, особенно при голодании по аминокислотам или неорганическому фосфору, ppGpp обеспечивает «строгий» ответ (англ, stringent response) — репрессирует одни и активирует другие гены. Это способствует выживанию бактерий, в частности благодаря дифференциации покоящихся клеток.
Очень важна способность полифосфатов хелатировать катионы металлов (Са2+, Mg2+, Мn2+ др.). Например, не имеющая супероксиддисмутазы молочнокислая бактерия Lactobacillus plantarum таким способом накапливает до 30 мМ марганца, с помощью которого устраняется токсичный супероксид (Мn2+ + О2- • —> 2Н+ —> Н2О2 + Мn3+; Мn3+ + О2- —> Mn2+ + О2). Для многих прокариотов поли фосфаты служат крайним средством для обезвреживания проникших в клетку токсичных катионов Cd2+, Cu2+, Fe2+ или Zn2+.
Практическое использование бактерий, активно синтезирующих полифосфаты, позволит удалять растворенный фосфор из сточных вод и тем самым предотвращать эвтрофикацию. Кроме того, выделенные из бактерий полифосфаты могли бы применяться в пищевой индустрии, а также служить заменителем асбеста при изготовлении негорючих тканей.
Полиглюкозидные гранулы (α-гранулы). Эти твердофазные цитоплазматические микрокомпартменты состоят из гидрофильного нерастворимого полиглю- козида молекулярной массой до 200 кДа.
Способностью синтезировать полиглюкозиды обладают отдельные представители всех трех глобальных доменов. Среди прокариотов это многие бактерии из разных фил, а также некоторые археи, например Methanococcus thermolithotrophicus (фила АII Euryarchaeota) и Sulfolobus acidocaldarius (фила AI Crenarchaeota).
Полиглюкозиды состоят из остатков глюкозы, соединенных связями α1 —>4 (неразветвленная цепь) или связями α1—>4 и связями α1—>6 (разветвленная цепь). Бактерии и археи обычно синтезируют только разветвленный полиглюкозид — аналог животного гликогена. Значительно реже у бактерий встречается только неразветвленный полиглюкозид - гранулоза, которую в количестве до 60% сухого веса накапливают перед началом спорообразования многие виды рода Clostridium. Единственным известным примером, когда у прокариотов образуется аналог растительного крахмала (тривиальное название смеси неразветвленного полиглюкозида амилозы и разветвленного полиглюкозида амилопектина), служит цианобактерия Prochloron didemni.
Прокариотный гликоген постоянно присутствует в клетке в небольшом количестве, а его накопление происходит при неблагоприятных условиях (например, при субоптимальном pH) или, когда рост прекращается из-за недостатка экзогенного азота, серы, фосфора или железа на фоне относительного избытка экзогенного углерода.
Электроноплотные полиглюкозидные гранулы равномерно распределены в цитоплазме и имеют форму гомогенных или структурированных палочек диаметром 30 нм и длиной 50-300 нм.
При биосинтезе прокариотного гликогена донором глюкозильного мономера служит АДФ-глюкоза, которая образуется из глюкозо-1-фосфата под действием растворимой индуцибельной АДФ-глюкозо-пирофосфорилазы (50 кДа; ген glgC):
АТФ + глюкозо-1-фосфат —> АДФ-глюкоза + ФФ.
Затем гликогенсинтаза (40 кДа; ген glgA), иммобилизованная на затравке α1 —> 4- глюкана, переносит на нее глюкозильный мономер:
АДФ-глюкоза + α1 —> 4-глюкан —> α1 —> 4-глюкозилглюкан + АДФ.
В завершение всего ~10% глюкозильных мономеров соединяются связями α1 —> 6. Катализатором реакции служит глюкозилтрансфераза (85 кДа; ген glgB), которая играет роль «разветвляющего» фермента (англ. branching enzyme).
При биосинтезе гранулозы, которая накапливается в клетках представителей рода Clostridium, неразветвленная полиглюкозидная цепь может образовываться из дисахаридов — мальтозы или сахарозы — соответственно с помощью амиломальтазы и амилосахаразы:
мальтоза + (α1-4-глюкозил)n —> глюкоза + (α1-4-глюкозил)n+1, сахароза + (а1-4-глюкозил)n —> фруктоза + (α1-4-глюкозил)n+1.
Прокариотный гликоген используется в результате совместного действия индуцибельной α-амилазы (гидролизует неразветвленный участок цепи на декстрины), пуллуланазы (атакует точки ветвления) и глюкоамилазы (образует мономеры глюкозы).
Цианофициновые гранулы. «Полипептид цианофициновых гранул» (англ. cyanophycin granule polypeptide, CGP), или просто «цианофицин» — это уникальный полипептид, который образуют только некоторые цианобактерии. Он синтезируется из отдельных мономеров нерибосомным АТФ-зависимым путем с помощью транспептидазного ферментного комплекса. Готовый полимер представляет собой линейную цепь, состоящую из остатков L-аспарагиновой кислоты, β-карбоксильные группы которых связывают остатки L-аргинина через их α-аминогруппы:
Цианофицин имеет молекулярную массу 25-125 кДа и находится в нерастворимом состоянии в цитоплазме, образуя округлые негомогенные гранулы разного размера.
В условиях стресса (слабое освещение, низкая температура, голодание по сере, фосфору и т. д.) или при избытке внешнего источника азота содержание цианофицина может достигать 20% клеточной массы. Он используется в качестве источника азота и углерода при сбалансированном росте. Сборка цианофицина осуществляется на затравке с помощью димерной цианофицинсинтазы (130 кДа; ген cphA). Для действия этого фермента необходимы оба мономера, АТФ, катионы К+ и Mg2+, а также вспомогательный физиологический реагент, содержащий SH-группы. Деполимеризует цианофицин мономерная экзопептидаза — циафициназа (30 кДа; ген cphB), которая в качестве конечного продукта образует дипептид Asp-Arg.
Устойчивый к протеазам цианофицин интересен для биотехнологии, поскольку из него легко получить полиаспарагиновую кислоту, которая может быть заменителем технических полиакрилатов. Поскольку цианобактерии растут довольно медленно, для производства цианофицина выгоднее использовать рекомбинантные штаммы Е. coli.
8.3.11.5. Цитоплазматические микрокристаллы
Случаи, когда в цитоплазматическом компартменте бактерий встречаются не ограниченные мембраной гидрофильные кристаллы, довольно редки. Будучи по сути балластными или запасными включениями, они играют важную роль в жизни отдельной клетки или популяции в целом.
Микрокристаллы кальцита. Уникальным примером бактерии, образующей цитоплазматические кристаллы кальцита (СаСОз), является гигантская серная бактерия Achromatiumoxaliferum. Включения занимают больше 70% ее объема, составляющего 3 • 104 мкм3. По-видимому, благодаря этому клетка увеличивает количественное соотношение между площадью своей поверхности и объемом цитоплазмы, что частично снимает ограничение, которое «гигантский» размер накладывает на интенсивность транспортных процессов (см. раздел 8.1.3).
Микрокристаллы селена. Некоторые анаэробные бактерии, например, Sulfо spirillum bamesii (класс «Epsilonproteobacteria») содержат в цитоплазме округлые кристаллы элементного селена диаметром до 200 нм. Они образуются в результате восстановления аниона селенита SеО32-, что позволяет обезвредить это ядовитое соединение, которое при высокой концентрации в окружающей среде неспецифически импортируется сульфатпермеазой или нитратпермеазой. Таким образом, кристаллы селена — это крайне специфичные балластные включения, образование которых служит своеобразной «платой» за сохранение клетки в жизнеспособном состоянии.
Микрокристаллы теллура. Представители семи видов квази-фототрофных бактерий из класса «Alphaproteobacteria» (см. II том учебника) — Erythrobacter litoralis, Erythromicrobium ezovicum, E. hydrolyticum, E. ramosum, Erythromonas ursincola, Roseococcus thiosulfatophilus и Sandaracinobacter Sibiriens — обладают способностью восстанавливать анион теллурита ТеО32- до элементного теллура. Игольчатые кристаллы теллура, очень крупные (30 x 500 нм) у одних и мелкие (8 x 50 нм) у других видов, образуются в цитоплазме и в некоторых случаях занимают до 30% объема клетки.
Как и в случае восстановления селенита в селен, восстановление теллурита в относительно инертный теллур служит эффективным способом детоксикации этого ядовитого поллютанта. Данное свойство бактерий может быть использовано в целях ремедиации, а также для концентрирования теллура для нужд металлургии.
Микрокристаллы железа. Цитоплазматические микрокристаллы железа обычно окружены унитарной мембраной и вместе с ней образуют органеллы, способствующие пассивной ориентацией бактериальной клетки во внешнем магнитном поле (см. раздел 8.3.11.6). В то же время в цитоплазме ряда бактерий, например, Rhodopseudomonas palustris (класс «Alphaproteobacteria»), а также архей, например, Halococcus morrhuae (фила АII Euryarchaeota) в последнее время обнаружены не окруженные мембраной магниточувствительные кристаллы железа. Физиологический смысл образования этих включений, по-видимому, заключается в детоксикации хорошо растворимого катиона Fe2+путем его частичного окисления в малорастворимый катион Fe3+. Образовавшийся кристалл магнетита FеО • Fе2О3 может выполнять первичную функцию запасного включения, а за счет своих магнитных свойств — вторичную функцию органеллы, обеспечивающей магнитную ориентацию.
Параспоральные микрокристаллы. На одной из промежуточных стадий спорообразования у некоторых бацилл, например, Bacillus thuringiensis (фила ВХIII
Firmicutes) в цитоплазме материнской клетки, или экзоспориуме образуется один или несколько кристаллов бипирамидальной, кубоидной или полиэдральной формы размером 0,1-1,5 мкм. Они состоят из протоксина Р1, или δ-эндотоксина — белка молекулярной массой 130-140 кДа, который кодируется плазмидным (реже —хромосомным) геном. После попадания в кишечник личинки насекомого из протоксина Р1 под воздействием щелочных протеаз образуется биологически активный полипептид молекулярной массой 30-80 кДа. Он изменяет ионную проницаемость клеток кишечного эпителия, что быстро вызывает летальный энтомопатогенный эффект.
Благодаря способности к образованию параспоральных микрокристаллов бактерии имеют возможность занять благоприятную нишу. Путем биотехнологического производства 6-эндотоксина человек получил в свои руки сильное средство для борьбы с вредителями сельского хозяйства.
8.3.11.6. Магнитосомы, ацидокальцисомы и керитомические вакуоли
Микрокомпартменты, принадлежащие к данной группе, окружены унитарной мембраной и заполнены твердофазным неорганическим или органическим материалом. По характеру выполняемых функций они относятся к запасающим включениям.
Магнитосомы. Магнитосома — это одетый унитарной мембраной монокристалл химически чистого магнетита или грейгита (Fe3S4), имеющий кубооктаэдрическую, пулевидную или гексагонально-призматическую форму и размер 35-120 нм. Она представляет собой элементарный магнитный домен с постоянным направлением вектора и максимально возможной напряженностью магнитного поля. Как правило, магнитосомы объединяются в продольную цепочку, реже они образуют две или больше параллельных, или пересекающихся цепочек; некоторые виды магнитных бактерий содержат одиночные магнитосомы.
Бактерии, способные накапливать магнетит, выделены из морских, речных и озерных донных илов, а также из песчаных пляжей, рисовых полей и затапливаемых почв. Все они — хемоорганотрофные микроаэрофилы или анаэробы.
Свойства магнитосом и их физиологичесая роль лучше всего изучены на примере Magnetobacterium gryphiswaldense и М. magnetotacticum. В середине 1990-х годов были разработаны методы культивирования этих бактерий, что позволило получить препараты мембраны магнитосомы (англ. magnetosome membrane, ММ), играющей ключевую роль в процессе цитоплазматической биоминерализации магнетита.
В биогенезе магнитосомы участвуют белки MagA и Mps (рис. 28). У магнитобак- терий предполагается процесс, аналогичный эндоцитозу — от СМ отпочковываются везикулы, играющие роль предшественников магнитосом, а затем в них из цитоплазмы закачивается железо. Белок MagA действует как Fе2+/Н+-антипортер, переносящий из цитоплазмы катионы Fe2+. Белок Mps содержит ацил-СоА-связывающий мотив (его гомолог стимулирует отпочкование мембранных везикул от донорной trans-цистерны аппарата Гольджи) и может выполнять функцию ацилазы, с помощью которой запускается процесс инвагинации СМ.
Рис. 28. Биогенез магнитосомы. СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; MagA — Fе2+/Н+-антипортер; Мрs — ацилаза.
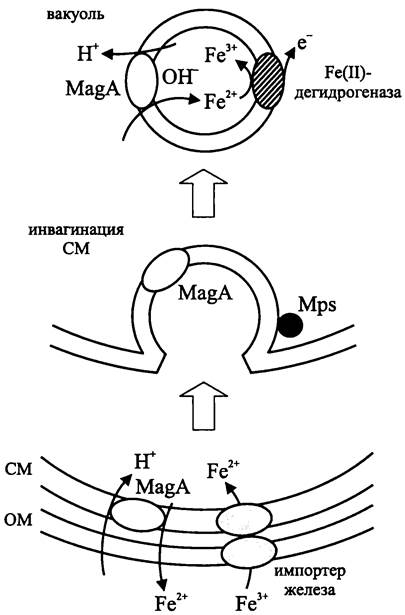
Уникальный механизм образования ММ —это исключение, подтверждающее правило Роджера Стэниера: «Бактерии не имеют цитоза». Это же правило не позволяет одной бактерии находиться в цитоплазматическом компартменте другой бактерии. Известны только два исключения: хищные бактерии рода Daptobacter прони-
кают в цитоплазму пурпурных бактерий рода Сhromatium, а некоторые виды протестов и насекомых имеют интрамитохондриальных симбионтов, принадлежащих к классу «Alphaproteobacteria» (см. раздел 8.5.1.3).
Считается, что магнитосомы в первую очередь обеспечивают магнитную ориентацию. Благодаря тому, что они обладают собственным или наведенным магнитным моментом, клетка приобретает магнитный диполь, расположенный вдоль ее продольной оси. Это позволяет пассивно ориентироваться вдоль силовых линий магнитного поля Земли, которые в обоих полушариях проходят под углом к горизонту. Данное явление, известное еще с конца XVI в., называется наклонением магнитной стрелки.
Внутренняя магнитная стрелка направлена к переднему концу клетки у «северных» магнитных бактерий и к заднему концу у «южных» магнитных бактерий (рис. 29). В обоих случаях активно плавающим магнитным бактериям задается направление движения — ко дну водоема. Там они находят благоприятные микроаэрофильные условия, а также необходимые для роста органические субстраты.
Рис. 29. Магнитная ориентация бактерий. Нh и Нv—горизонтальная и вертикальная составляющие вектора геомагнитного поля.

По-видимому, магнитосомы изначально служили для запасания железа, и на этой основе развилась вторичная функция — обеспечение магнитной ориентации. Неслучайно магнитосомы образуются не только у многих бактерий, живущих в наземных нишах с высокой концентрацией железа, но и у некоторых морских бактерий, обитающих в верхнем водном слое открытой акватории, где ориентация по отношению ко дну смысла не имеет.
Железо требуется и для роста клеток, и для образования магнетита. Импорт Fе3+, сопряженный с его восстановлением, приводит к накоплению катиона Fе2+, который способствует повреждению ключевых биомолекул. Поэтому клетке необходимо иметь специализированную систему, с помощью которой внутриклеточная концентрация закисного железа поддерживалась бы на низком уровне. Наилучший способ, позволяющий защититься от чрезмерного накопления токсичного Fе2+- это
депортировать его в твердофазный магнетит, упакованный в мембрану магнитосо мы. Если это действительно так, то магнитосому можно считать функциональным аналогом бактериоферритина, который позволяет клетке запасать железо без вредных последствий.
Ацидокальцисомы. Как уже отмечалось, типичные полифосфатные гранулы не имеют ограничивающей мембраны (см. раздел 8.3.11.4). Однако получены данные, что в клетках Agrobacterium tumefaciens они окружены унитарной мембраной. В ее состав входит протонная «неорганическая» пирофосфатаза, которая переносит протоны внутрь этой органеллы в результате гидролиза цитоплазматического ФФ (см. II том учебника). Очевидно, что для биосинтеза полифосфатов внутри этой органеллы должна находиться полифосфаткиназа и туда должен транслоцироваться АТФ.
Аналогичные включения, названные «ацидокальцисомами», были описаны только у некоторых паразитических протистов, например, Toxoplasma gondii и Trypanosoma cruzi.Предполагается, что их функция выражается в запасании катионов Са2+, иммобилизованных на полифосфатном носителе.
Керитомические вакуоли. Микрокомпартменты с подобным названием (греч. keratos — рог и tomein — срезать; «похожий на слоистый спил рога») встречаются у многих нитчатых цианобактерий (см. раздел 7.2.2.10).
Эти органеллы варьируют по размеру, имеют сферическую форму, кристаллическое или аморфное содержимое и окружены унитарной мембраной (по-видимому, она представляет собой видоизмененную мембрану тилакоида). Биохимический состав керитомических вакуолей не определялся; возможно, он меняется от случая к случаю. По характеру контрастирования можно заключить, что в керитомической вакуоли накапливается органический материал. Функции данных включений неизвестны; не исключено, что они являются балластом, изолирующим от цитоплазмы дефектные продукты метаболизма, например, прионизированные белки.
8.3.11.7. Карбоксисомы и полигидроксиалканоатные гранулы
В данном разделе речь пойдет о двух типах, комбинированно ограниченных микрокомпартментов, которые окружены неунитарной мембраной и расположены в цитоплазматическом компартменте.
Карбоксисомы (полиэдральные тела). Эти микрокомпартменты имеются у всех цианобактерий. Кроме того, они описаны у многих хемолитоав- тотрофов и, по крайней мере, у одной пурпурной бактерии — Rhodomicrobium vannielii. Анализ препаратов этих включений, полученных из серной бактерии Halothiobacillus neapolitanus, показал, что они содержат иммобилизованный ключевой фермент «цикла Кельвина-Бенсона-Бессема» — рибулозобисфосфат-каброксилазу/оксигеназу RuВisСО-I (см. II том учебника), на основании чего они были названы «карбоксисомами».
Карбоксисомы (1-50 на клетку) расположены в зоне нуклеоида и представляют собой электроноплотные многогранники размером 100-700 нм, состоящие из «протомеров» диаметром 10 нм и окруженные монослойной белковой мембраной толщиной 3-4 нм. Если вначале считали, что карбоксисома имеет форму икосаэдра, или правильного 20-гранника, то сейчас установлено, что это сложный полиэдр, обладающий 6-лучевой симметрией в горизонтальной и двумя осями симметрии в вертикальной проекции.
Карбоксисома содержит до 300 молекул RuBisСО и на 60% состоит из молекул этого фермента, упакованного в виде паракристаллической структуры. Вопрос о том, является ли каброксисома монолитом, или внутри нее есть свободное пространство, остается открытым.
Ограничивающая мембрана, которая составляет до 20% массы карбоксисомы, содержит не менее 5 белков молекулярной массой 5-130 кДа. Гены, кодирующие эти белки (ccmK-ccmO, cso1-cso3), расположены на хромосоме по соседству с генами большой и малой субъединиц RuBisCO (rbcL и rbcS). Остальные 20% массы карбоксисомы приходятся на ~25 неидентифицированных белков, локализация которых неизвестна.
Другие ферменты, кроме RuBisCO, в карбоксисоме достоверно не выявлены, хотя главным претендентом на эту роль является карбоангидраза, катализирующая реакцию: СО2 + Н2О <—> HCO3- + Н+.
Хотя RuBisCO основательно изучена в плане молекулярного состава и ферментативной кинетики, физиологическая роль компартментализации этого фермента в карбоксисоме по-прежнему остается загадкой (он вполне успешно функционирует и в монодисперсном виде в цитозоле). На этот счет существуют две основные гипотезы:
1. Карбоксисома является формой запасания RuBisCO, что позволяет сохранить фермент в интактном состоянии и, таким образом, уменьшить метаболические издержки на его адаптивный биосинтез и обновление.
2. Иммобилизованная RuBisCO обладает модифицированными кинетическими свойствами по сравнению с растворимой формой, что способствует более эффективному участию этого фермента в ассимиляции неорганического углерода.
В пользу первого предположения (о запасающей функции карбоксисомы) говорит тот факт, что доля иммобилизованной RuBisCO изменяется в зависимости от содержания неорганического углерода, а при гетеротрофном росте карбоксисомы вообще не образуются.
В пользу второго предположения (об активной роли карбоксисомы в ассимиляции СО2) свидетельствует то, что RuBisCO имеет очень низкое сродство к этому субстрату (Кm >150 мкМ). Поэтому физиологически целесообразно сконцентрировать СО2 в области активного центра или сделать так, чтобы кинетические свойства последнего существенно изменились. Действительно, концентрация «растворенного неорганического углерода» (англ. dissolved inorganic carbon, DIC) в среде обитания цианобактерий и хемолитоавтотрофов обычно не превышает 15 мкМ, и бактерии необходимо сконцентрировать его до физиологически оптимального уровня ~1 мМ. При невозможности накопить субстрат RuBisCO — газообразный СО2, остается другой путь — повысить его концентрацию в зоне активного центра. Для этого в мембране карбоксисомы должна находиться карбоангидраза (см. выше).
Механизм биогенеза карбоксисомы неизвестен. Есть предположение, что вначале путем самосборки образуется белковая мембрана, а затем оболочка карбоксисомы заполняется молекулами RuBisCO.
Время внесло определенные коррективы в традиционные представления о карбоксисомах. В частности, показано, что генетические детерминанты карбоксисом существуют не только у автотрофов — некоторые энтеробактерии, например, Е. coli, Klebsiella pneumoniae и Salmonella entérica содержат оперон pdu (сокр. англ. propanediol utilization), ответственный за использование пропан- диола и этаноламина. Один из генов в составе этого оперона, pduA, гомологичен генам ссmК и csoS1, кодирующим белки мембраны карбоксисомы. Продукты экспрессии этих генов образуют полиэдральные включения, названные «энтеросомами». Считается, что их роль связана с утилизацией пропандиола и других редких субстратов. Точнее, внутри энтеросомы могут быть изолированы токсичные продукты, образующиеся при расщеплении пропандиола и этаноламина. Кроме того, ферменты, действующие на эти субстраты (диолдегидратаза и этаноламин-аммиаклиаза), могут изолироваться в энтеросоме от ингибирующего воздействия кислорода.
Полигидроксиалканоатные гранулы. Эти запасающие компартменты широко распространены у бактерий, а также встречаются у некоторых архей (примером
служит Halobacterium mediterranei). В зависимости от объекта и окружающей среды полигидроксиалканоатные гранулы являются эндогенным источником углерода или энергии, обеспечивающим конкурентоспособность микроорганизма в субоптимальных и стрессовых условиях.
Полигидроксиалканоаты (англ. polyhydroxyalkanoic acid, PHA) — это сложные полиэфиры 3-, 4- или 5-гидроксикарбоновых кислот (рис. 30). Azospirillum brasilense образует гомополимер 3-гидроксибутирата (англ. polyhydroxybutyrate, РНВ). В отличие от этого Alcaligenes eutrophus образует сополимер гидроксиалканоатов с короткой цепью из 3-5 атомов углерода (3-гидроксипропионата, 3-гидроксибутирата, 4-гидроксибутирата, 3-гидроксивалерата и 5-гидроксивалерата), a Pseudomonas oleovorans — сополимер 3-гидроксиалканоатов с цепью средней длины (С6-С12)- Молекулярная масса полигидроксиалканоатов составляет 1-250 кДа, и они образуют крупные (100-800 нм) округлые кристаллические включения, состоящие из нескольких тысяч молекул. Они окружены неунитарной белковой мембраной (толщиной 2-4 нм), в которой, по-видимому, находится комплекс PHA-синтазы, а также деполимеризующие ферменты.
Рис. 30. Мономеры полигидроксиалканоатов. А — 3-гидроксипропионат; Б — 3-гидроксибутират; В — 4-гидроксибутират; Г — 3-гидроксивалерат; Д — 5-гидроксивалерат; Е — 3-гидроксиалканоат с цепью средней длины.
Полигидроксиалканоаты накапливаются в количестве до 50% биомассы при несбалансированном росте, когда клетке не хватает азота при избытке экзогенного источника органического углерода, в частности ацетата или глюкозы. Реже они образуются у (фото)автотрофных организмов, например, у цианобактерий.
Реконструирован биосинтетический путь, приводящий к накоплению РНВ у А. brasilense. Он начинается с конденсации двух молекул ацетил-СоА при помо-
щи 3-кетотиолазы (ген рhbА). Образовавшийся ацетоацетил-СоА восстанавливается НАДФН-зависимой редуктазой (ген рhbВ) в 3-гидроксибутирил-СоА, который затем полимеризуется РНВ-синтазой (ген рhbС). Все три гена сгруппированы в общий кластер на хромосоме.
Полигидроксиалканоаты рассматриваются как перспективный пластический материал бытового назначения, поскольку им можно придать любую форму, а после использования они легко поддаются биологическому разрушению.
8.4. Периплазматический компартмент
Поскольку периплазматический (лат. peri — вокруг) компартмент лежит вокруг цитоплазматического, может сложиться впечатление, что это лишь преддверье бактериальной клетки. На самом же деле это мультифункциональный компартмент, и его значение очень велико. Грамотрицательные бактерии получили его в наследство от своего прародителя, и он представлен во всех филах древа Bacteria за исключением филы BXIV Actinobacteria.
Происхождение ОМ, или наружной границы периплазматического компартмента неизвестно. Существует версия, что эта специализированная билипидная мембрана сформировалась вторично на основе S-слоя, или архаичной белковой мембраны (см. раздел 8.5.2.3). По-видимому, в период ранней эволюции бактерий в него поэтапно интегрировались липиды и целый ряд специфических белков. В пользу такого сценария свидетельствуют данные о слабой гомологии между субъединицами S-слоя и поринами, входящими в состав ОМ (см. раздел 8.4.1.2).
Как мы уже отмечали, наружную границу периплазматического компартмента образует ОМ (см. раздел 8.1.1).
Помимо периплазматического компартмента, который служит цитологическим «маркером» грамотрицательных бактерий, у разных прокариотов встречается его аналог — квази-периплазматический компартмент. Это понятие появилось сравнительно недавно. Оно имеет отношение к бактериям из группы «mycolata» (см раздел 8.5.2.2), а также к бактериям и археям, содержащим поверхностный S-слой (см. разделы 8.5.2.3 и 8.5.3.1).
В обоих случаях под квази-периплазматическим компартментом имеют в виду зону, которая снаружи ограничена липопротеиновыми или белковыми структурами и содержимым которой является морфоскелет, ферменты, транспортные белки, сигнальные молекулы и т. д.
Относительный размер квази-периплазматического компартмента у бактерий меньше, чем у архей (соответственно, 10 и 20-40% объема цитоплазматического компартмента). Очень редко, например, у экстремального термофила Ignicoccus sp. он достигает 60-75% объема цитоплазматического компартмента.
Размер периплазматического компартмента обычно не превышает 5% объема клетки, хотя бывают исключения. В качестве примера можно привести род Nitrospira (фила BVIII Nitrospirae), где эта величина составляет ~30%, при расстоянии 30-40 нм между СМ и ОМ. «Гигантский» периплазматический компартмент, до 70% объема клетки, имеют ядерные симбионты протистов — бактерии рода Holospora (класс «Alphaproteobacteria»).
В том случае, когда между СМ и ICM поддерживается прямая связь, или мембранный континуум (см. раздел 8.3.5), периплазматический компартмент сообщается с полостями ICM, и за счет этого его объем многократно увеличивается.
На электронных микрофотографиях расстояние между СМ и ОМ близко к толщине унитарной мембраны (7-8 нм). Это мешает воспринимать данную часть клетки кале компартмент. Иногда зону между СМ и ОМ даже обозначают как «слой» L3 клеточной стенки (три других «слоя» это: L1 — между СМ и муреином, L2 — муреин и L4 — наружная мембрана).
Важно понять, что, независимо от объема, зона между СМ и ОМ — это настоящий компартмент, уникальный атрибут основного бактериального морфотипа и полифункциональная органелла грамотрицательных бактерий.
Знакомство с периплазматическим компартментом мы начнем с описания наружной мембраны.
8.4.1. Наружная мембрана
ОМ принадлежит к числу унитарных мебран (см. раздел 8.3.1). Она состоит из билипидного слоя, с которым связаны интегральные, амфипатические и поверхностные белки.
По виду ОМ напоминает СМ или ICM. Это выражается в такой же, как у них, толщине (7-8 нм) и таком же трехслойном контрастировании на ультратонких срезах (темный-светлый-темный). Однако она выполняет уникальные функции, связанные с особенностями ее молекулярного состава.
Общие свойства. Унитарные мембраны обычно характеризуются ярко выраженными диэлектрическими свойствами. Они не пропускают гидрофильные ионы — прежде всего «сопрягающие» катионы Н+ или Na+(см. II том учебника). Это позволяет им запасать энергию в форме разности электрохимических потенциалов этих катионов и в результате служить энергетическими станциями прокариотной клетки.
Хотя ОМ является унитарной мембраной, в ней имеются постоянно раскрытые гидрофильные поры, что не позволяет поддерживать на ней разность электрохимических потенциалов катионов Н+ или Na+. Поэтому ОМ, в отличие от СМ и ICM, не принимает участия в энергетическом метаболизме.
ОМ почти не задействована в конструктивном метаболизме и не имеет отношения к биосинтезу белков, липидов и полисахаридов, хотя участвует в их таргетинге и секреции. Вклад ОМ в ферментативные процессы довольно специфичен — в ней содержатся пермеазы и гидролазы (см. ниже), а в редких случаях с ней связаны окислительные ферменты, например, марганец-оксидаза.
Тем не менее, ОМ является полифункциональной органеллой. У бактерий, обладающих грамотрицательным морфотипом, она отвечает за ряд очень важных клеточных функций. Эта мембранная структура:
— совместно с СМ образует периплазматический компартмент;
— наряду с клеточной стенкой выполняет роль ригидного слоя;
— служит фильтром, который пропускает в периплазматическое пространство мелкие и задерживает крупные гидрофильные молекулы;
— задерживает гидрофобные молекулы;
— иммобилизирует содержимое периплазматического компартмента, в частности «осморегулируемые периплазматические глюканы» (см. раздел 8.4.3);
— участвует в трофическом транспорте, т. е. в поступлении в клетку питательных субстратов (см. II том учебника);
— участвует в экспорте метаболитов и секреции белка, а также в экскреции токсикантов и антибиотиков (см. раздел 8.6.1);
— участвует в неспецифической адгезии, избирательно взаимодействует с антителами, поверхностными рецепторами прокариотных и эукариотных клеток, а также с бактериофагами.
Через узкие поры в ОМ не проходят макромолекулы — прежде всего экзогенные ферменты, которые могли бы лизировать муреиновый саккулус. Поэтому грамотрицательные бактерии «позволяют себе» иметь однослойный муреин стандартного типа, тогда как грамположительные бактерии образуют разнообразные варианты многослойного муреина, более устойчивого против действия гидролитических экзоферментов (см. раздел 8.4.2).
По той же причине гидрофильные антибиотики и амфифильные детергенты, например, желчные кислоты оказывают более слабое воздействие на грамотрицательные бактерии, по сравнению с грамположительными бактериями.
Наконец, в состав ОМ входят липополисахариды — специфические липиды, в молекуле которых имеется экзоплазматический гидрофильный домен. Это ограничивает доступ к поверхности СМ липофильных антибиотиков и других антибактериальных агентов.
8.4.1.1. Липиды наружной мембраны
Рассматривая СМ и ICM, мы отмечали, что они состоят из двух симметричных слоев фосфолипидов — главным образом из цвиттерионного фосфатидилэтаноламина, анионного фосфатидилглицерола и анионного дифосфатидилглицерола (см. раздел 8.3.3.1). Жирнокислотные «хвосты» длиною в 12-18 атомов углерода часто содержат циклопропановые кольца, что стабилизирует жидкостно-кристаллическое состояние мембраны (у эукариотов аналогичную роль выполняют ненасыщенные жирные кислоты).
Переходя к ОМ, мы должны отметить ее первую особенность — липиды распределены между двумя полумембранными листками асимметрично. В то время как цитоплазматический полумембранный листок по составу не отличается от СМ и ICM, и липиды покрывают до 50% его поверхности, экзоплазматический полумембранный листок, как правило, образован липополисахаридами. Исключение составляет род Neisseria, а также цианобактерии, у которых экзоплазматический полумембранный листок содержит одновременно липоолигосахариды и фосфолипиды.
Липополисахариды. В конце XIX в. было показано, что грамотрицательные бактерии образуют термостабильные токсины, которые вызывают лихорадку и сопутствующие болезненные проявления у подопытных животных. В отличие от термолабильных белковых токсинов грамположительных бактерий, например, дифтерийного или столбнячного токсина, они имеют небелковую природу и представляют собой липополисахариды. Поскольку эти токсины не выделяются из клеток, а прочно связаны с ними, их называют «эндотоксинами».
Липополисахариды, сокращенно ЛПС (англ. lipopolysaccharide, LPS), являются главными компонентами ОМ грамотрицательных бактерий.
У кишечных бактерий и псевдомонад ЛПС состоят из четырех структурных блоков (рис. 31):
1. «Липида А»;
2. Олигосахаридного внутреннего кора из двух молекул 8-углеродного сахара (октозы) и трех молекул 7-углеродного сахара (гептозы);
3. Олигосахаридного наружного кора из пяти молекул 6-углеродного сахара (гексозы);
4. Полисахаридной наружной цепи из повторяющихся олигосахаридных субъединиц, каждая из которых образована 2-8 мономерами гексозы.
Рис. 31. Строение липополисахарида.
АглюN — остаток N-ацетилглюкозамина; КДО — остаток кетодезоксиоктоновой кислоты; гал, геп и глю — остатки галактозы, манногептозы и глюкозы; R1 -R3 —остатки жирных кислот; О-цепь — полисахаридная наружная цепь; Р — фосфатная группа. Арабскими цифрами обозначены номера атомов углерода.
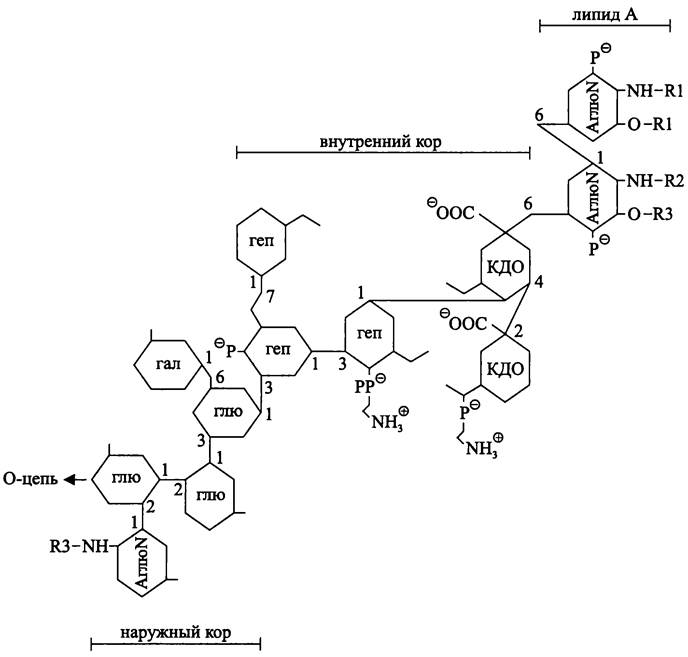
Bordetella pertussis, Haemophilius influenzae, Neisseria meningitidis и ряд других представителей филы ВХII Proteobacteria, а также Bacteroides fragilis (фила ВХХ Bacteroidetes)синтезируют ЛПС без наружной цепи. Представители рода Chlamydia (фила BXVI Chlamydiae) образуют ЛПС не только без наружной цепи, но и без наружного кора. Наконец, в редких случаях (Borrelia burgdorferi, Deinococcus radiodurans, Sphingomonas paucimobilis, Thermotoga maritima и Treponema pallidum) грамотрицательные бактерии вообще не синтезируют ЛПС, и их функции выполняют гликосфинголипиды.
Липид А. В начале 1980-х годов была получена серия мутантов Е. coli и Salmonella typhimurium с различными нарушениями структуры ЛПС. В отличие от образующих гладкие колонии штаммов дикого типа, или «S-вариантов» (от англ. smooth — гладкий), они не содержат наружной полисахаридной цепи и образуют колонии с шероховатой поверхностью, или «R-варианты» (от англ. rough — шершавый).
В зависимости от структуры сохранившейся части ЛПС, R-варианты обозначаются как Ra-Re (рис. 32). На примере мутантов Е. coli, дефектных по синтезу гептоз, показано, что для их роста минимально необходим ЛПС Re-типа, состоящий из липида А с присоединенным к нему остатком «кетодезоксиоктоновой» кислоты (КДО), она же «манноктулозоновая», она же 2-кето-3-дезокси-D-глицеро-D- манно-октоновая (см. рис. 33). В виде исключения, например, у Acinetobacter sp. и Burkholderia sepacia, место КДО занимает «талоктулозоновая» кислота, она же D- глицеро-D-тало-октоновая, производное редкого сахара талозы (см. рис. 55).
Рис. 32. Изменение доменной структуры липополисахарида у мутантов Е. coli (варианты Ra-Re) по сравнению с диким типом (S-вариантом). АглюN — остаток N-ацетилглюкозамина; КДО — остаток кетодезоксиоктоновой кислоты; гал, геп и глю — остатки галактозы, манногептозы и глюкозы.

Липид А служит якорем, который фиксирует ЛПС в ОМ. Он представлен множеством копий (до 106 в клетке Е. coli). Его химическое строение было определено в начале 1980-х годов (рис. 33).
Рис. 33. (2 х КДО)-липид А. КДО — остаток кетодезоксиоктоновой кислоты; Р — фосфатная группа; геn — остаток манногептозы.
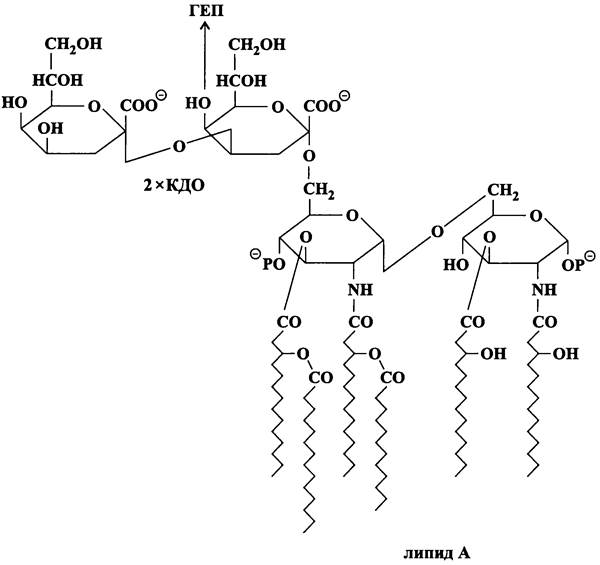
Основу липида А составляют две молекулы глюкозамина, соединенные между собой гликозидной связью β1 —> 6. В положении 1 на редуцирующем конце и положении 4 на нередуцирующем конце имеется по фосфатной группе. К обоим остаткам глюкозамина через амидную и сложноэфирную связь присоединены С14-жирные кислоты, гидроксилированные в положении 3. Благодаря этому нередуцирующий конец дисахарида «вторично» ацилируется, в результате общее число жирнокислотных радикалов достигает шести. В отличие от С16-С20 жирных кислот в глицерофосфолипидах жирные кислоты липида А обычно состоят из 12-14 атомов уг-
лерода и не содержат двойных связей. К нередуцирующему концу липида А через связь β2 —> 6 присоединяется первая молекула КДО. Мутанты по биосинтезу КДО нежизнеспособны, т.е. этот компонент кора имеет решающее значение для функций ЛПС.
Отклонения от консервативной структуры липида А встречаются крайне редко. В качестве примера можно привести липид А клубеньковой бактерии Rhizobium leguminosarum — вместо глюкозамина он содержит другие сахара и не фосфорилирован.
Биосинтез Re-эндотоксина — (2 х КДО)-липида А. Исходным материалом для биосинтеза липида А служит активированная форма глюкозы — уридиндифосфат- глюкоза, или УДФ-глюкоза (рис. 34). Она превращается в УДФ-глюкозамин, а затем в N-ацетилглюкозамин. После этого при помощи ацилтрансферазы Lp x A в положение 3 вводится остаток 3-гидроксимиристиновой кислоты (С14). Он поступает от особого донора — «ацилпереносящего» белка АСР (сокр. англ. acyl carrier protein), с которым он связан тиоэфирной связью. Затем цинкзависимая деацетилаза Lp x C удаляет из положения 2 ацетильную группу. С помощью ацилтрансферазы Lp x D на
освободившееся место вводится второй остаток 3-гидроксимиристиновой кислоты. В результате образуется УДФ-диацилглюкозамин. Наконец, пирофосфатаза LрхН расщепляет пирофосфатную связь, и УДФ-диацилглюкозамин превращается в диацил- 1-Р, или «липид X».
Рис. 34. Биосинтез (2 х КДО)-липида А. УДФ — уридиндифосфат; УМФ — уридинмонофосфат; ЦМФ — цитидинмонофосфат; АглюN — остаток N-ацетилглюкозамина; глюN — остаток глюкозамина; КДО — остаток кетодезоксиоктоновой кислоты; Р — фосфатная группа; АСР — «ацилирующий» белок.

На следующем этапе дисахаридсинтаза LрхВ переносит остаток диацилглюкоза- мина от УДФ-диацилглюкозамина в положение 6 липида X. В результате образуется тетраацил-дисахарид- 1-Р. Последний фосфорилируется в положении 4 с помощью специфической киназы LрхК. Образуется тетраацил-дисахарид-1,4-бисфосфат, или «липид IVА».
Первый остаток КДО вводится в положение 6 на нередуцирующем конце дисахарида с помощью КДО-трансферазы WaaA, а второй остаток КДО — в положение 4
первого остатка. После присоединения двух остатков КДО нередуцирующий конец (2 х КДО)-липида IVA «вторично» ацилируется с помощью ацилтрансфераз LpxL и LpxM. Как и раньше, донорами жирнокислотных остатков служат лаурил-АСР (С12) и миристил-АСР (С14), в результате чего образуется (2 х КДО)-липид А, т.е. Re-эндотоксин.
Все ферменты, участвующие в биосинтезе (2 х КДО)-липида А, находятся в цитоплазме или на внутренней поверхности СМ. Поскольку дальнейшая сборка ЛПС и его интеграция в ОМ происходят в периплазматическом компартменте, необходимо, чтобы (2 х КДО)-липид А транслоцировался через ОМ. Детали этого процесса неизвестны. По-видимому, в нем участвует АВС-пермеаза MsbA (см. II том учебника). Этот транспортер выполняет роль флиппазы, переносящей липофильный субстрат с одной поверхности мембраны на другую; он представляет собой гомодимер из шестидоменных субъединиц, между которыми находится экспортный канал. Для таргетинга (2 х КДО)-липида А используются неидентифицированные периплазматические белковые факторы и система, обеспечивающая его интеграцию в ОМ.
Олигосахаридный внутренний кор. На следующих этапах биосинтеза ЛПС к Re- эндотоксину присоединяются 6-8 моносахаридов кора. Затем к нему добавляются до 50 повторяющихся единиц О-цепи (см. далее). Олигосахаридный кор состоит из внутренней «гептозно-октозной» части и наружной «гексозной» части.
Внутренний кор имеет консервативное строение и содержит редкие сахара — КДО и L-глицеро-D-манногептозу (или D-глицеро-D-манногептозу), остатки которых соединены между собой связями β1 —> 3 и β1 —> 7. К каркасу кора могут присоединяться остатки этаноламина (рис. 33).
У представителей рода Chlamydia внутренний кор которых не содержит гептоз, а образован тремя остатками КДО.
Биосинтез внутреннего кора. Субстратами при биосинтезе КДО служат арабино-5-Р и фосфоэнолпируват, а ее активированной формой служит цитидинмонофосфат-КДО (ЦМФ-КДО). В свою очередь, манногептоза синтезируется из седогепту- лозо-7-P с помощью изомеразы GmhA, а ее активированной формой является АДФ- L-глицеро-D-манногептоза.
Оба мономера внутреннего кора переносятся из цитоплазмы в периплазматиче- ский компартмент. Там с помощью гликозилтрансфераз WaaA и WaaQ они присоединяются к ранее экспортированному (2 х КДО)-липиду А.
Олигосахаридный наружный кор. Это также довольно консервативная часть ЛПС — например, все иммунологически различимые разновидности, или серотипы рода Salmonellaотносятся к Ra-варианту. Сюда входят пиранозные формы гексоз (2- амино-2-дезоксигалактоза, 2-амино-2-дезоксиглюкоза, N-ацетилглюкозамин, галактоза, глюкоза и др.), соединенные между собой связями β1 —> 2, β1 —> 3 и β1 —> 6.
Биосинтез наружного кора. Мономеры наружного кора активируются с помощью УДФ. После экспорта в периплазматическое пространство гликозилтрансфераза WaaG присоединяет их к внутреннему кору.
Полисахаридная наружная цепь («О-специфическая» цепь, О-антиген). Это самый вариабельный домен ЛПС. Буква «О» является сокращением от немецкого предлога ohne —без («без подвижности») и указывает на то, что данный антиген является не жгутиковым, а «соматическим», т. е. он связан с телом клетки.
Наружные цепи состоят из разнообразных сахаров (> 60 структурных типов), которые часто модифицируются путем амидирования, ацетилирования, метилиро-
вания и фосфорилирования, а также могут присоединять к себе аминокислотные остатки (что в целом дает >30 типов заместителей).
Чаще всего наружная цепь содержит до 50 повторяющихся олигосахаридных субъединиц, хотя иногда встречаются гомополимерные варианты. Например, Legionella pneumophilaобразует полимер из 75 остатков «легионаминовой» кислоты (5-ацетамидино-7-ацетамидо-8-O-ацетил- 3,5,7, 9-тетрадезокси-D-глицеро-L-галакто-нононовой).
Характерным примером повторяющейся олигосахаридной субъединицы является разветвленный тетрасахарид галактоза абеквоза-манноза-рамноза, который образуется у возбудителя сальмонеллеза S. typhimurium (рис. 34).
У разных бактерий повторяющиеся олигосахаридные субъединицы варьируются за счет использования разных мономеров (в том числе энантиомеров) в разных количественных сочетаниях и за счет разного взаимного расположения мономеров, а также благодаря разным неуглеводным заместителям. Поэтому у каждого штамма наружная цепь уникальна и высокоиммуногенна — например, в случае Е. coli различают до 170 разных О-серотипов.
Дистальные участки наружной цепи ЛПС иногда выступают за пределы капсулы. Это имеет негативные последствия для бактерии, поскольку через ЛПС они могут связываться с антителами и другими белковыми факторами плазмы крови. Неслучайно некоторые патогенные бактерии проявляют адаптационную «фазовую» изменчивость структуры О-антигена, благодаря чему им удается уклониться от иммунной системы макроорганизма. Например, в дистальном участке ЛПС Neisseria gonorrhoeae повышается содержание сиаловой кислоты (N-ацетилнейра- миновой). Это яркий пример мимикрии у бактерий, поскольку сиаловая кислота служит типичным компонентом поверхностного гликопротеинового антигена животной клетки.
В некоторых случаях бактерии образуют сразу несколько ЛПС, которые различаются по длине наружной цепи. В частности, Р. aeruginosa содержит А-ЛПС (общий антиген) с короткой незаряженной цепью и В-ЛПС, боковая цепь которого длиннее и заряжена отрицательно. Последняя выдается на 40 нм над поверхностью ОМ и постоянно совершает колебательно-вращательные движения.
Биосинтез наружной цепи. В отличие от кора, мономеры которого наращиваются по отдельности, наружная цепь образуется в периплазматическом компартменте из повторяющихся олигосахаридных субъединиц и уже в готовом виде «пришивается» к кору.
Сборка олигосахаридных субъединиц из активированных гликозидных мономеров происходит с помощью периферических гликозилтрансфераз на внутренней поверхности СМ. Готовая субъединица присоединяется к липидному переносчику и экспортируется с помощью флиппазы.
В качестве переносчика олигосахаридных субъединиц используется ундекапре- нилфосфат (англ, undecaprenyl phosphate, und-P). Это липофильное соединение представляет собой фосфорилированную форму С55спирта ундекапренола, или «бактропренола», состоящего из 11 изопреноидных единиц (рис. 35). Ундекапренил- фосфат также используется в качестве переносчика субъединиц муреина и капсульных полисахаридов (см. разделы 8.5.2 и 8.5.4). У эукариотов аналогичную роль выполняет долихол, состоящий из 14-22 изопреновых единиц. Он переносит олигосахариды, которые используются при биосинтезе поверхностных гликопротеинов.
Рис. 35. Ундекапренилфосфат.
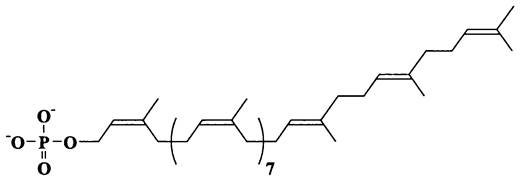
Рис. 36. Сборка О-цепи липополисахарида (Wzy-путь). СМ — цитоплазматическая мембрана; und-Р — ундекапренилфосфат; Р — фосфатная группа; гал, ман, рам и абе — остатки галактозы, маннозы, рамнозы и абеквозы; УДФ, ГДФ, ТДФ и ЦДФ — уридиндифосфат, гуанозиндифосфат, тимидиндифосфат и цитидиндифосфат; УМФ — уридинмонофосфат.
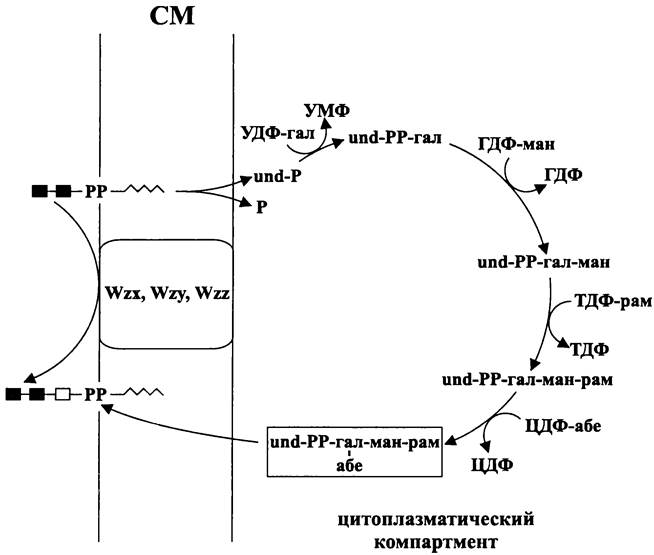
В случае S. typhimurium механизм сборки О-цепи, или «Wzy-путь», выглядит следующим образом (рис. 36). Сначала активированная форма галактозы (УДФ-галактоза) с помощью гликозилтрансферазы WbaP передает остаток галактозо-1-P на und-P, в результате чего образуется und-PP-галактоза, играющая роль затравки. На нее последовательно переносятся остатки маннозы, рамнозы и абеквозы. Все три реакции катализируются соответственно гликозилтрансферазами WbaU, WbaN и WbaW и протекают с участием активированных форм сахаров (ГДФ-маннозы, ТДФ-рамнозы и НДФ-абеквозы). После сборки олигосахаридной субъединицы она переносится с помощью флиппазы Wzx к сайту полимеризации, расположенному на наружной поверхности СМ. Реакция осуществляется с помощью О-полисахарид- полимеразы Wzy, состоящей из 11-13 трансмембранных доменов. Растущая цепь полимера «перебрасывается» от und-PP, который заякоривает ее в наружной поверхности СМ, на нередуцирующий конец только что экспортированной олигосахаридной субъединицы. Освободившийся und-PP после отщепления ортофосфата превращается в свою активную форму — und-P, которая используется при сборке новой олигосахаридной субъединицы и т. д. Таким образом, за каждый цикл к редуцирующему концу полисахаридной цепи присоединяется одна олигосахаридная единица. Конечная длина цепи контролируется белком Wzz. Полученный полимер «пришивается» к кору ЛПС за счет использования энергии сахар-фосфатной связи.
Кроме описанного Wzy-пути существует самостоятельный путь для биосинтеза неразветвленного О-полисахарида. Полимер удлинняется трансацетилазой WecA путем поочередного присоединения гликозильных остатков на внутренней поверхности СМ, а затем экспортируется с помощью АВС-пермеазы. Существует и третий путь, когда О-полисахарид образуется с помощью интегральной синтазы, которая осуществляет векторную полимеризацию и одновременно с этим проталкивает полимер через СМ.
Транслокация липополисахарида на экзоплазматическую поверхность наружной мембраны. Чтобы окончательно занять свое положение, ЛПС должен пересечь периплазматический компартмент и ОМ. Как это происходит, еще не ясно. Есть данные, что новые молекулы ЛПС появляются на клеточной поверхности в локальных участках, которые могут соответствовать «контактам Байера», или зонам адгезии между СМ и ОМ (см. раздел 8.4.1.4). По-видимому, через периплазматический компартмент переброшен транслокационный мост, образованный «адгезионным» белком (англ. membrane fusion protein). ЛПС может выходить на поверхность ОМ через отверстие порина ТоIС, с помощью которого Е. coliэкспортирует гемолизин, импортирует бактериоцины и взаимодействует с бактериофагами.
Функциональное значение липополисахаридов. Липид А и кор несут большое количество отрицательно заряженных фосфорильных остатков и карбоксильных
групп, т. е. являются полианионами. Через катионные мостики они электростатически взаимодействуют и друг с другом, и с интегральными белками. Это стабилизирует структуру самой ОМ и придает ригидность всей клеточной оболочке.
Однако свойство полианиона имеет и негативную сторону — поликатионы и хелатирующие агенты связываются с ЛПС, что повышает проницаемость ОМ. Именно это лежит в основе антибактериального действия ЭДТА и таких антибиотиков, как дефензины и полимиксины.
За счет гидрофильности наружной цепи ЛПС грамотрицательные бактерии защищены от гидрофобных антибактериальных агентов, а также от нейтральных и анионных детергентов. В свою очередь, через белковые поры в ОМ проходят только мелкие гидрофильные молекулы. Большинство антибиотиков гидрофобны или, если гидрофильны, то имеют относительно большую молекулярную массу и поэтому остаются за пределами ОМ.
Взаимодействие эндотоксина с клетками животных. При бактериальных инфекциях ЛПС проявляют высокую биологическую активность, которая полностью зависит от липида А.
В ходе размножения бактерий, а также после их гибели ЛПС высвобождаются в плазму крови. Там они связываются со «связывающим» белком LBP (англ. lipopolysaccharide binding protein), который доставляет их к рецепторам CD14, расположенным на поверхности макрофагов. Через рецепторный мембранный белок TLR4 и цитоплазматический адапторный белок MyD88 активируется транскрипция генов, кодирующих эндогенные медиаторы воспаления (TNF-α, IL1-β, тромбоксан А2, окись азота NO и др.). В малых количествах эндотоксин постоянно поступает в кровь через кишечник и оказывает благотворное действие на макроорганизм в качестве иммуностимулятора, а также активатора защиты от инфекций и злокачественного роста. Однако передозировка эндотоксина при раневых поражениях и обширных ожогах вызывает синдром септического шока, который может даже привести макроорганизм к гибели.
8.4.1.2. Белки наружной мембраны
ОМ богата белками, которые покрывают до 50% ее поверхности и обозначаются как Оmр (сокр. англ. outer membrane protein).
Их разнообразие очень велико — например, у Pseudomonas aeruginosa число идентифицированных Omp-белков достигает 30, а число предполагаемых Оmр-белков — 70.
Оmр-белки подразделяются на главные и минорные. Главные Omp-белки синтезируются постоянно. Они составляют ~80% белков ОМ и представлены большим числом копий (> 105), и поэтому относятся к самым многочисленным клеточным белкам грамотрицательных бактерий. Их характерными примерами служат липопротеин Брауна, липопротеины спирохет, белок ОmрА и порин «общего назначения» OmpF (см. ниже).
В отличие от них минорные белки синтезируются адаптивно и в относительно малом числе копий. Их примерами являются «специализированный» порин PhoE и «транспортный» рецептор FhuA (см. ниже).
Некоторые белки ОМ были вначале описаны как рецепторы для бактериофагов, что и отразилось в их тривиальных названиях. Например, белок LamB (от англ. lambda) используется в качестве рецептра бактериофагом![]()
В структурно-функциональном отношении белки ОМ подразделяются на:
— амфипатические липопротеины, которые содержат липидный домен на N-концевом участке и связывают ОМ с муреиновым саккулусом;
— интегральные белки (например, ОmрА), которые в основном выполняют структурную роль;
— интегральные белки (например, порин «общего назначения» OmpF), которые образуют гидрофильные каналы и участвуют в импорте или экспорте на уровне ОМ.
Препротеины белков Оmр синтезируются с длинной N-концевой последовательностью из ~20 аминокислотных остатков, которая требуется для транслокации через СМ с помощью генеральной секреторной Sec-системы. Эта последовательность удаляется с помощью «сигнальной», или «лидерной» пептидазы, расположенной на периплазматической поверхности СМ (см. раздел 8.6.1).
Липопротеины. В состав ОМ у разных бактерий входят несколько главных и минорных липопротеинов.
Главный липопротеин Брауна (англ. Braun’s lipoprotein). Этот белок назван в честь американского биохимика Брауна (V.Braun), который обнаружил его в конце 1960-х годов у Е. coli.Это был первый Omp-белок, полученный в очищенном виде. Липопротеин Брауна (ген lрр, синоним — mlpА) образуется в числе 7 • 105 копий на
клетку. Он имеет небольшую молекулярную массу (7,2 кДа) и состоит из 58 аминокислотных остатков; полипептидная цепь представляет собой α-спираль.
Липопротеин Брауна уникален тем, что к N-концевому остатку (Суs-1) через тиоэфирную связь присоединяется глицерол, этерифицированный двумя жирными кислотами. Третья жирная кислота связана амидной связью с тем же N-концевым остатком (рис. 37). Примерно одна треть молекул липопротеина Брауна ковалентно присоединяется к муреиновому саккулусу (см. раздел 8.4.3.1). Связь осуществляется между ε-аминогруппой остатка Lys-58 и свободной карбоксильной группой каждого 10-12-го остатка мезодиаминопимелиновой кислоты в составе муреина. Отсюда происходит второе название этого белка — «муреиновый липопротеин».
Рис. 37. Липопротеин Брауна. ДАП — остаток диаминопимелиновой кислоты; R1-R3 — остатки жирных кислот.

На примере Е. coli показано, что препротеин белка Брауна секретируется через СМ при помощи генеральной Sec-системы. Затем он «липидируется» тремя ферментами. Вначале диацилглицерол-трансфераза Lgt (от англ. glycerohprolipoprotein diacylglycérol transferase) переносит диацилглицерол от мембранного фосфатидилг- лицерола на остаток Cys. Затем чувствительная к антибиотику глобомицину пептидаза (англ. lipoprotein signal peptidase, Lsp) отрезает от препротеина сигнальный пептид. Теперь новым N-концевым остатком становится Cys. Наконец, трансацилаза (англ. phospholipidrapolipoprotein transacylase, Lnt) переносит третий остаток жирной кислоты от мембранного фосфолипида на свободную NH2-rpynny N-концевого остатка Lys. Таргетинг липопротеина обеспечивает молекулярный шаперон LolA, а его правильное встраивание в ОМ — молекулярный шаперон LolB.
Предполагается, что липопротеин Брауна поддерживает структуру ОМ, благодаря чему клетки сохраняют форму и целостность. Точечные мутанты по гену lрр и lрр- -мутанты обладают измененной морфологией и образуют дефектную оболочку. Однако у некоторых бактерий (например, Neisseria gonorrhoeae) данный белок отсутствует даже у штамма дикого типа. Липопротеин Брауна не способен быть рецептором для бактериофагов или колицинов.
Помимо липопротеина Брауна в клетках Е. coli содержится минорный липопротеин ЕхсС, также ковалентно связанный с пептидогликаном.
Главные липопротеины спирохет. ОМ зоопатогенных бактерий из филы BXVII Spirochaetes имеет особый белковый состав. Это обеспечивает высокую эластичность клеточной оболочки и работу уникального локомоторного аппарата — периплазматических жгутиков (см. раздел 8.6.2).
Спирохеты образуют от 20 (возбудитель сифилиса Treponema pallidum) до 100 (агент воспалительной болезни Лайма Borrelia burgdorferi) различных липопротеинов, которые играют структурную роль и являются ключевыми факторами вирулентности. Главные липопротеины спирохет (LipL32, OspA, SmpA, OspA, Vmp и др.) отличаются от липопротеина Брауна по строению, механизму экспорта и выполняемым функциям. Они расположены не в цитоплазматическом, а в экзоплазматическом полумембранном листке, поэтому их полипептидный домен ориентирован не в сторону муреинового слоя, а во внешнюю среду. Как и в случае липопротеина Брауна, препротеин секретируется через СМ с помощью генеральной Sec-системы. Затем он липидируется тремя ферментами. Однако за его таргетинг и интеграцию в ОМ отвечают не молекулярные шапероны LolAB, а уникальная система, узнающая кластер из полярных аминокислот в области N-концевого остатка Lys. Паттерн экспрессии липопротеинов изменяется в зависимости от условий окружающей среды (прежде всего, температуры), что позволяет спирохетам адаптироваться к клещам- переносчикам, диким животным и человеку.
Липопротеины являются факторами адгезии спирохет к фибронектину и клеткам различных тканей человеческого организма. Из-за высокой концентрации липопротеинов другие белки ОМ экранируются от хозяйских антител, а благодаря высокой антигенной вариабельности они сами уходят от иммунного ответа. Подобно ЛПС, липопротеины спирохет являются стимуляторами воспаления. Они тоже связываются с рецепторами CD14 на поверхности макрофагов, но это происходит без участия доставляющего белка LBP, и мембранным рецептором служит не белок TLR4, а белок TLR2. На основе липопротеинов разрабатываются вакцины для борьбы с боррелиозами, лептоспирозами и другими заболеваниями, возбудителями которых являются патогенные спирохеты.
Минорные липопротеины. В настоящее время описан ряд минорных липопротеинов. Они не вступают в ковалентную связь с пептидогликаном, и выполняемые ими функции неизвестны. Примерами служат липопротеины LppL, OmlA и OprI, имеющиеся у Р. aeruginosa.
OmpA (от англ. outer membrane protein А). Это главный белок ОМ. Е. coli (~105 копий на клетку). Он представлен in situ в мономерной форме.
OmpA состоит из 325 аминокислотных остатков. В него входят три функциональных домена:
— N-концевой гидрофильный экзоплазматический домен, построенный из четырех длинных гидрофильных петель;
— N-концевой трансмембранный бочковидный домен, образованный восемью антипараллельными β-складчатыми цепями, которые соединены тремя короткими периплазматическими шпильками;
— С-концевой периплазматический домен, непосредственно взаимодействующий с пептидогликаном.
ОmрА также называют «термолабильным» белком, поскольку его электрофоретическая подвижность изменяется после кипячения в растворе додецилсульфата натрия.
Белок OmpA выполняет структурную роль, нековалентно связывая ОМ с муреи- новым саккулусом, что стабилизирует клеточную оболочку. В то же время через его бочковидный домен отфильтровываются мелкие гидрофобные молекулы, т. е. одновременно он является порином «общего назначения» (см. ниже). Помимо структурной и транспортной функции белок OmpA служит рецептором для бактериофагов M1, К3, Ox2 и Tull, а также для антимикробных белков бактериального происхождения — колицинов L и К. Благодаря взаимодействию OmpA с гликозилированными
эпитопами рецепторных белков эндотелия облегчается проникновение патогенных бактерий сквозь гематоэнцефалический барьер.
Таким образом, белок ОmрА полифункционален. Его гомологи (например, белок OprF Р. aeruginosa) имеются у всех бактерий, обладающих грамотрицательным морфотипом.
Белок ОтрА и другие бочковидные трансмембранные белки, содержащие β-цепи, находятся только в ОМ. В то же время в ней отсутствуют α-спиральные белки, входящие в состав СМ или ICM.
Порины. Как уже было сказано, ОМ является полупроницаемым барьером для гидрофильных молекул. Большинство содержащихся в ней белков участвует в образовании гидрофильных пор. Такие белки известны под общим названием «порины», и к ним относятся:
— неспециализированные порины, или порины «общего назначения», через которые диффундируют небольшие гидрофильные молекулы;
— специализированные порины, которые имеют сайты для связывания лигандов, что облегчает их диффузию;
— высокоаффинные порины, которые участвуют в энергозависимой транслокации крупных молекул (например, витамина В12 или хелатов Fe3+), присутствующих в окружающей среде в низкой концентрации.
Порины представляют собой интегральные белки с молекулярной массой 30-50 кДа. Они образуют в СМ крупные гидрофильные поры, через которые путем диффузии в периплазматический компартмент поступают питательные субстраты и из него выделяются продукты метаболизма. В настоящее время установлено, что порины встречаются и у бактерий, обладающих грамположительным морфотипом (см. раздел 8.5.1).
В отличие от интегральных белков СМ, порины не содержат протяженных гидрофобных доменов. В то же время, в отличие от цитозольных белков, гидрофобные остатки у них сосредоточены преимущественно на внешней, а на внутренней поверхности. Канал порина выстлан полярными остатками, образующими систему водородных связей. Поэтому порины образно называют белками, «вывернутыми наизнанку». Ближе к поверхности билипидного слоя сосредоточены анионные группы, которые через кальциевые и магниевые мостики взаимодействуют с анионными группами кора ЛПС.
Порины in situ образуют тримеры (исключение составляет мономерный порин OprF Р. aeruginosa).
Согласно рентген-структурным данным (впервые полученным для Rhodobacter capsulatus), основу поринов составляет бочковидная структура из 16 антипараллельных β-цепей, расположенных под углом 30-60° к плоскости ОМ. С экзоплазматической стороны β-цепи соединены длинными гидрофильными α-спиральными петлями, а с периплазматической — короткими шпильками из 1- 4 аминокислотных остатков. Третья наружная петля загибается внутрь гидрофильного канала, сужая его просвет до глазка (англ. eyelet) диаметром около 10 А, что и определяет «эксклюзивный диаметр» порина, т. е. максимальную массу проходящей через него молекулы. Таким образом, канал порина имеет широкий вход, широкий выход и центральное сужение, размер которого может изменяться в зависимости от липидного окружения, концентрации осмолитиков в периплазматическом компартменте (см. раздел 8.4.3), ионной силы и pH окружающей среды, а также от воздействия приложенного электрического поля.
Неспециализированые порины, или «порины общего назначения». Эти порины образуют гидрофильные каналы, пронизывающие ОМ. Возможность прохождения
через них и его скорость зависят только от физико-химических свойств субстрата. Эксклюзивный диаметр поринов общего назначения составляет ~600 Да.
Хотя неспециализированные порины не содержат сайтов для связывания субстрата, все же они проявляют определенную пропускную селективность. Например, порин OmpF оказывает предпочтение нейтральным молекулам и катионам. Его гомолог ОmрС обладает аналогичными свойствами. В обоих случаях они создают барьер на пути проникновения неполярных молекул.
Несмотря на то, что порины «общего назначения» не обладают строгой пропускной селективностью, их биосинтез находится под регуляторным контролем. Например, когда Е. coliпопадает в пищеварительный тракт, т. е. в среду с относительно высоким осмотическим давлением, это регистрируется осмосенсорным белком EnvZ, который расположен в СМ и выполняет роль проводника сигнала в двухкомпонентной сигнальной системе. Белок EnvZ фосфорилирует регулятор ответа — цитозольный белок OmpR. В свою очередь, фосфорилированная форма белка OmpR подавляет экспрессию порина OmpF и стимулирует экспрессию порина ОmрС. Поскольку эксклюзивный диаметр ОmрС меньше, чем у OmpF, это повышает устойчивость микробной клетки к желчным кислотам. В связи с вышеописанной избирательностью порины OmpF и ОmрС (а также их гомолог ОmрКЗб, имеющийся у Klebsiella pneumoniae) называют «осмопоринами».
Специализированные порины. В отличие от поринов «общего назначения», специализированные порины имеют стереоспецифические сайты связывания субстрата, которые подчиняются закономерностям ферментативной кинетики, в частности способны к насыщению. Диаметр гидрофильной поры у специализированных поринов обычно меньше, чем у поринов «общего назначения», и составляет ~6 Â.
К числу специализированных поринов относятся прежде всего «фосфопорины», хотя реально они оказывают предпочтение анионам вообще, а не только фосфату и фосфорилированным соединениям. Они обычно образуются при фосфорном голодании. Примерами служат транспортер ортофосфата PhoE, одновременно являющийся рецептором для бактериофага ТС45 (Е. coli), транспортер ортофосфата ОргР (Р. aeruginosa) и транспортер пирофосфата OprO (Р. aeruginosa).
Другие специализированные порины используются для транспорта моно-, ди- и олигосахаридов. Например, порин ОргВ (Р. aeruginosa) предназначен для транспорта глюкозы. В свою очередь, «мальтопорин» LamB (Е. coli) индуцируется мальтозой и облегчает ее диффузию, а также диффузию мальтодекстринов. Он имеет маленькую пору, адаптированную к линейным олигсахаридам, и пропускает только глюко- зильные остатки. Его гомолог ScrY (5. typhimurium) используется для транспорта сахарозы.
Для транспорта дипептидов существует специализированный порин OprD (Р. aeruginosa), а для транспорта нуклеозидов — порин Tsx (Е. coli).
«Выделительные» порины (англ. efflux porin). С помощью этих поринов бактерии приобретают резистентность, в частности, к антибактериальным перпаратам. Выделительные порины функционируют не самостоятельно, а в составе трехсубъединичной экспортной системы. Она состоит из двух α-спиральных компонентов — АТФ-зависимой помпы и «адгезионного» белка, а также из ß-спирального домена — порина ТolС. У Е. coli такая система (АсгАВ-ТolC) обеспечивает множественную лекарственную устойчивость и выброс из клетки токсичных соединений. В свою очередь, в клетках Р. aeruginosa имеются 18 выделительных поринов, причем некоторые из них гомологичны порину ТolС (например, ОmрН).
Порин TolC входит также в состав секреторной системы I типа (см. раздел 8.6.1).
Высокоаффинные порины («транспортные» рецепторы). В отличие от поринов «общего назначения» и специализированных поринов, эти порины не образуют постоянно открытых каналов, и поэтому их называют «раскрывающимися» поринами (англ. gated porins). Другая особенность этих поринов состоит в том, что они не только обладают высокой специфичностью по отношению к лигандам, но и участвуют в энергозависимом транспорте, т. е. обладают свойствами пермеаз (см. раздел 9.1.1). Это единственный случай, когда энергетически «инертная» ОМ может преобразовывать энергию.
В качестве примеров высокоаффинных поринов можно назвать белки FecА, FepA, FhuA (TonA) и Cir, участвующие в транспорте хелатов железа, а также белок BtuB, который транспортирует витамин В12. Для детального знакомства с ними мы отсылаем читателя ко II тому учебника.
Завершая тему, коснемся ее эволюционного аспекта. Сохранились ли порины в наружной мембране митохондрий и пластид, произошедших от бактерий? В настоящий момент известен только один «органельный» гомолог порина — анионный канал VDAC (сокр. англ. voltage-dependent anione channel), имеющийся у митохондрий.
Прочие белки ОМ. Помимо рассмотренных белков в состав ОМ входят:
— ферменты (фосфолипаза OmpLA и протеаза ОmрТ);
— белок ОmрХ;
— вспомогательные белки (англ. outer membrane auxilary protein).
Фосфолипаза OmpLA используется при секреции белка колициногенными штаммами Е. coli и этиологическим агентом язвы желудка — Helicobacter pylori. Она проделывает отверстия в ОМ, через которые наружу выходят антибактериальные полипептиды — колицины, а также белковые факторы вирулентности. Белок OmpLA содержится в мембране в мономерной или димерной форме. По строению он сходен с белком ОmрА.
Белок ОmрТ представляет собой специализированную протеазу, которая является одним из факторов патогенности Е. coli.
Белок ОтрХ играет важную роль в жизни вирулентных штаммов Е. coli, поскольку он участвует в нейтрализации иммунных систем макроорганизма. Когда бактерии испытывают стресс, этот белок синтезируется в большом количестве. Он входит в общую группу с такими белками, как Ail-белок Yersinia enterocolitica, способствующий адгезии этой бактерии и ее проникновению внутрь хозяйской клетки, а также с белками PagC и Rck, обеспечивающими персистенцию S. typhimurium в макрофагах. Все эти белки существуют in situ как мономеры и по строению сходны с белком ОmрА. В отличие от последнего, у них половина ß-спиральной бочковидной структуры выступает наружу и связывается с чужеродными белками, которые, в свою очередь, содержат поверхностные ß-спиральные участки.
К вспомогательным белкам относятся представители группы «секретинов». Они образуют дистальный участок систем II и III секреции белка (см. раздел 8.6.1), а также дистальный участок систем секреции полисахаридов (см. раздел 8.5.4.1).
8.4.1.3. Везикулы наружной мембраны
Процесс отпочкования от ОМ мембранных везикул диаметром 20-500 нм называется «блебингом» (англ. bleb — пузырек). Он выполняет важную приспособительную роль в жизни многих грамотрицательных бактерий. Сходство между блебингом и
апокриновой секрецией у эукариотов чисто внешнее. Во-первых, она происходит на уровне СМ. Во-вторых, в ней принимает участие цитоскелет.
Образование блебинг-везикул может быть неспецифическим явлением и вызываться тем, что ОМ растет быстрее муреинового слоя (см. раздел 8.4.3.1). С другой стороны, блебинг зависит от специфических условий питания. Например, у Bacteroides gingivalis его предпосылкой служит дефицит геминовых соединений.
В состав блебинг-везикул входят типичные компоненты ОМ — ЛПС, фосфолипиды и Оmр. Все они имеют такую же ориентацию, как в бактериальной клетке. При образовании блебинг-везикула захватывает часть содержимого периплазматического компартмента, за исключением муреина. На примере Р. aeruginosa показано, что блебинг-везикулы содержат периплазматические ферменты, в том числе гемолизин, пептидогликан-гидролазы (автолизины), протеазу, проэластазу, щелочную фосфатазу и фосфолипазу С (см. раздел 8.4.2).
Блебинг-везикулы служат надежным средством дистанционной доставки всех этих ферментов вместе с упаковочным мембранным материалом к клеткам-мишеням или в тот участок внешней среды, где проявится их биологическое действие.
Если блебинг-везикулы соприкасаются с клеткой грамположительной бактерии, они вскрываются и освобождают автолизины, что вызывает локальное разрушение стенки (чему не препятствует пористый S-слой; см. раздел 8.5.2.3). В отличие от этого, при контакте с грамотрицательной бактерией мембрана блебинг-везикулы сливается с ОМ, и внутреннее содержимое поступает в периплазматический компартмент. Автолизины диффундируют по всему объему периплазматического компартмента, и клеточная стенка разрушается сразу во многих местах. Блебинг-везикулы малоэффективны в отношении активно растущих и делящихся клеток, где нанесенные повреждения легко репарируются. Благодаря действию на гетерологичные бактерии продуцент автолизина добывает себе субстраты для роста, в то время как его сородичи устойчивы к «знакомым» автолизинам.
Для патогенной бактерии доставка факторов вирулентности в мембранной упаковке служит ценной альтернативой свободной диффузии, поскольку защищает их от инактивации сывороточными агентами.
Благодаря тому, что блебинг-везикулы инкапсулируют протеазы и фосфолипазы, оба фермента выделяются в инфекционный очаг в нативном состоянии и в высокой концентрации. В частности, этим объясняется сильная коллагеназная активность возбудителей парадонтоза В. gingivalis и Capnocytophaga gingivalis, а также их способность разрушать мембраны эпителия и клеток соединительной ткани.
Когда эндотоксин и мурамилпептиды (см. раздел 8.4.3.1), содержащиеся в блебинг-везикулах, доставляются в очаг воспаления, они вызывают синергический эффект, т. е. более сильную ответную реакцию по сравнению с действием по отдельности.
Показано, что при блебинге Р. aeruginosa в ОМ-везикулы, в числе прочего, попадает β-лактамаза. Этот фермент позволяет бактериям не только разрушать антибиотики во внеклеточной среде, но и гидролизовать пенициллины и другие β- лактамные антибиотики, проникшие в периплазматический компартмент. Блебинг служит средством кооперации патогенов — продуценты β-лактамазы снабжают этим ферментом тех бактерий, которые сами его не синтезируют.
8.4.1.4. Контакты Байера
Во второй половине 1960-х годов на ультратонких срезах плазмолизированных клеток грамотрицательных бактерий были обнаружены зоны адгезии между ОМ и СМ. Число таких зон составляет 200-400 на клетку, они покрывают до 5% поверхности СМ и имеют толщину ~18 нм при ширине 20-30 нм. В честь открывшего эти структуры американского цитолога Байера (М. Bayer) их называют «контактами Байера» (англ. Bayer junction).
В области контактов Байера в муреиновом слое образуется крупное отверстие, что позволяет СМ и ОМ сблизиться вплотную. Однако окончательного слияния с образованием гибридной мембраны, по-видимому, не происходит.
Физической основой контакта Байера могут быть компоненты экспортных комплексов, выделяющих из клетки акридиновые красители (комплекс АсгАВ), гемолизин (комплекс HlyBD) и гидрофобные антибактериальные соединения (комплекс EmrAB). Со стороны ОМ в состав этих комплексов входит выделительный по- рин ТоlС, а со стороны СМ — выделительная помпа (соответственно, АсгВ, НlуВ и EmrB). Контакт между порином и помпой обеспечивает «адгезионный» белок (соответственно, AcrA, HlyD и EmrA).
Функциональная роль контактов Байера проблематична. Есть основания считать, что через них выделяются вышеперечисленные вещества, транслоцируются ЛПС и капсульные полисахариды, а также экспортируются субъединицы фимбрий. Установлено, что в контактах Байера адсорбируются некоторые бактериофаги. По- видимому, через них они инъецируют свою нуклеиновую кислоту.
Считается, что контакты Байера — это «долговечные» структуры, хотя высказана и оппортунистическая точка зрения, согласно которой они образуются только в момент секреции и исчезают, как только она завершилась.
8.4.2. Периплазма
Термин «периплазма» (англ. periplasm), обозначающий растворимое содержимое периплазматического компартмента, предложил Питер Митчел в начале 1960-х годов. Задолго до того, как на электронно-микроскопических снимках обнаружили свободную зону между СМ и ОМ, уже было известно, что при осмотическом шоке в окружающую среду «вытекает» ряд ферментов (англ. menstruum). На основании этого был сделан упреждающий вывод, что за пределами СМ у грамотрицательных бактерий существует особый компартмент, в котором содержатся ферменты и неферментные белки.
Периплазма представляет собой гелеобразный матрикс, состоящий из белков и уникальных олигосахаридов — «осморегулируемых периплазматических глюканов». Наряду с этим в ней присутствует транзитный материал, необходимый для обновления клеточной стенки, ОМ, экзоплазматического ригидного слоя и покровов (капсулы и чехла); кроме того, в нее секретируются субъединицы придаточных структур (жгутиков, фимбрий и шипов). Таким образом, через периплазму проходят разнообразные макромолекулы, качественный состав и соотношение которых определяются индивидуальным типом метаболизма, эндогенной регуляцией и условиями внешней среды.
Периплазматические белки. К белковой фракции периплазмы принадежат: ферменты, связывающие белки, транспортные белки и молекулярные шапероны.
Ферменты. Важнейшими периплазматическими ферментами являются тиолгди- сульфид-оксидоредуктазы, участвующие в окислительном образовании или восстановительном разрыве дисульфидной связи между двумя остатками Cys:
R1-SH + HS-R2<-> R1-S-S-R2+ 2e- + 2Н+,
где R1и R2— участки полипептидной цепи, фланкирующие аминокислотный мотив СС (Cys-Cys).
Тиол:дисульфид-оксидоредуктазы и тиол:дисульфид-изомеразы исправляют положение дисульфидной связи, что позволяет белкам принять правильную стабильную конформацию. Соответствующие ферменты входят в суперсемейство тиоредоксинов, активный центр которых содержит мотив СХХС (два остатка Cys, разделенные двумя аминокислотами).
Цитоплазма представляет собой среду с сильнее выраженными восстановительными свойствами, чем периплазма. Дисульфидные связи в цитоплазматических белках восстанавливаются при помощи тиоредоксинов (исключение составляют некоторые археи, в цитоплазме которых содержатся белки с дисульфидными связями). В периплазме, напротив, создаются окислительные условия, и там сосредоточены белки с дисульфидными связями. Таким образом, если цитоплазма проявляет только дисульфидредуктазную активность, то в периплазме помимо нее локализованы дисульфидоксидазная и дисульфидизомеразная активности.
Дисульфидные связи в белках, которые Е. coli секретирует в периплазму, например в щелочной фосфатазе, белке ОmрА, холерном и коклюшном токсинах, а также в субъединицах адгезионных фимбрий образуются при помощи тиол:дисульфид- оксидоредуктазы DsbA — растворимого мономерного белка с молекулярной массой 21 кДа (рис. 38). Этот фермент имеет редокс-потенциал -120 мВ. С помощью гидрофобных взаимодействий он связывается с развернутыми или частично свернутыми полипептидами. Для регенерации окисленной формы DsbA используется тиол:дисульфид-оксидоредуктаза DsbB (20 кДа), состоящая из четырех интегральных доменов. В свою очередь, DsbB «сбрасывает» электроны на хиноны (ан)аэробной дыхательной цепи (см. II том учебника). Гомологи белков DsbA и DsbB существуют у многих грамотрицательных бактерий.
Рис. 38. Коррекция дисульфидных связей в периплазматических белках. СМ — цитоплазматическая мембрана; DsbА и DsbВ — тиол:дисульфид-оксидоредуктаза; DsbС — тиол:дисульфид-изомераза; Dsb — дисульфидредуктаза; Тгх — тиоредоксин; DsbЕ — тиоредоксиноподобный белок.

Для коррекции ненативного положения дисульфидной связи в периплазматиче- ских белках, содержащих больше двух остатков Cys, некоторые представители классов «Betaproteobacteria» и «Gammaproteobacteria» используют растворимую гомо- димерную тиол:дисульфид-изомеразу DsbC (23 кДа). Она восстанавливает «неправильную» дисульфидную связь электронами, полученными от дисульфидредуктазы DsbD, в которой имеются 8 трансмембранных доменов. В свою очередь, DsbD получает восстановительные эквиваленты от НАДФН. Промежуточным переносчиком является представитель семейства тиоредоксинов (белков, участвующих в окислительно-восстановительных реакциях путем обратимого окисления своих тиоловых групп), редокс-потенциал которого равняется -270 мВ. «Правильная» дисульфидная связь, как и обычно, образуется с помощью DsbA.
Кроме того, DsbD может передавать электроны альтернативному акцептору — амфипатической субъединице DsbE, или CcmG (20 кДа). Этот тиоредоксиноподоб- ный белок восстанавливает отстатки Cys в апопротеине цитохрома с, что на завершающей стадии созревания позволяет ковалентно присоединить к нему геминовую группу.
Помимо тиол:дисульфид-оксидоредуктаз и тиол:дисульфид-изомераз в периплазме содержатся разнообразные гидролазы. К их числу относятся гидролазы олиго-
нуклеотидов (фосфогидролазы), гидролазы олигопептидов и гидролазы олигосахаридов. Обязательным компонентом периплазмы является щелочная протеаза. Все эти ферменты выполняют преимущественно катаболические функции и расщепляют на простые молекулы те питательные субстраты, для которых в СМ нет собственных транспортных систем.
Помимо перечисленных ферментов в периплазме присутствуют муреин-гидролазы (автолизины), участвующие в клеточном делении и обновлении муреина.
Связывающие белки. В периплазме находятся белки, которых иногда называют «хеморецепторными», хотя нужно говорить—«периплазматические связывающие белки» (англ. periplasmic binding protein, ВР). Лигандами для них служат органические субстраты, которые диффундируют через каналы поринов в периплазматическое пространство.
Комплекс связывающий белок-субстрат взаимодействует с периплазматическим доменом одного из трех хеморецепторов в СМ:
1. Тар (от англ. transductor for dipeptides);
2. Таг (от англ. transductor for aspartate);
3. Trg (от англ. trans ductor for ribose and galactose).
Четвертый хеморецептор, Tsr (от англ. transductor for serine) распознает серин непосредственно, без участия, связывающего ВР-белка. Через трансферный и цито-
плазматический домены этих рецепторов, которые сообща играют роль проводника сигнала в двухкомпонентной регуляторной системе, информация поступает на регулятор клеточного ответа.
ВР являются индуцибельными белками и в мономерной форме скапливаются на полюсах клетки, достигая концентрации >1 мМ. Три основных класса ВР, несмотря на слабо выраженную взаимную гомологию, имеют сходное строение — два глобулярных домена соединяются при помощи гибкой петли, конформационное изменение которой обеспечивает связывание лиганда.
Конкретными примерами ВР служат:
— Dpp (связывает дипептиды; распознается хеморецептором Тар);
— МаIЕ (связывает мальтозу и мальтодекстрин; распознается хеморецептором Таг);
— MlgB (связывает галактозу и глюкозу; распознается хеморецептором Trg);
— RbsB (связывает рибозу; распознается хеморецептором Trg).
Помимо этого, существует специальный ВР для катиона Ni2+, выступающего в качестве репеллента. Его комплекс с лигандом распознается хеморецептором Таг.
Транспортные белки и молекулярные шапероны. В периплазме находятся специализированные переносчики, осуществляющие таргетинг интегральных белков ОМ. На примере Е. coliпоказано, что как только липопротеин Брауна при помощи генеральной Sec-системы транслоцируется через СМ, он образует комплекс с растворимым белком р20 (20 кДа), который присутствует в числе 200-400 копий на клетку. Только в такой связанной форме он доставляется в ОМ.
Наряду с транспортными белками в периплазме содержатся белки, выполняющие роль молекулярных шаперонов. Они обеспечивают правильный фолдинг секретированных белков, а также корректируют структуру неправильно свернутых белков (см. раздел 8.3.10.2). В частности, роль такого шаперона (помимо основной тиол: дисульфид-изомеразной функции) играет белок DsbC.
Примерами белков, которые служат субстратами для периплазматических шаперонов, является экзофермент пуллуланаза PulD (атакующий связи α1 —> 6 в амилопектине, разветвленной фракции крахмала), а также субъединицы фимбрий и ряд других белков.
Потребность в молекулярных шаперонах для периплазмы стоит особенно остро, поскольку в условиях высокой гидролитической активности там сохраняются белки только в устойчивой к протеолизу конформации, тогда как незрелые белки нуждаются в защите.
Периплазматические олигосахариды. Представители этого класса олигосахаридов, получившие название «мембрано генные» олигосахариды (англ. membrane-derivedoligosaccharide, MDO), или «осморегулируемые периплазматические глюканы» (англ. osmoregulated periplasmic glucan, OPG) пока обнаружены только у ряда представителей класса «Alphaproteobacteria» (Agrobacterium tumefaciens, Azospirillum braziliense, Brucella abortus, Rhodobacter sphaeroides, Sinorhizobium meliloti), класса «Betaproteobacteria» (Ralstoniasolanaceae) и класса «Gammaproteobacteria» (Erwinia chrysanthemi, E. coli, Pseudomonas syringae, Xanthomonas campestris). Однако не исключено, что они шире распространены среди грамотрицательных бактерий. Несмотря на видовые различия в деталях строения, OPG обладают двумя универсальными свойствами:
— они состоят из 5-40 мономеров D-глюкозы, которые соединены преимущественно β-гликозидными связями;
— их концентрация в периплазме возрастает при уменьшении осмомолярности окружающей среды.
Молекулярная масса ОРG (>900 кДа) больше «эксклюзивного диаметра» пори- нов (600 кДа), и поэтому они застревают в периплазматическом компартменте.
ОРG имеют разветвленную линейную или (не)разветвленную циклическую структуру. Остатки глюкозы соединены связями β1 —> 2, β1 —> 3, β1 —> 4 и β1 —> 6 (реже связями α1 — >3 и α1 —> 6). Синтезируются ОРС в цитоплазме из активированного предшественника — УДФ-глюкозы с помощью глюкозилтрансферазы МdоН (97 кДа). По каналу, образованному восемью трансмембранными доменами этого белка, они выводятся на экзоплазматическую поверхность СМ. Там периферическая глюкозилтрансфераза МdоG (56 кДа) присоединяет к ним боковые мономеры.
Рис. 39. Биосинтез осморегулируемых периплазматических глюканов. СМ — цитоплазматическая мембрана; глю — остаток глюкозы; УДФ — уридиндифосфат;
Ф — фосфатная группа.
Во многих случаях углеводный скелет Орв модифицируется путем введения заместителей. Первый из них — это сукцинил. Он передается от сукцинил-СоА с помощью трансферазы МdоС, имеющей 10 трансмембранных доменов. Второй заместитель — это фосфоглицерол. В ходе обновления фосфоглицеролипидов, входящих в СМ, он передается на ОРG с помощью амфипатической трансферазы МdоВ (отсюда и возникло название «мембраногенные» олигосахариды).
За счет большого числа заместителей ОРG приобретают свойство полианиона. Противоионы (К+, Nа+ и др.), находящиеся в донановом равновесии с экзогенными электролитами, служат источником осмотического давления в периплазме. Таким образом, ОРG играют роль осмотического «буфера», который выравнивает тургор-
ное давление по обеим сторонам СМ. При попадании бактерий в гипертоническую среду удельное содержание OPG уменьшается на порядок (с 5% до 0,5%), что связано с ингибированием их биосинтеза. В гипотонической среде биосинтез OPG, наоборот, индуцируется.
Если OPG не имеют анионных заместителей, они выступают в ином качестве — как своеобразные «информационные» молекулы. Дело в том, что при размножении в гипертонической среде в дочерних клетках содержание OPG уменьшается в геометрической прогрессии. Это отслеживается с помощью мембранных сенсоров (EnvZ/ResC у Е. coli) и служит косвенным показателем, что популяция увеличилась в размере. Иными словами, бактерии косвенным путем узнают, что достигнут «кворум», а это важно для кооперативно действующих патогенов. Таким образом, при инфекциях, когда бактерии попадают в среду с более высоким осмотическим давлением, OPG выполняют роль, аналогичную роли феромонов (хотя эффект достигается разными способами), которые секретируются во внешнюю среду и с которыми у бактерий связано «чувство кворума» (см. III том учебника).
Наконец, Agrobacterium tumefaciens и Rhizobium sp. образуют внеклеточные OPG, которые могут играть определенную роль в становлении симбиоза между этими бактериями и растениями. По аналогии с липидом А они оказывают на макроорганизм регуляторное воздействие.
8.4.3. Периплазматические ригидные слои
Известно, что за форму эукариотной клетки отвечает цитоскелет, расположенный в цитоплазматическом компартменте. По последним данным, прокариоты также имеют цитоскелет (см. раздел 8.3.1). Однако форма прокариотов и их способность не разрушаться в гипотонической среде зависят не от него, а от экзоскелета — ригидного слоя клеточной стенки.
Общие сведения о ригидных слоях у прокариотов. Ригидный слой грибов, растений и некоторых из протистов имеет целлюлозную или хитиновую основу. Линейные молекулы этих биополимеров образуют мицеллы, взаимодействующие за счет водородных связей; у прокаритов такой ригидный слой есть только у некоторых архей (см. раздел 8.5.1). Обычно же ригидный слой у прокариотов состоит из сетчатого полимера — пептидогликана, в котором гликановые цепи связаны друг с другом с помощью боковых пептидных цепей.
Пептидогликан уникален тем, что это мешковидная молекула, или саккулус, форма и размер которого соответствуют форме и размеру клетки. Реже встречается саккулус, состоящий из белковых или гликопротеиновых субъединиц; он существует сам по себе или в сочетании с пептидогликаном.
Особый пример связан с «промежуточным» периплазматическим слоем у Deinococcus radiodurans (фила BIV «Deinococcus-Thermus»). По электронно-микроскопическим данным, у этой бактерии между толстым муреиновым слоем и ОМ расположен тонкоструктурированный, или «компартментализованный» слой (англ. compartmentalized layer), который, возможно, обладает ригидностью. Его состав неизвестен.
Ригидные слои грамположительных бактерий и архей находятся в экзоплазматическом компартменте, а ригидные слои грамотрицательных бактерий могут быть в двух компартментах — периплазматическом и экзоплазматическом (см. рис. 4).
Прокариоты без ригидного слоя, или микоплазмы. Микоплазмы встречаются среди представителей обоих прокариотных доменов. Они входят в состав фи-
лы ВХIII Firmicutes (класс «Mollicutes»), а также в состав филы АII Еuryarchaeota (Thermoplasma acidophilum и Т. volcanium).
Физическая целостность микоплазм-бактерий сохраняется за счет того, что они поддерживают низкое осмотическое давление, экскретируя катионы Na+, а также благодаря тому, что они являются паразитами животных и растений, т. е. живут в среде с относительно высоким содержанием осмолитиков. В отличие от микоплазм- бактерий, микоплазмы-археи имеют ригидную СМ. Ее основу составляет монослой кальдархеолов (см. раздел 8.3.3.2). Дополнительную прочность ей придают особые ЛПС (см. раздел 8.5.1.2).
Бактерии без периплазматического ригидного слоя. Примеры бактерий, не имеющих периплазматического ригидного слоя, довольно редки. Можно считать, что это вторично приобретенный «негативный» признак, поскольку у представителей наиболее архаичной филы бактерий (ВI Aquifiqae) уже есть пептидогликан, а у представителей «молодой» филы ВХУ Planctomycetes он отсутствует.
8.4.3.1. Муреиновый саккулус
У большинства бактерий, обладающих грамотрицательным морфотипом, имеется пептидогликановый периплазматический ригидный слой — муреин (лат. тише — стенка). Бактерии, обладающие грамположительным морфотипом, также в большинстве случаев содержат муреин, однако он у них находится в экзоплазматическом компартменте (см. раздел 8.5.2.1).
Поскольку муреин образуется у представителей самых архаичных фил бактерий — ВI Aquificae и ВII Thermotogae, способность к синтезу этого биополимера можно рассматривать как одно из характерных свойств, которое бактерии приобрели сразу же после разветвления ствола глобального древа на три домена.
По химической структуре муреин представляет собой пептидогликан, т. е. гибридный полимер, в котором параллельно ориентированные неразветвленные полигликозильные цепи соединены поперечными олигопептидными цепями.
В результате этого муреин образует гигантскую сетчатую молекулу массой ~5 • 106 Да. Она обволакивает клетку, чем и объясняется название «саккулус». Муреин может состоять из одного или нескольких слоев, связанных олигопептидными цепями. У бактерий, обладающих грамотрицательным морфотипом, муреин обычно однослойный. У бактерий, обладающих грамположительным морфотипом, муреин чаще всего состоит из 20-50 слоев и, кроме того, ковалентно связан с другими гликановыми полимерами (см. ниже).
Под толщиной муреина нужно понимать число молекулярных слоев, а не ширину темного контура, окаймляющего СМ на электронных микрофотографиях ультратонких срезов. У грамотрицательных или грамположительных бактерий она составляет 10-50 нм (у некоторых штаммов цианобактерий из порядка «Oscillatoriales» рекордная толщина — 0,5 мкм). Однако на самом деле муреин не компактен. Судя по результатам, полученным с помощью метода «сверхбыстрой» фиксации (см. раздел 8.3.5), он сильно гидратирован и равномерно диспергирован по всему периплазматическому компартменту, образуя «периплазматический гель».
В то же время муреиновый саккулус — это жесткая структура, прочная на разрыв. Он предохраняет СМ от механического разрушения под действием цитоплазматического тургора; кроме того, форма муреинового саккулуса определяет форму клетки.
Строение муреина. Лучше всего оно изучено на примере грамположительных бактерий Micrococcus lysodeikticus и Staphylococcus aureus.
Гликановая часть муреина консервативна и состоит из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных связями β1 —> 4. Гликановые цепи полидисперсны (10-100 дисахаридов).
Гликозильные мономеры иногда содержат в положении 6 минорные заместители, в частности ацетильную группу. Обычно же через фосфатную группу в положении 6 муреин взаимодействует с другими полимерами клеточной стенки — тейхоевыми и тейхуроновыми кислотами (см. раздел 8.5.2.1). Редко встречаются минорные модификации гликозильного скелета муреина за счет введения манномурамовой кислоты вместо производного глюкозы, а также путем использования вместо N-ацетилглю- козамина деацетилированного или гликозилированного глюкозамина.
Олигопептидная часть более разнообразна (> 100 хемотипов) и, скорее всего, является полидисперсной системой. В вегетативных клетках каждая карбоксильная группа мурамовой кислоты образует амидную связь с аминогруппой концевой аминокислоты боковой пептидной цепи.
Боковые олигопептидные цепи представляют собой три- или тетрапептиды. Они имеют следующие особенности (рис. 40):
— в них регулярно чередуются L-аминокислоты с аминокислотами в «необычной» D-конфигурации;
— проксимальное положение занимает L-аминокислота (L-аланин, глицин или L- серин), а дистальное положение D-аланин;
— в их состав входят «редкие» аминокислоты L, D-диаминопимелиновая (5- карбоксилизин), L-орнитин или D,L-лантионин ((2-амино-2-карбоксиэтил)-цистеин);
— в них присутствует «необычная» пептидная связь, которую образует![]() карбоксильная группа глутаминовой кислоты (α-карбоксильная группа свободна, амиди- рована или замещена на аминокислоту) с α-аминогруппой диаминокислоты: L,D- диаминопимелиновой или L-лизина (α-карбоксильная группа которой свободна или амидирована).
карбоксильная группа глутаминовой кислоты (α-карбоксильная группа свободна, амиди- рована или замещена на аминокислоту) с α-аминогруппой диаминокислоты: L,D- диаминопимелиновой или L-лизина (α-карбоксильная группа которой свободна или амидирована).
Рис. 40. Строение муреина. АглюN — остаток N-ацетилглюкозамина; МК — остаток N-ацетилмурамовой кислоты; МКанг-— остаток ангидрида М-ацетилмурамовой кислоты; ала, гли, глу, ДАП и лиз — остатки аланина, глицина, глутаминовой кислоты, диаминопимелиновой кислоты и лизина; 1 и 1' — N-ацетилмурамидаза; 2 и 2' — N-ацетилглюкозаминидаза; 3 — N-ацетилмурамил-L-аланин-амидаза; 4 — эндопептидаза; 5 — L,D-карбоксипептидаза. Крестом обозначены пептидные мостики в муреине А-типа или В-типа.

Боковые олигопептидные цепи, отходящие от соседних гликановых цепей, соединены друг с другом напрямую или через вставочные пептидные мостики, благодаря чему и образуется саккулус.
Монослой муреина — это упорядоченная гексагональная сеть с порами диаметром ~2 нм, через которые может пройти белок молекулярной массой до 55 кДа. Из- за спиральной закрученности гликановой цепи боковые олигопептидные цепи ориентированы «вверх» или «вниз», что позволяет ковалентно связать друг с другом параллельно лежащие монослои.
От частоты расположения боковых олигопептидных цепей и количества связей между ними зависит жесткость клеточной стенки, а также ее устойчивость к химическим и ферментативным воздействиям.
В соответствии со строением боковых олигопептидных цепей и особенностями связи между ними различают два типа муреина, каждый с несколькими подтипами.
Муреин A-типа (рис. 40) характеризуется тем, что ε-аминогруппа диаминокислоты, находящейся в положении 3 одной олигопептидной цепи, взаимодействует с карбоксильной группой D-аланина, находящегося в положении 4 другой олигопептидной цепи. Связь между ними образуется напрямую или через вставочные пептидные мостики. Последние содержат 1-5 аминокислотных остатков (Lacn, Dacп-Lлиз,
гли5 и др.). В некоторых случаях аминокислотная последовательность пептидных мостиков такая же, как в боковой олигопептидной цепи.
Для большинства грамотрицательных бактерий характерен муреин А1-типа, в котором L, D-диапимелиновая кислота одной боковой пептидной цепи образует амидную связь с D-аланином другой боковой пептидной цепи. Реже вместо L, D-диапимелиновой кислоты используются L-лизин, L-орнитин и L, D-лантионин. Количество
пептидных мостиков в таком муреине обычно невелико. Муреин грамположительных бактерий образует более частую сеть, поскольку содержит многочисленные пептидные мостики.
Муреин В-типа характеризуется тем, что α-карбоксильная группа D-глутами- новой кислоты в положении 2 одной о лигопептидной цепи (положение 1 занимают глицин или серин) взаимодействует с карбоксильной группой D-аланина, находящегося в положении 4 другой о лигопептидной цепи. Поскольку две карбоксильные группы не могут связаться «встык», между ними нужен вставочный мостик, содержащий какую-нибудь диаминокислоту (L-лизин, L-орнитин или L-α, у-диаминомасляную кислоту). Он состоит из 1-2 аминокислотных остатков (Dорн, гли-Lлиз и др.).
Муреин В-типа более редок и встречается только у грамположительных бактерий из фил ВХIII Firmicutes (Acetobacterium woodii, Clostridium barkeri, Eubacterium limosum) и BXIVActinobacteria (Brevibacterium sp., Corynebacterium sp., Microbacterium lacticum).
Особенности строения муреина в эндоспорах. Дифференциация эндоспор — покоящихся резистентных клеток бактерий — происходит в ряд этапов и завершается полным созреванием. Между СМ зрелой эндоспоры и СМ спорангия (материнской клетки) лежит слой муреина, в свою очередь, состоящий из двух субслоев. Тонкий внутренний субслой называется «примордиальным» муреином, а толстый наружный субслой — муреиновым «кортексом» (англ. cortex).
Примордиальный муреин сохраняется при прорастании эндоспоры и по строению идентичен муреину вегетативной клетки. Напротив, муреиновый кортекс обладает уникальными свойствами и быстро разрушается при прорастании эндоспоры. В частности, 25% остатков мурамовой кислоты присоединяют тетрапептидную боковую цепь, а еще 25% присоединяют только L-аланин. Карбоксильная группа остальных 50% остатков вместо того, чтобы присоединять боковую пептидную цепь, ковалентно связывается с собственной деацетилированной аминогруппой, в результате чего образуется 6-лактам мурамовой кислоты. Кроме того, только 3% боковых пептидных цепей сшиты друг с другом (по сравнению с 30% в примордиальном муреине).
Видоизменение муреина при дифференциации эндоспоры преследует, по меньшей мере, две цели. Во-первых, из-за слабой сетчатости кортекс приобретает повышенную гибкость, что способствует его сжатию с целью максимального удаления воды из протопласта эндоспоры. Во-вторых, 6-лактам мурамовой кислоты служит мишенью для действия автолитических ферментов — мурамидазы и глюкозаминидазы. Они начинают разрушать муреиновый кортекс при прорастании эндоспоры, оставляя примордиальный муреин интактным. Функциональная роль монопептид- ной L-аланиновой боковой цепи не выяснена.
Биосинтез блока-мономера муреина. Строительный блок муреина представляет собой дисахарид с боковой пептидной цепью: N-ацетилглюкозамин-N-ацетил- мурамил-пентапептид. Он синтезируется в цитоплазме и экспортируется на наружную поверхность СМ, где включается в состав уже существующего саккулуса.
Сначала активированная форма глюкозы — УДФ-глюкоза превращается в УДФ- N-ацетилглюкозамин (см. рис. 34).
Затем с помощью энолтрансферазы MurA из УДФ-N-ацетилглкжозамина и фосфоэнолпирувата образуется УДФ-N-ацетил-энолпируват (рис. 41). После этого НАДФН-зависимая редуктаза MurB восстанавливает энолпируватную часть молекулы в УДФ-N-ацетилмурамовую кислоту. Далее АТФ-зависимые лигазы МurС, МurD и МurЕ последовательно присоединяют к карбоксильной группе мурамовой кислоты аминокислотные остатки. Наконец, лигаза МurF добавляет к трипептидной боковой цепи дипептид Dала-Dала, в результате чего образуется УДФ-N-мурамил- пентапептид.
Рис. 41- Биосинтез мурамил-пентапептида. УДФ — уридиндифосфат; ФЭП — фосфоэнолпируват; Ф — фосфатная группа; АглюN — остаток N-ацетилглюкозамина; МК — остаток N-ацетилмурамовой кислоты; ала, глу и ДАП — остатки аланина, глутаминовой кислоты и диаминопимелиновой кислоты; МuгС-F — лигазы.

На следующем этапе N-мурамил-пентапептид перебрасывается от гидрофильного переносчика (УДФ) на липофильный переносчик — ундекапренилфосфат, в результате чего образуется «липид I» (см. рис. 35). Потом в результате переноса от УДФ-N-ацетилглюкозамина второго гликозильного остатка ундекапренил-моносахарид-пентапептид превращается в ундекапренил-дисахарид-пентапетид, или «липид II» (рис. 42).
Рис. 42. Биосинтез блока-мономера и его встраивание в муреиновый саккулус. СМ — цитоплазматическая мембрана; АглюN—остаток N-ацетилглюкозамина; МК — остаток N-ацетилмурамовой кислоты; УДФ — уридиндифосфат; УМФ — уридинмонофосфат; Р — фосфатная группа; 1—трансгликозилазная реакция; 2 — транспептидазная реакция. Гармошкой обозначен ундекапренил, черными кружками — аминокислотные остатки в боковой пептидной цепи.
Интересно, что некоторые мелкие бактериофаги вызывают лизис хозяйской клетки, образуя особый Е-белок, который блокирует трансферазу, ответственную за образование липида I. В данном случае проблема выхода из хозяйской клетки решается через воздействие на биосинтез муреина.
Сборка муреина. Ундекапренил-дисахарид-пентапептид спонтанно или при участии флиппазы транслоцируется на наружную поверхность СМ. Там он встраивается в уже существующую сеть муреина, которая через пирофосфатную связь связана с ундекапренилом (который, в свою очередь, заякорен в наружном полу- мембранном листке).
Для встраивания мономерного блока используются мембраносвязанные ферменты, которые ковалентно встраивают его в муреиновый саккулус (рис. 42). Этот процесс состоит из трансгликозилазной реакции (удлинение линейной полигликозидной цепи) и транспептидазной реакции (сшивание боковых пептидных цепей). Они катализируются семейством бифункциональных ферментов, которые расположены в СМ, имеют молекулярную массу 30-90 кДа и называются «пенициллинсвязывающими» белками (англ. penicillin-bindingprotein, РВР).
Антибиотик пенициллин содержит β-лактамное кольцо (рис. 43), сходное по структуре с последовательностью Dала-Dала в о лигопептидной цепи муреина. Поэтому он вступает в ковалентную связь с транспептидирующими активными центрами пенициллинсвязывающих белков PBP1a, PBP1b и РВР2 и необратимо инактивирует их. Таким образом, механизм антибактериального действия пенициллина и других β-лактамных антибиотиков заключается в нарушении сборки муреина.
Рис. 43. Общая формула пенициллинов. R — органической радикал; 1— β-лактамаза (пенициллиназа); 2 — пенициллин-амидаза. Темным фоном выделено 3-лактамное кольцо.

У пенициллина нет мишеней в эукариотной клетке, и поэтому его терапевтическое применение почти не вызывает побочных эффектов. Поскольку бактерии выработали защиту от этого антибиотика, основанную на действии пенициллиназ и пенициллинамидаз, разработано много полусинтетических производных природного бензилпенициллина (пенициллина G). Они устойчивы к этим ферментам, лучше всасываются при пероральном введении и дольше удерживаются в циркуляции. Кроме того, используются химические соединения, блокирующие активный центр пенициллиназы (например, клавулановая кислота).
Аналогичный эффект в отношении сборки муреина оказывает гликопептидный антибиотик ванкомицин, нековалентно блокирующий последовательность Dала-Dала в олигопептидной цепи. Кроме того, биогенез муреина ингибируется D-циклосерином, который из-за стереохимического сходства с D-аланинрм препятствует рацемизации L-аланина в D-аланин и подавляет биосинтез дипептида Dала-Dала.
Сборка муреина требует энергии. Для трансгликозилазной реакции источником энергии служит расщепление пирофосфатной связи между старым муреином и его ундекапренильным «якорем». Освобожденный ундекапренил-пирофосфат «кувырком» перемещается на внутреннюю поверхность СМ, гидролизуется до ундекапре- нилфосфата и повторно используется для биосинтеза и транслокации мономерного строительного блока. В свою очередь, старый муреин переносится на новый мономерный блок, заякоренный в СМ, с образованием гликозидной связи β1 —> 4.
Одновременно с этим образуется прямая (или опосредованная пептидным мостиком) связь между боковой пептидной цепью старого муреина и боковой пептидной цепью мономерного строительного блока. Для транспептидазной реакции источником энергии служит гидролиз связи в концевой последовательности Dала-Dала мономерного строительного блока. Его осуществляют чувствительные к пенициллину мембранные DD-карбоксипептидазы РВР4-6.
Существуют РВР-белки широкого и узкого спектра действия. В частности, у Е. coli бифункциональные PBP1a и PBP1b одновременно осуществляют и трансгли- козилазную, и транспептидазную реакцию, а монофункциональные белки РВР2 и РВРЗ катализируют только транспептидазную реакцию. Первый из них используется для роста клетки в длину, а второй — для образования перегородки при делении.
Лизис муреина. Ферментативное расщепление муреина происходит при помощи муреингидролаз. Эндогенные муреингидролазы бактерий называются авто- лизинами. Они участвуют в обновлении муреина, «переваривают» его при гибели клетки, а также могут секретироваться и в качестве экзогенных литических агентов воздействовать на другие бактерии.
Существуют четыре основных типа муреингидролаз. Одни из них выступают в качестве пептидаз и разрывают амидные связи в боковых пептидных цепях или между ними. Другие являются гликозидазами и разрывают связь β1—>4 в полигли- козидной цепи (рис. 40).
N-ацетил-мурамидаза. Этот фермент расщепляет гликозидную связь между остатком мурамовой кислоты и остатком N-ацетилглюкозамина (рис. 40; 1 и 11). Мурамидазы, относящиеся к «литическим» трансгликозилазам, переносят гликозидную связь в положение 6 того же самого остатка мурамовой кислоты, в результате чего образуется ангидромурамовая кислота. Чем меньше боковых пептидных цепей соединяются пептидными связями, тем больше мономерных блоков муреина оказывается среди продуктов реакции. Мурамидазы, не обладающие трансгликозилазной активностью и поэтому не образующие ангидромурамовую кислоту, называются лизоцимами (англ. lysozyme). Об их широком распространении свидетельствует то, что они имеются у бактерий, грибов, растений и животных. Литические вирусы также кодируют муреингидролазы, что способствует выходу из клетки новых вирионов. У млекопитающих лизоцим содержится в сыворотке крови, слезной жидкости и слюне; больше всего его в лизосомах макрофагов. В случае Е. coliлитическая транс- гликозилаза Slt70 (70 кДа) локализуется в периплазме, а литические трансгликозилазы MltA (38 кДа) и MltB (36 кДа) имеют липопротеиновую природу и связаны с ОМ. Все три фермента действуют выступают в качестве экзомурамидаз и последовательно отщепляют по одному дисахарид-пентапептиду от нередуцирующего конца гликановой цепи.
У грамположительных бактерий (например, Bacillus subtilis) помимо экзомурамидаз существуют эндо мурамидазы, которые проделывают внутренние разрывы в гликановой цепи.
N-ацетил-глюкозаминидаза. Этот фермент расщепляет гликозидную связь между концевым, в случае экзоглюкозаминидазы, или внутренним, в случае эндо- глюкозаминидазы, остатком N-ацетилглюкозамина и остатком мурамовой кислоты (рис. 40,2 и 21).
N-ацетилмурамил-b-аланин-амидаза. Этот фермент расщепляет амидную связь между мурамовой кислотой и остатком L-аланина в боковой пептидной цепи (рис. 40,3). У Е. coliамидаза AmiA (39 кДа) локализуется в периплазме и действует не на полимерный муреин, а на продукты его расщепления — блоки-мономеры.
Эндопептидаза. Этот фермент атакует связи внутри боковой полипептидной цепи или между боковыми пептидными цепями, разрушая сеть муреина (рис. 40,4). У Е. coliэндопептидазы РВР4 (50 кДа) и РВР7 (32 кДа) входят в состав СМ и ингибируются пенициллином.
LD-карбоксипептидаза. Этот фермент расщепляет связь между диаминокислотой и остатком D-аланина в боковой пептидной цепи (рис. 40,5). В отличие от «поставляющей энергию» DD-карбоксипептидазы (см. выше) он мало чувствителен к β-лактамным антибиотикам.
Структурные перестройки муреина. Уже говорилось о том, что муреин обладает высокой ригидностью, что позволяет ему задавать клетке определенную форму и предотвращать осмотический лизис в гипотонической среде. В то же время он подвержен структурным перестройкам, что позволяет бактерии увеличиваться в размере, делиться, а также изменять форму в ходе дифференциации.
При перестройках муреина в нем с помощью автолизинов производятся локальные разрывы. В частности, для расслоения перегородки между дочерними клетками В. subtilisиспользует автолизины LytC (амидаза), LytD (глюкозаминидаза) и LytE/F (эндопептидазы). Бреши в старых акцепторных цепях заполняются новыми донорными цепями, и пептидные связи возобновляются с помощью «интеркалирующих» транспептидаз.
Перестройка структур, находящихся под внутренним напряжением — это сложная задача, поскольку нужно локально лизировать муреин при сохранении его ригидности. Она по-разному решается грамотрицательными и грамоположительными бактериями.
Грамотрицательные бактерии образуют однослойный муреин, что не позволяет без потери ригидности проделать брешь в старом муреине, чтобы заполнить ее новым фрагментом. Выход был найден — согласно модели, получившей название «три в обмен на один», под акцепторный триплет пептидогликановых цепей подслаивается донорный триплет (рис. 44, А). Затем они соединяются пептидными связями (рис. 44, Б). Только после этого удаляется центральная цепь в акцепторном триплете (рис. 44, В), и донорный триплет вдвигается в монослой муреина (рис. 44, Г), увеличивая площадь его поверхности. Естественно, такая модель требует строго согласования при функционировании отдельных компонентов внутри мультиэнзимного комплекса.
Рис. 44. Схема перестройки муреина у грамотрицательных бактерий. А, Б, В, Г — последовательные этапы. Белыми кружками обозначены поперечные срезы акцепторных, черными кружками—донорных пептидогликановых цепей.
Грамположительные бактерии образуют муреин, состоящий из многих слоев, поэтому в этих слоях можно поочередно проделывать бреши, заполняя их свежим материалом. Согласно модели, получившей название «рост снизувверх», при помощи эндопептидаз во внутреннем слое удаляется фрагмент старого муреина (рис. 45, А).
Интактный наружный слой принимает тургорное давление на себя (рис. 45, Б). Брешь заполняется новым фрагментом муреина, который не испытывает механического стресса (рис. 45, В). Затем брешь проделывается в наружном слое, и тургорное давление передается внутреннему слою вместе с расправленным новым фрагментом (рис. 45, Г). В заключение замуровывается брешь в наружном слое (рис. 45, Д).
Рис. 4,5. Схема перестройки муреина у грамположительных бактерий. А, Б, В, Г, Д — последовательные этапы; S — пептидогликановая цепь под механическим стрессом; R — пептидогликановая цепь, не испытывающая стресса.
Сквозные отверстия в муреиновом слое. То, что от непрерывности муреинового слоя зависит физическая целостность клетки, казалось бы, исключает возможность образования в нем сквозных отверстий. Однако это вполне реально, если канал не открыт, а в него вставлена белковая «пробка», и вместе они выполняют роль транспортной органеллы.
Первый пример такой органеллы — это межмембранный контакт Байера (см. раздел 8.4.1.4). Хотя по определению для его образования необходимо пробуравить отверстие в клеточной стенке (у трихомных бактерий это продольная стенка), существование канала не подтверждено прямыми морфологическими наблюдениями.
Второй пример — комплекс микроплазмодесмы, находящийся в поперечной стенке трихомных бактерий. Существование микроплазмодесм подтверждается морфологическими наблюдениями. Однако неизвестно, каково внутреннее строение этой транспортной органеллы, состоящей из канала в клеточной стенке, который изнутри выстлан мембраной десмотубулы.
Третий пример — это «комплекс контактной поры» (англ. junctional pore complex, JPC). У трихомных цианобактерий сквозные отверстия в муреине расположены в продольной стенке наклонно и образуют одинарные или двойные кольца, которые с обеих сторон фланкируют поперечную стенку. Отдельный комплекс контактной поры состоит из трубчатого канала диаметром 14-16 нм, проходящего сквозь СМ и периплазматический компартмент с муреиновым слоем, и из терминальной части размером 20 x 30 нм. Последняя связана с ОМ и образует в ней отверстие диаметром 8 нм. Через канал со скоростью 3 мкм • с-1 выдавливается слизь, что обеспечивает реактивное движение трихома (см. раздел 8.6.2.1). Контактные поры также могут играть роль «перфорационных» отверстий, облегчающих спонтанную или принудительную фрагментацию трихома. В результате этого трихом размножается путем образования многоклеточных дочерних особей.
Любопытно, что цепочка аналогичных органелл (до 50) расположена на «брюшной» стороне одноклеточной бактерии Acetobacter xylinum, синтезирующей внеклеточную целлюлозу.
Обновление и процессинг муреина. Рост и деление клеток сопровождаются активным обновлением муреина. Кроме того, процессинг муреина происходит при его созревании, а также при биогенезе локомоторных органелл, при специфической секреции и приобретении компетентности для генетической трансформации.
Ди-, три- и тетрапептиды, образующиеся в ходе обновления и процессинга муреина, реутилизируются, а часть выделяется в окружающую среду (до 50% в каждом поколении бактерий).
Рудименты муреина. Варианты бактерий, которые не имеют муреинового или иного саккулуса, но тем не менее размножаются на агаризованных средах, называются «L-формами» (от англ. Lister Institute). Они могут появляться спонтанно или под влиянием разных внешних воздействий, в частности при продолжительном культивировании в присутствии пенициллина. При удалении пенициллина они ревертируют в формы, имеющие клеточную стенку. Стабильным L-формам это не свойственно. Они не содержат затравочных фрагментов муреинового саккулуса, хотя сохраняют рудименты муреина. Это не только простые нуклеотидные предшественники моносахаров, но и блок-мономер муреина — ундекапренил-дисахарид- пентапетид. Скорее всего, сборке полимерного муреина мешает отсутствие пенициллинсвязывающих белков, хотя некоторые L-формы синтезируют эти ферменты.
В качестве компенсации стабильные L-формы содержат более жесткую СМ, чем у протопласта, что связано с появлением в ней особых ЛПС, а также с увеличением содержания жирных кислот с короткой углеводородной цепью.
Отдельно упомянем представителей филы ВII Thermotogae. Возможно, что эти архаичные бактерии содержат «лоскуты» муреина или муреиновый саккулус с очень большими сквозными отверстиями.
Наконец, у хламидий (см. ниже) с помощью всех имеющихся методов не удается выявить муреин. Тем не менее, они образуют РВР и проявляют чувствительность к пенициллину, которая пока не находит объяснения.
8.4.3.2. Белковый саккулус
Альтернативой муреиновому саккулусу служит очень редко встречающийся у бактерий белковый саккулус.
Белковый периплазматический ригидный слой у хламидий. Хламидии (фила BXVI Chlamydiae), которые являются облигатными внутриклеточными паразитами и размножаются в вакуолях человека и животных, проявляют ряд парадоксальных свойств. В частности, они не содержат маркера муреина — мурамовой кислоты, хотя у них есть РВР и они чувствительны к ингибиторам биосинтеза пептидогликана, пенициллину и D-циклосерину. На электронных микрофотографиях хламидий не видно муреинового слоя, что подтверждается негативной иммунохимической реакцией с использованием антител к муреину.
В то же время показано, что в периплазме покоящейся внеклеточной стадии хламидий («элементарных тел») накапливаются два белка, один из которых имеет молекулярную массу 12, а другой 60 кДа. Из-за высокого содержания остатков Cys они получили название «цистеинбогатых» белков (англ, cysteine-rich protein, CRP). Гены CRP образуют дицистронный оперон. Его анализ показал, что малый CRP содержит 68 аминокислотных остатков (из них 14 остатков Cys), а большой 517 аминокислотных остатков (из них 37 остатков Cys). Сульфгидрильные группы CRP образуют многочисленные дисульфидные мостики.
Малый CRP содержит ковалентно связанный липидный домен, заякоренный во внутреннем полу мембранном листке ОМ. По-видимому, он выполняет такую же роль, что и муреин-липопротеин Брауна (см. раздел 8.4.1.2). В свою очередь, молекулы большого CRP связаны друг с другом дисульфидными мостиками и образуют саккулус, к которому через дисульфидные мостики прикрепляются отдельные молекулы малого CRP.
Эволюционное происхождение белкового периплазматического ригидного слоя у хламидий неизвестно. Есть идея, что большие CRP ведут начало от белковых субъединиц S-слоя (см. раздел 8.5.2.3), которые в ходе эволюции потеряли способность транслоцироваться через ОМ остались в периплазматическом компартменте. Там при помощи дисульфидредуктаз из них образовался саккулус. Дополнительным селектирующим фактором послужила потеря способности к образованию пептидо- гликана.
Структура ригидного слоя хламидий изменяется в зависимости от условий внешней среды. На вегетативной внутриклеточной стадии они представляют собой «ретикулярные тела», у которых ригидный слой ослаблен в результате восстановления дисульфидных мостиков. В свою очередь, на расселительной стадии «элементарных тел» между CRP образуются многочисленные дисульфидные мостики.
Белковый периплазматический слой у Thermomicrobium. Согласно данным конца 1970-х годов, существует еще одна бактерия, содержащая периплазматический слой белковой природы. Это облигатно анаэробный гипертермофил Thermomicrobium roseum (оптимум роста 70-75°С), единственный представитель филы BVII Thermomicrobia.
В отсутствии муреина функцию клеточной стенки у этой бактерии выполняет белковый каркас. О нем известно лишь то, что он богат аланином, глицином, глутаминовой кислотой и пролином и состоит из субъединиц с молекулярной массой ~75 кДа.
8.4.4. Специализированные компартменты
Периплазматический компартмент — это мультифункциональная и динамичная транзитная зона. Обычно он мал по сравнению с цитоплазматическим компартмен- том и не годится для запасания полимеров, что потребовало бы секреции в периплазму ферментов и энергетических затрат на импорт образующихся мономеров. Видимо, по этим причинам в нем редко встречаются специализированные компартменты.
В качестве примеров рассмотрим серные глобулы и периплазматические микрокристаллы.
8.4.4.1. Серные глобулы
Серные глобулы (англ. sulfur globule) образуются у ряда хемолитотрофных и фотолитотрофных бактерий, принадлежащих к филе ВХП Proteobacteria, например, Achromatium oxaliferum, Beggiatoa alba, Chromatium vinosum и Thiocapsa roseopersicina. Они содержат серу в качестве промежуточного продукта окисления экзогенного донора электронов — сульфида. Гидратированная «элементная» сера (точнее, полисульфан Sn2-, где n>2) образуется при окислении сульфида с помощью флавоцитохрома с или сульфид:хинон-оксидоредуктазы. Активные центры этих ферментов расположены на наружной поверхности СМ. Энергия, освобождающаяся при окислении сульфида, ассимилируется в форме Рmf, и это один из примеров механизма, который называется экстрацитоплазматическим окислением «простых» субстратов (см. II том учебника).
Когда запасы сульфида во внешней среде подходят к концу, в качестве эндогенного донора электронов начинает использоваться сера, которая окисляется до сульфита, а затем до сульфата.
Серные глобулы имеют округлую форму. Их размер составляет от 100 нм до >1 мкм. Они расположены в глубоких карманах периплазматического компартмента, и создается впечатление, что прямо в цитоплазме. Окружает серную глобулу неунитарная мембрана толщиной 3-4 нм, состоящая из гидрофобных полипептидов молекулярной массой 8-11 кДа (гены sgpА, sgpВ и sgpС). Все три препротеина содержат сигнальные пептиды и секретируются в периплазматический компартмент с помощью генеральной Sес-системы (см. раздел 8.6.1). Белки SgpABC богаты глицином и гомологичны цитоскелетным кератинам и фиброину шелка. Считается, что они выполняют не ферментативную, а структурную роль.
8.4.4.2. Микрокристаллы
Выше уже шла речь о том, что семь видов квазифототрофных бактерий из класса «Alphaproteobacteria» способны восстанавливать теллурит ТеО32- до элементного теллура Те0 (см. раздел 8.3.11.5).
Кристаллы теллура образуются и в цитоплазме, и в периплазматическом ком- партменте. Во втором случае они, как правило, мельче (средний размер 8x50 нм).
Естественно, что периплазматическое восстановление теллурита в малотоксичный теллур является более эффективным способом детоксикации по сравнению с восстановлением в цитоплазме.
8.5. Экзоплазматический компартмент
В отличие от цитоплазматического и периплазматического компартментов, эк- зоплазматический компартмент не отделен от окружающей среды унитарной мембраной. Но хотя он не имеет барьера, который отделял бы его от внешней среды, это неотъемлемая часть клетки. В нем находятся следующие органеллы:
— надмембранные комплексы (у грамположительных бактерий и архей);
— экзоплазматические ригидные слои;
— придаточные структуры.
Все эти органеллы являются либо специализированными компартментами, либо внешней частью специализированного компартмента. Они не обязательно присутствуют в прокариотной клетке; некоторые встречаются эпизодически, а некоторые — в виде исключения.
8.5.1. Надмембранные комплексы у грамположительных бактерий и архей
Термин «надмембранный комплекс», или гликокаликс (греч. glycos — сладкий и лат. calyx — кожица; «гликозилированный покров») обычно используется по отношению к ядерным организмам, точнее, по отношению к животным, а также по отношению тем протистам и грибам, которые не имеют клеточной стенки. Однако этот термин можно применять и как общецитологический, когда поверхностная мембрана эукариотной или прокариотной клетки сообщается с экзоплазматическим компартментом или прямо с окружающей средой.
Надмембранные комплексы содержат гидрофобный якорный домен, который участвует в образовании мембранного матрикса, и гидрофильный домен, находящийся на экзоплазматической поверхности мембраны.
У эукариотов надмембранные комплексы являются частью СМ и представлены гликозилированными белками, а также липогликанами с короткой боковой цепью. Они отвечают в основном за сигнальную рецепцию и рецептор-обусловленный везикулярный транспорт.
В отличие от эукариотов, у прокариотов локализация надмембранных комплексов определяется морфотипом — в грамотрицательных клетках они находятся в ОМ, а в клетках, относящихся к грамположительному морфотипу или к археотному морфотипу — в СМ.
Надмембранные комплексы бактерий, обладающих грамотрицательным морфотипом, представлены ЛПС или негликозилированными липопротеинами (см. раздел 8.4.1).
В свою очередь, у бактерий, обладающих грамположительным морфотипом, и у архей надмембранные комплексы представлены:
— мембранными тейхоевыми кислотами;
— липополисахаридами, липогликанами или липоманнанами;
— негликозилированными липопротеинами и гликопротеинами.
Как и у эукариотов, важнейшей функцией надмембранного комплекса прокариотов является поверхностная рецепция. Ее специфичность зависит от структуры полигликозидного или полипептидного домена, выставленного на экзоплазматиче- скую поверхность СМ.
Кроме того, у лишенных клеточной стенки архей (р. Thermoplasma) надмембранный комплекс выполняет не свойственную ему механическую роль. Он придает СМ повышенную прочность, что необходимо для сохранения физической целостности клетки в гипотонической среде, а также дает опору базальному телу вращающегося жгутика.
8.5.1.1. Мембранные тейхоевые кислоты
Общие свойства тейхоевых кислот мы рассмотрим в разделе 8.5.2.1. Здесь же мы остановимся только на полиглицеролфосфатных тейхоевых кислотах, которые получили название «мембранных», «липотейхоевых» или «внутриклеточных».
В отличие от «стеночных» тейхоевых кислот (которые связаны с муреином ковалентно) и капсульных тейхоевых кислот (которые не связаны с другими полимерами ковалентно), мембранные тейхоевые кислоты образуют ковалентную фосфодиэфирную связь с гликолипидом, входящим в состав СМ.
Многие грамположительные бактерии одновременно содержат и стеночные, и мембранные тейхоевые кислоты. Последние более распространены, а их биосинтез меньше зависит от условий роста.
Главное различие между мембранной тейхоевой кислотой и стеночной тейхое- вой кислотой (см. раздел 8.5.2.1) заключается в том, что в первом случае донором мономера служит мембранный липид — фосфатидил-1-глицерол, и поэтому она состоит из глицерол-1-фосфата (рис. 46, А). Во втором случае донором мономера служит растворимый ЦДФ-З-глицерол, и поэтому она состоит из глицерол-3-фосфата (рис. 46, Б). Таким образом, глицеролфосфатные остатки этих биополимеров представляют собой два разных энантиомера.
Рис. 46. Изомеры глицеролфосфата, используемые при биосинтезе двух типов тейхоевых кислот. А — глицерол-1-фосфат (мембранная тейхоевая кислота); Б — глицерол-3-фосфат (стеночная тейхоевая кислота).

Цепь мембранной тейхоевой кислоты обычно состоит из 16-40 мономеров глицерол-1-фосфата. В случае Lactococcus garviae между глицеролфосфатными остатками встраивается дигалактозил. К основной цепи присоединяются различные заместители — остатки D-аланина, а также галактозы, глюкозы и N-ацетилглюкозамина. Реже это ди-, три- и тетрасахариды.
Липидный «якорь» мембранной тейхоевой кислоты имеет разное строение, хо-
тя обычно он представлен дигексозил-диацил-глицеролом (рис. 47). Негликозилиро- ванные варианты очень редки и встречаются только у некоторых штаммов Bacillus coagulans и В. megaterium.
Рис. 47. Мембранная тейхоевая кислота. X — остаток D-аланина (или — Н); глю — остаток глюкозы; R1 и R2 —остатки жирных кислот.

Биосинтез мембранных тейхоевых кислот осуществляется при помощи полимеразы на экзоплазматическом полумембранном листке СМ. Мы уже упоминали о том, что в отличие от стеночных тейхоевых кислот, донором мономера для мембранных тейхоевых кислот служит не растворимый ЦТФ-З-глицерол, а мембранный фосфатидил- 1-глицерол.
Мембранные тейхоевые кислоты полифункциональны. Прежде всего они связывают дивалентные катионы. Это обеспечивает ионный гомеостаз экстраплазматических ферментов, в частности, регулирует активность автолизинов.
Особо важна минорная мембранная тейхоевая кислота, которая в качестве липофильного носителя участвует в биосинтезе стеночных тейхоевых кислот (см. раздел 8.5.2.1). В отличие от главной фракции она содержит меньше субъединиц и не два, а целых три жирнокислотных «хвоста».
Длинные цепи мембранных тейхоевых кислот пронизывают клеточную стенку насквозь и в отдельных случаях выступают над поверхностью. Там они, по аналогии со стеночными тейхоевыми кислотами и ЛПС, проявляют комплекс адгезионных, антигенных и токсигенных свойств.
8.5.1.2. Липополисахариды и липогликаны
ЛПС и липогликаны (липоглюканы и липоманнаны) содержат липидный кор и боковую цепь. У липополисахаридов она состоит из повторяющихся олигосахаридных субъединиц, а у липоглюканов и липоманнанов — из идентичных гликозидных мономеров.
Липополисахариды и липогликаны у архей, лишенных клеточной стенки. Напомним, что уникальные липиды архей представляют собой простые ди- и тетраэфиры глицерола или кальдитола со спиртами, имеющими полиизопреноидный С15-, С20- и С25- и С40-радикал. Они образуют билипидный или квази-билипидный
слой, лежащий в основе СМ (см. раздел 8.3.3). В некоторых случаях такие липиды служат гидрофобным якорем для липоманнана.
Термофильный археот Thermoplasma acidophilum (фила АII Euryarchaeotá) лишен клеточной стенки и является аналогом бактериальных микоплазм. До 3% его клеточной массы составляет ЛПС (5,3 кДа). Боковая цепь состоит из восьми блоков (каждый из трех остатков маннозы, соединенных связями α1 —> 2, α1 —> 3 и α1 —> 4). Кор состоит из остатка глюкозы, который в положении 1 присоединяется к свободному гидроксилу полиизопреноидного тетраэфира. Благодаря тому, что у термоплазм в состав СМ входят ЛПС, она приобретает повышенную прочность, что позволяет клеткам сохраниться в агрессивной физико-химической среде. Жесткость СМ дает возможность заякоривать в ней базальные тела жгутиков, несмотря на отсутствие ригидной клеточной стенки (см. раздел 8.6.2.3).
Археотные липоглюканы, боковая цепь которых состоит из трех остатков (не)фосфорилированной глюкозы, имеются у представителей родов Pyrobaculum и Thermoproteus, а также у других гипертермофильных архей из филы AI Crenarchaeota.
Липогликаны у грамположительных бактерий, имеющих клеточную стенку. В качестве интересного примера можно привести мультидоменный полимер Bifidobacterium bifidum. Его боковая цепь состоит из двух субдоменов — дистального полигалактофуранозного и проксимального полиглюкопиранозного (связи β1 —> 4). Концевой участок образован глицерол-1-фосфатом и замещен L-аланином в положении 3. Липидный якорь представлен галактозил-диацил-глицеролом и аналогичен липидному якорю мембранной тейхоевой кислоты (см. рис. 47).
Характерными примерами бактериальных липоманнанов служат мультидоменные полимеры Micrococcus flavus, М. luteus и М. sodonensis. Боковая цепь у них образована 50-70 остатками маннопиранозы (связи α1 —> 2, α1 —> 3 и α1 —> 6), из которых 10-25% несут сукцинильные заместители. Концевой участок состоит из несукцинилированных остатков маннозы, а липидный якорь аналогичен липидному якорю мембранной тейхоевой кислоты и представлен дигалактозил-диацил-глицеролом (см. рис. 47). Судя по многочисленным анионным группам, такие липоманнаны функционально сходны с мембранными тейхоевыми кислотами.
Липополисахариды и липогликаны у мико плазм. Из всех микоплазм способностью образовывать ЛПС, по-видимому, обладает только Mycoplasma neurolyticum.
Примерами микоплазм-бактерий, содержащих липогликаны, являются Achole- plasma granularum и Anaeroplasma abactoclasticum. В отличие от микоплазм-архей (термоплазм), их гидрофобный якорный домен образован не полиизопреноидным тетраэфиром, а «обычным» диацилглицеролом.
8.5.1.3. Негликозилированные липопротеины и гликопротеины
Липопротеины удерживаются в клеточной оболочке с помощью липидного якоря. Это позволяет им выполнять разнообразные функции, находясь в иммобилизованном состоянии.
Негликозилированные липопротеины у грамположительных бактерий. В качестве примера можно привести пенициллиназу Bacillus licheniformis и Staphylococcus aureus. Этот фермент подвергается посттрансляционной модификации — подобно липопротеину Брауна он «липидируется» на своем N-конце (см. раздел 8.4.1.2). Сигнальная пептидаза, участвующая в этом процессе, так же ингибируется глобомицином.
По аналогии с грамотрицательными бактериями, у которых имеются периплазматические «связывающие» BP-белки (см. раздел 8.4.2), грамположительные бактерии содержат «связывающие» липопротеины, которые иммобилизованы на СМ. Например, у Streptococcus pneumoniae продукты оперонов mal и ami участвуют соответственно в транспорте мальтозы и олигопептидов. Дополнительными компонентами системы транспорта олигопептидов, которые не кодируются опероном ami, служат «связывающие» липопротеины АliА и АliВ. Примером «связывающего» белка с широкой субстратной специфичностью по отношению к аналогам раффинозы служит липопротеин MsmE, обнаруженный у возбудителя кариеса Streptococcus mutans.
Липопротеины грамположительных бактерий выполняют роль адгезинов. В качестве примера можно привести липопротеин SsaB (35 кДа) Streptococcus sanguis, который взаимодействует с гликопротеинами эпителиальных клеток слизистой оболочки ротовой полости человека.
Другим примером служит липопротеин PrtM (от англ. proteinase maturation protein). Он участвует в посттрансляционном процессинге протеазы. С помощью этого фермента бактерии рода Lactococcus расщепляют казеин молока на макропептиды, что приводит к образованию сырного сгустка.
Наконец, липопротеины грамположительных бактерий могут участвовать в сигнальном каскаде при дифференциации эндоспор и при конъюгации, а также выполнять структурную роль, взаимодействуя с пептидогликаном.
Гликопротеины у архей, не имеющих клеточной стенки. Гликопротеин, входящий в состав СМ археотной микоплазмы Thermoplasma acidophilum, содержит большой гликановый домен, присоединенный по остатку Asp. К дистальному концу кора, состоящего из десяти остатков галактозы, глюкозы, маннозы и N-ацетилглюко- замина прикрепляются две разветвленные полиманнозные цепи (общее число остатков маннозы — 34). Совместно с ЛПС эти гликопротеины отвечают за механическую прочность СМ, а также определяют адгезивные свойства клеточной поверхности.
8.5.2. Экзоплазматические ригидные слои у бактерий
Экзоплазматические ригидные слои у бактерий гетерогенны по строению и функциям.
Напомним, что большинство фил домена Bacteria (23 из 24) состоит из объектов, обладающих грамотрицательным морфотипом. Если у них имеется ригидный слой, то это обычно муреиновый саккулус, расположенный в периплазматическом компартменте (см. раздел 8.4.3.1).
В некоторых случаях грамотрицательные бактерии помимо муреина содержат дополнительные ригидные слои, которые находятся в экзоплазматическом компартменте. Они не только дублируют механопротекторную функцию муреинового саккулуса, но и используются для других целей. Исключение составляют представители филы BXV Planctomycetes, у которых заместителем муреинового саккулуса служит паракристаллический поверхностный слой (см. раздел 8.5.2.3).
Напомним о том, что бактерии, обладающие грамположительным морфотипом, относятся к филам ВХIII Firmicutes и BXIV Actinobacteria. За исключением микоплазм все они содержат экзоплазматический саккулус. Однако в отличие от муреинового саккулуса грамотрицательных бактерий это бинарный полимер — наряду
с муреиновым доменом в него входят другие гликановые полимеры (производные тейхоевых и тейхуроновых кислот, или тейхоаты, и производные миколовых кислот, или миколаты).
8.5.2.1. Муреин-тейхоатный саккулус
В разделе 8.4.3.1 мы уже рассматривали особенности строения муреина в эндоспорах (которые образуются у многих грамположительных и некоторых грамотри- цательных бактерий). Однако различия в строении муреина наблюдаются и между вегетативными клетками двух морфотипов — грамотрицательные и грамположительные бактерии не только образуют муреин разного молекулярного состава, но и с разным пространственным строением.
Специфика муреина у грамположительных бактерий. Помимо того, что муреин грамположительных бактерий образует несколько монослоев, он содержит большое количество пептидных сшивок — и в плоскости каждого монослоя, и между соседними монослоями. Например, 40% остатков мурамовой кислоты в вегетативных клетках Bacillus subtilisприсоединяют боковые олигопептидные цепи, причем в них 99% остатков ДАП амидируются по свободной карбоксильной группе. Боковые цепи, не участвующие в образовании поперечных сшивок, на 85% «урезаны» до трипептидов. Гликановые цепи муреина В. subtilis состоят из ~200 моносахаров, причем около 2% остатков мурамовой кислоты ковалентно связаны с тейхоевыми кислотами.
Таким образом, муреин грамположительных бактерий имеет большую толщину, содержит много сшивок и высокоригиден. В то же время он представляет собой молекулярное сито, в ячейках которого помещаются вспомогательные полимеры клеточной стенки, удерживаются ферменты и ионы. Это своеобразный квази- периплазматический компартмент, хотя в отличие от квази-периплазматического компартмента бактерий группы «mycolata» (см. раздел 8.5.2.2) он не имеет внешней билипидной границы, а в отличие от паракристаллического S-слоя (см. раздел 8.5.2.3) не построен из белковых субъединиц.
Стеночные тейхоевые кислоты. Мы уже рассматривали «мембранные» тей- хоевые кислоты (см. раздел 8.5.1.1), а в дальнейшем уделим внимание и «капсульным» тейхоевым кислотам (см. раздел 8.5.4.1).
Помимо двух этих типов тейхоевых кислот существуют также «стеночные» тейхоевые кислоты, или собственно тейхоевые кислоты (греч. teichos — стенка). Они входят в состав клеточной стенки большинства грамположительных бактерий. Некоторые грамположительные бактерии их не содержат, а у грамотрицательных бактерий они полностью отсутствуют.
Тейхоевые кислоты накапливаются в клеточной стенке в примерно равном или даже в большем количестве, чем другие компоненты. На их долю приходятся 30- 70% массы муреино-тейхоатного саккулуса (75% у Listeria monocytogenes).
Тейхоевые кислоты весьма разнообразны, и даже отдельная бактерия может образовывать несколько разных по составу и полидисперсных по длине тейхоевых кислот.
Мономерами тейхоевых кислот являются рибитол-5-фосфат (рибитолтейхоевые кислоты) или глицерол-3-фосфат (глицеролтейхоевые кислоты). Реже образуются сополимеры двух этих полиолов. Иногда в состав основной цепи тейхоевой кислоты внедряются остатки сахаров, например, галактозы. Уникальные тейхоевые кисло-
ты некоторых актиномицетов содержат четырехатомный эритритол. Общая длина тейхоевой кислоты составляет 7-40 полиольных остатков.
Через гидроксилы, не участвующие в образовании фосфодиэфирных связей, к полиолфосфатному скелету тейхоевой кислоты могут присоединяться моно-, ди-, три и тетрасахариды из остатков галактозы, глюкозы, N-ацетилгалактозамина или N-ацетилглюкозамина, связанных α- и β-гликозидными связями. Помимо глико- зильных остатков среди заместителей встречаются остатки D- или L-аминокислот, ацетила и сукцинила (рис. 48).
Рис. 48. Стеночная тейхоевая кислота. LU — линкер; ала — остаток аланина; AглюN—остаток N-ацетилглюкозамина; AманN — остаток N-ацетилманноэамина; und — ундекапренил; Ф — фосфатная группа.

Тейхоевые кислоты присоединяются к 6-му атому углерода мурамовой кислоты через линкерный участок (англ. linkage unit, LU), образованный тремя остатками 3-глицеролфосфата, остатком N-ацетил-маннозамина и остатком N-ацетилглюкоза- мина (рис. 48).
Биосинтез тейхоевых кислот. Линкерный участок собирается на ундекапре- нилфосфатном носителе из молекулы УДФ-ацетилманнозы, молекулы УДФ-N-ацетилглюкозамина и трех молекул ЦДФ-З-глицерола. В этом процессе участвуют несколько интермедиатов (липиды I — IV), и он избирательно подавляется антибиотиком туникамицином.
Отдельно от линкерного участка осуществляется поэтапная сборка полирибитол- фосфата или полиглицеролфосфата. Для этого используется «липотейхоевая кислота-переносчик» (англ. lipoteichoic acid carrier, LTC), структурно сходная с мембранной тейхоевой кислотой. Сборка осуществляется при помощи мембраносвязанной полимеразы. Донорами полиола служат ЦДФ-5-рибитол или ЦДФ-3-глицерол (рис. 49). Готовая цепь переносится на линкерный участок, который через фосфо- эфирную связь присоединяется к муреину.
Рис. 49. Транспортные формы многоатомных спиртов, используемые при биосинтезе стеночных тейхоевых кислот. А — цитидиндифосфат-рибитол; Б — цитидиндифосфат-3-глицерол; Р — фосфатная группа.
Функции тейхоевых кислот. Подобно нуклеиновым кислотам, тейхоевые кислоты имеют спиртофосфатный скелет и поэтому являются полианионами. Благодаря разнообразию боковых заместителей они приобретают свойства «информационных» молекул, которые участвуют во взаимном распознавании живых объектов, и в этом
отношении напоминают ЛПС. Как и большинство бактериальных биополимеров, тейхоевые кислоты полифункциональны:
— в качестве компонента грамположительной клеточной стенки они участвуют в ее обновлении, а также в клеточном росте и делении;
— они иммобилизуют катионы и служат ионным буфером для ферментов, связанных с клеточной оболочкой;
— являются фактором (не)специфической адгезии бактерий к живым и мертвым субстратам, что способствует образованию биопленок;
— служат рецепторами для некоторых бактериофагов;
— обладают антигенными свойствами и являются фактором патогенности; по аналогии с ЛПС в малых дозах они стимулируют иммунную систему, а в больших дозах вызывают септический шок.
Благодаря взаимному отталкиванию фосфатных групп муреин-тейхоатный саккулус расправляется и приобретает пористую структуру. Эти же группы образуют мостики с катионами. При наличии гликозильных и других заместителей вторая валентность магния используется для связывания отрицательно заряженных молекул, например, белковых адгезинов.
Тейхуроновые кислоты. При фосфорном голодании грамположительные бактерии синтезируют стеночные тейхуроновые кислоты, которые через неидентифи-
цированный линкерный участок присоединяются к мурамовой кислоте в составе муреина. Моноблок тейхуроновой кислоты Micrococcus lysodeikticus представляет собой дисахарид, состоящий из глюкозы и N-ацетил-маннозаминуроновой кислоты, которые связаны α1 —> 4 гликозидной связью. Неразветвленная цепь состоит из 10-40 моноблоков, соединенных β1 —> 6 гликозидной связью (рис. 50).
Рис. 50. Тейхуроновая кислота. LU — линкер; глю — остаток глюкозы; МанК — остаток N-ацетил-маннаминуроновой кислоты.

Полимер синтезируется из УДФ-глюкозы и УДФ-N-ацетил-маннозаминуроновой кислоты. По аналогии с тейхоевой кислотой, линкерный участок собирается на липидном носителе, которым служит ундекапренилфосфат. На нем же происходит сборка цепи тейхуроновой кислоты из дисахаридных блоков.
Относительно короткие тейхуроновые кислоты (n <100) встречаются только у грамположительных бактерий, в то время как длинноцепочечные тейхуроновые кислоты молекулярной массой ~500 кДа обнаружены и у некоторых грамотрицательных бактерий. Примером служит Е. coli, у которой они входят в состав капсулы (см. раздел 8.5.4.1).
Функции стеночных тейхуроновых кислот более узкие, чем функции тейхоевых кислот, поскольку из-за отсутствия гликозильных и иных заместителей их структура однообразна.
8.5.2.2. Муреин-миколил-арабиногалактановый саккулус. Особенности клеточной оболочки у бактерий группы «туcolata»
В сборную группу «туcolata» входят представители восьми родов из филы BXIV Actinobacteria — Corynebacterium, Dietzia, Gordona, Mycobacterium, Nocardia, Rhodococcus, Skermaniaи Tsukamurella. Среди них есть как сапрофиты, так и патогены животных, поселяющиеся в макрофагах, в частности М. tuberculosis, которая инфицирует ~30% населения Земли и ежегодно убивает ~3 млн людей. Микобактерии не образуют токсинов, и вызываемые ими патологии связаны с неадекватным иммунным ответом хозяина.
Традиционные возбудители туберкулеза и проказы (М. tuberculosis и М. leprae) устойчивы к неблагоприятным физико-химическим условиям, а также к «обычным» антибиотикам и химиотерапевтическим препаратам. Это касается и сапрофитных форм, которые поражают лиц с нарушенной иммунной системой, особенно если они инфицированы вирусом HIV (М. kansasii, М. chelonae и др.). В первую очередь это
объясняется очень слабой проницаемостью клеточной оболочки. Тем не менее, бактерий группы «mycolata» подавляет аминогликозидный антибиотик рифампицин и химиопрепарат изониазид.
Генеральный план строения клеточной оболочки. Клеточная оболочка «mycolata» уникальна.
К муреиновому саккулусу ковалентно присоединяется гетерополимер арабино- галактан, этерифицированый «миколовыми» кислотами (англ. mycolic acid). В результате этого возникает тройственный муреин-миколил-арабиногалактановый саккулус.
Периферические углеводородные цепи миколовых кислот образуют липидный монослой, на поверхности которого лежит второй монослой, состоящий из «свободных» липидов и гликолипидов со встроенными белками. Таким образом, у «mycolata» есть билипидная мембрана, аналогичная ОМ. Такая квази-ОМ является внешней границей квази-периплазматического компартмента (рис. 51).
Рис. 51. Строение клеточной оболочки у бактерий из группы «mycolata». СМ — цитоплазматическая мембрана; квази-ОМ — квази-наружная мембрана; LM — липоманнан; LAM — липоарабиноманнан; LU —линкер; рам — остаток рамнозы; АглюN — остаток N-ацетилглюкозамина; Р — фосфатная группа; ТМНА — тетрамиколил-гексаарабинозид; CF — корд-фактор; SL — сульфолипид; PGL — фенольный гликолипид; О-рам — остаток оксиметилрамнозы; фен — фенил; TAG — триацилглицерол; LOS — липоолигосахарид; osc — олигосахаридная боковая цепь; GPL — гликопептидолипид; PIM — фосфоинозитол-диманнозид.

Клеточная оболочка «mycolata» исключительно сложна и вариабельна, что связано с составом ее компонентов и их взаимодействием. Хотя она обогащена липидами, ее наружная часть в основном содержит полисахариды и белки.
Надмембранный комплекс: липоманнаны и липоарабиноманнаны. Липоманнаны и липоарабиноманнаны (англ. lipomannan, LM; lypoarabinomannan, LAM) — еще один пример надмембранных комплексов, которые мы уже рассматривали в разделе 8.5.1.2.
Эти полимеры относятся к семейству фосфатидилглицерол-маннозидов. У них имеется кор, состоящий из мембранного липида фосфатидилинозитола и двух остатков маннозы (англ, phosphatydylinositol dimannoside, PIM2).
Основу липоманнана образует линейная полигликозидная цепь из остатков ман- нопиранозы, которые соединены связями α1 —> 6. В отдельных местах через связи α1 —> 2 к ней присоединяются боковые остатки маннозы (рис. 51).
В свою очередь, липоарабиноманнаны состоят из проксимальной маннановой части и дистальной арабинановой части. Последняя содержит кэп из двух остатков маннозы, который при фагоцитозе взаимодействует с маннозным рецептором.
Липоманнаны и липоарабиноманнаны являются функциональной альтернативой тейхоевых кислот (см. раздел 8.5.2.1). Они тоже проявляют биологическую активность, в частности выступают в качестве иммуномодуляторов и ключевых лигандов при фагоцитозе. Кроме того, они выполняют важную роль в вирулентности этих бактерий.
Особенности муреина у бактерий группы «mycolata». Почти у всех «mycolata», исключение составляет род Corynebacterium, мурамовая кислота содержит в N-положении не ацетил, а остаток гликолевой кислоты СН2(ОН)СООН. Появление дополнительного гидроксила не может не сказаться на системе водородных связей внутри саккулуса, хотя конкретный результат этой модификации не ясен. Вторая особенность муреина «mycolata» — это то, что боковые пептидные цепи могут быть соединены как через остатки ДАП и Dала, так и через остатки ДАП.
Связанные липиды: миколил-арабиногалактан. Эти липиды называются «связанными», поскольку они составляют неотъемлемую часть саккулуса, и их нельзя экстрагировать органическими растворителями. Арабиногалактановый домен содержит неразветвленную цепь из ~30 остатков галактофуранозы, поочередно соединенных связями α1 —> 5 и α1 —>6. При помощи линкерной единицы, состоящей из рамнопиранозы и ацетилглюкозамина, галактановая цепь через фосфоэфирный мостик присоединяется к 6-му атому углерода у 10-12% остатков мурамовой кислоты. К галактановой цепи прикреплены разветвленные арабинановые цепи из 10-20 остатков арабинофуранозы, соединенных связями α1 —> 3 и α1 —> 5 (рис. 51).
На концах арабинановых цепей расположены три липидных домена. Каждый из них представляет собой тетрамиколил-гексаарабинозид (англ. tetramycolyl hexoarabinoside, ТМНА). Его углеводный каркас состоит из остатков арабинофуранозы, соединенных связями α1 —> 3, α1 —> 5, β1 —> 2 и β1 —> 5. Четыре боковых остатка гексаарабинозного блока этерифицированы в положении 5 остатками «миколовых» кислот (рис. 52).
Рис. 52. Тетрамиколил-гексаарабинозид. ара — остаток арабинозы.
Миколовые кислоты — это β-гидроксикислоты с двумя алкильными радикалами (рис. 52). Более консервативный радикал из 20-25 атомов углерода занимает α-положение. Более вариабельный радикал из 50-65 атомов углерода, или «меромиколат» (англ. meromycolate) занимает β-положение. В нем имеются двойные связи, и он содержит циклопропановые и эпокси-кольца, а также метильные, кето- и метокси- группы.
Как правило, миколовые кислоты представляют собой не индивидуальные соединения, а набор гомологов (числом до 15). Биосинтез миколовых кислот, составляющих 30-40% массы клеточной оболочки — серьезная нагрузка на метаболизм, особенно для медленно растущих форм, время генерации которых свыше 24 ч. Поэтому эндосимбиотические микобактерии для образования липидов используют жирные кислоты, которые они получают от макроорганизма. Биосинтез миколовых кислот ингибируется изониазидом (англ. isoniazid, INH), который действует на меромиколат-синтазу и поэтому является сильным противотуберкулезным препаратом.
Свободные липиды. «Свободными» эти липиды называются потому, что не образуют ковалентной связи с муреин-арабиногалактановым саккулусом. Их можно экстрагировать органическими растворителями, и они подразделяются на полярные и неполярные. Как уже отмечалось, на основе свободных липидов, с внешней стороны, и тетрамиколил-гексаарабинозида, с внутренней стороны, образуется би- липидная мембрана (квази-ОМ).
Главные представители свободных полярных липидов содержат нередуцирующий дисахарид трегалозу, которая состоит из двух остатков глюкозы, связанных связью α1 —> 4. Их характерным примером служит 6,61-димиколат трегалозы (англ. trehalose dimycolate, TDM), или «корд-фактор» (англ. cord factor, CF). Смысл названия заключается в том, что димиколат трегалозы одно время считали главным фактором вирулентости М. tuberculosis; его присутствием также объясняли рост на поверхности жидкой питательной среды в виде «тяжей» (англ. cord — шнур). В дальнейшем корд-фактор был найден как у сапрофитных микобактерий, так и у не образующих тяжей патогенных микобактерий. Это вещество используется в качестве компонента «неполного», т. е. без добавления авирулентных клеток М. tuberculosis, адъюванта Фрейнда (J. Freund), который действует как сильный иммуностимулятор. Помимо димиколата трегалозы существует еще и мономиколат трегалозы (англ. trehalose monomycolate, ТММ), который является транспортной формой миколовых кислот.
Еще одним примером свободных полярных липидов, в составе которых есть трегалоза, служит сульфолипид SL (от англ. sulfolipid). Наряду с остатками «обычной» Cíe-пальмитиновой кислоты он содержит три остатка уникальной микоцерозной кислоты (англ, mycocerosic acid), гидрофобный «хвост» которой построен из изопропильных блоков (рис. 53). В связи с этим напомним, что множественное внутреннее ветвление липидных «хвостов» не свойственно бактериям, зато характерно для уникальных липидов архей (см. раздел 8.3.3.2).
Рис. 53. Сульфолипид (SL) и фенольный гликолипид (PGL). тре — остаток трегалоэы; О-рам — остаток оксиметилрамнозы.

Помимо тетраацил-сульфолипида SL, к числу гликолипидов на основе трегалозы принадлежат диацилтрегалоза DAT (от англ. diacyl trehalose), триацилтрегалоза TAT (от англ. triacyltrehalose) и пентаацилтрегалоза РАТ (от англ. pentaacyl trehalose).
Другим примером свободных полярных липидов служат фенольные гликолипиды (англ. phenolic glycolipids, PGL). Их гликозидная часть представлена О-метил- рамнозой. Через фенильную группу она связана с липидным доменом, который состоит из длинноцепочечного воскообразного спирта фтиоцерола (англ. phthiocerol). Фтиоцерол относится к двухатомным спиртам, или гликолям. Обе его гидроксильные группы этерифицированы микоцерозной кислотой (рис. 53).
Неполярные свободные липиды, представленные триацилглицеридами (англ. triacylglyceride, TAG), образуют минорную фракцию свободных липидов. В их состав входят жирные кислоты с алифатическими радикалами средней длины.
Ацилированные производные трегалозы (фтиоцерол-димикоцерозаты и три- ацилглицериды) однообразны по структуре и не отвечают за специфическую физиономию клеточной поверхности бактерий из группы «туcolata». Этим они отличаются от высоко специфичных липополисахаридов и гликопептидолипидов.
Липоолигосахариды (англ. lipooligosaccharide, LOS). Совместно со свободными липидами они образуют экзоплазматический полумембранный листок квази-ОМ. Представители данной группы гликолипидов состоят из трегалозы. Первый глюкозный остаток этерифицирован двумя длинноцепочечными жирными кислотами, а со вторым глюкозным остатком связана боковая олигосахаридная цепь (англ. oligosaccharide side chain, OSC). Она состоит из мономеров глюкозы, к которым в некоторых случаях присоединяются остатки пирувата.
Гликопептидолипиды (англ. glycopeptidolipide, GPL). Эти соединения также входят в состав экзоплазматического полумембранного листка квази-ОМ. Они являются одними из важнейших поверхностных антигенов бактерий из группы «mycolata». Тетрапептидный кор GPL состоит из остатков D-фенилаланина, D- треонина и D-аланина, а также остатка аминоспирта D-аланинола, с которым связан остаток О-диметил-L-рамнозы. К гидроксильному радикалу треонина присоединяется олигосахаридная цепь. Она состоит из остатков L-рамнозы и дезокситалозы, а аминогруппа фенилаланина несет остаток α-гидроксилированной жирной С28-кислоты.
С накоплением GPL связывают способность патогенных микобактерий модулировать иммунный ответ макроорганизма путем взаимодействия с поверхностью его клеток, а также персистировать, т. е. устойчиво сохраняться внутри макрофагов.
Квази-наружная мембрана. Под электронным микроскопом в оболочке «mycolata» видны чередующиеся зоны с высокой и низкой электронной плотностью, что внешне напоминает оболочку грамотрицательных бактерий. Действительно, помимо СМ клетка окружена вторым билипидным барьером — квази-ОМ.
Структурно-функциональное сходство между «mycolata» и грамотрицательными бактериями на самом деле существует, однако оно носит конвергентный характер, поскольку квази-ОМ не содержит «обычных» ЛПС, «обычных» поринов и липопротеина Брауна. Кроме того, между СМ и квази-ОМ не образуются зоны адгезии (контакты Байера). Подобно ОМ грамотрицательных бактерий, квази-ОМ ковалентно связана с муреином. Однако связь осуществляется не за счет полипептидной цепи липопротеина Брауна, а с помощью полигликозидной цепи миколил-арабиногалактана.
И все же квази-ОМ является самой настоящей унитарной мембраной. Углеводородные цепи миколовых кислот ориентируются перпендикулярно клеточной поверхности и благодаря своей плотной упаковке образуют цитоплазматический полу- мембранный листок. Экзоплазматический полумембранный листок состоит из свободных липидов, LOS и GPL (см. рис. 51). Поскольку миколовые кислоты намного превосходят по длине другие жирные кислоты, толщина квази-ОМ максимальна для унитарной мембраны (9-10 нм).
Квази-ОМ обладает низкой текучестью. Температура ее плавления превышает 70°С, в отличие от мембран мезофилов, которые обычно плавятся при -10°С (см. раздел 8.3.3.1). Латеральная диффузия молекул внутри квази-ОМ также ограничена. Таким образом, квази-ОМ представляет собой прочный гидрофобный барьер, который задерживает гидрофильные соединения. В то же время через него могут диффундировать гидрофобные субстраты, в том числе гидрофобные антибиотики.
Квази-ОМ бедна белками, и их функции в большинстве случаев неизвестны. В качестве примера можно привести VapA — минорный липопротеин Rhodococcus equi. В отличие от обычных липопротеинов, он не имеет N-терминального цистеинового мотива (см. раздел 8.4.1.2).
Проблема транспортировки гидрофильных соединений через квази-ОМ разрешается с помощью поринов.
Порины квази-наружной мембраны. В настоящее время описаны два белка, образующие гидрофильные поры в квази-ОМ. Это низкоактивный канал OmpATb и порин MspA.
Белок OmpATb гомологичен белку ОmрА грамотрицательных бактерий. С его помощью эндосимбиотическая бактерия М. tuberculosis экспортирует гидрофильные основания, в частности катион аммония, что позволяет ей бороться с высокой кислотностью внутри фагосом у макрофагов.
Порин MspA (100 кДа), имеющийся у М. smegmatis, не гомологичен поринам грамотрицательных бактерий. Он обладает бочковидной структурой и помимо антипараллельных β-цепей содержит 10% α-спиральных участков. Тетрамер этого белка образует конусовидную пору длиной 10 нм, через которую проходят разнообразные гидрофильные молекулы, в том числе питательные субстраты. В квази-ОМ М. tuberculosis и М. bovis содержатся, по крайней мере, два порина, не гомологичные порину MspA.
Квази-периплазматический компартмент бактерий из группы «mусоlata». В этом компартменте (см. рис. 51) находятся надмембранный комплекс и саккулус, а также квази-периплазма, содержащая растворимые белки, малые молекулы и ионы. В отличие от периплазмы у грамотрицательных бактерий (см. раздел 8.4.2), состав квази-периплазмы не детализирован. Однако ясно, что среди иммобилизованных ферментов должны быть гликозилтрансферазы и транспептидазы, с помощью которых синтезируется материал клеточной стенки.
8.5.2.3. Паракристаллический поверхностный слой
В экзоплазматическом компартменте бактерий из разных фил, обладающих как грамотрицательным, так и грамположительным морфотипом, находится белковый ригидный слой. Его называют «паракристаллическим поверхностным слоем», «поверхностным S-слоем», или просто «S-слоем» (от англ. surface — поверхность).
Хотя S-слой встречается довольно часто (он выявлен примерно у 350 штаммов, относящихся к 100 родам), преобладающее большинство бактерий его не образует. Напротив, у архей это почти универсальный и часто единственный компонент клеточной оболочки (см. раздел 8.5.3.1).
Следует подчеркнуть, что у бактерий S-слой очень редко является единственным ригидным слоем. Обычно он сосуществует с муреиновым саккулусом, и поэтому одно время его называли «A-слоем» (от англ. additional — вспомогательный). Как правило, он не выполняет формообразующей роли, и его присутствие или отсутствие не сказывается на внешнем виде клетки. При росте в лабораторных условиях бактерии часто теряют способность образовывать S-слой.
У грамотрицательных бактерий S-слой обычно существует наряду с пептидогли- кановым саккулусом, а у грамположительных бактерий — с пептидогликан-тейхоатным саккулусом или пептидогликан-арабиногалактан-миколатным саккулусом.
Строение S-слоя. По современным представлениям, S-слой — простейший тип биологической мембраны, которая образуется путем самосборки из идентичных белковых субъединиц. Возможно, что именно такая мембрана была у прогенота — общего предка клеточных организмов (см. раздел 5.3), и от нее произошли современные S-слои, а также унитарные протеолипидные мембраны.
Субъединицы S-слоя, или протомеры секретируются в экзоплазматический компартмент. Затем они спонтанно агрегируют за счет образования гидрофобных, электростатических и водородных связей. В результате образуется физически и химически стабильная паракристаллическая структура с квадратной, гексагональной или скошенно-трапециевидной симметрией. Толщина S-слоя составляет 5-15 нм. Он пронизан регулярно расположенными одинаковыми порами диаметром 2-6 нм, которые занимают 30-70% клеточной поверхности. В некоторых случаях у бактерий образуется несколько S-слоев, которые расположены друг над другом. Они имеют идентичный или разный состав и тип симметрии.
Протомеры S-слоя составляют значительную часть общего белка (до 15%) и синтезируются в большом числе копий (5 • 105), в результате чего укрывают всю поверхность клетки. У разных бактерий протомеры S-слоя обнаруживают слабую взаимную гомологию, или она отсутствует. У грамположительных бактерий они содержат консервативный мотив, или «домен гомологии S-слоя» (англ. S-layer homology domain, SLH), который взаимодействует с муреином. В ходе эволюции он включался в самые разные белки оболочки, например, у Thermotoga maritima он входит в состав периплазматического спейсера мембранного белка Ompct.
На проксимальном конце S-белка грамотрицательных бактерий находится «анкерный» домен, с помощью которого он интегрируется в ОМ. Чаще всего это происходит путем образования кальциевых мостиков с ЛПС. В свою очередь, у бактерий группы «mycolata» (см. раздел 8.5.2.2) протомеры S-слоя могут заякориваться и в наружном гидрофобном слое клеточной стенки.
Молекулярная масса протомеров S-слоя составляет 40-200 кДа. Большинство из них — кислые белки, содержащие относительно много гидрофобных и крайне мало серусодержащих аминокислотных остатков (или их нет вообще). Как правило, S- белки содержат большой фрагмент с апериодической структурой, 20-35% ß-цепей и 2-15% α-спиралей.
Во многих случаях протомеры S-слоя гликозилированы, причем углеводный домен по структуре напоминает О-антиген. Это линейная или разветвленная цепь, повторяющиеся гликозидные звенья которой состоят из 2-6 моносахаридов, связанных между собой а- и ß-гликозидными связями 1 —> 2, 1 —> 3, 1 —> 4 и 1 —> 6. Среди мономеров встречаются обычные гексозы — галактоза, глюкоза, манноза и рамноза, а также редкие N-ацетилированные и О-метилированные сахара, дезоксисахара и уроновые кислоты, которые могут дополнительно фосфорилироваться или сульфатироваться. Углеводная цепь содержит от нескольких до 150 моносахаридов и связана с полипептидным доменом 0-гликозидной связью (через остаток Ser или Туг) или уникальной N-гликозидной связью (через остаток Asn).
Благодаря гликозилированию протомеров S-слой приобретает термостабильность. Но, что еще важнее, это обеспечивает широкий спектр комплементарных взаимодействий с поверхностными структурами других живых объектов, от вирусов до дифференцированных клеток высших позвоночных.
Биогенез S-слоя. Гены протомеров обычно представлены единственной копией. Поскольку S-слой собирается со скоростью 500 субъединиц • с-1, они должны иметь очень сильные промоторные участки. Как правило, протомеры экспортируются с помощью генеральной Sec-системы, хотя известны случаи, когда их транспорт обеспечивает ABC-транспортер (см. II том учебника). Интересно, что протомеры почти не обновляются и редко сбрасываются в окружающую среду.
Как и в случае ЛПС, звенья углеводной цепи синтезируются из моносахаров, активированных с помощью нуклеотидов, а затем передаются полиизопреноидному носителю. В данном случае это не «бактериальный» ундекапренилфосфат, а «эукариотный» долихолилфосфат. Полимеризация углеводной цепи, по-видимому,
происходит за пределами цитоплазматического компартмента и ингибируется антибиотиком бацитрацином. Полностью сформированная гликановая цепь передается независимо экспортируемому протомерному белку.
Структурные перестройки S-слоя. Когда S-слой является единственным ригидным слоем, от него — по аналогии с муреином — зависит структурная целостность клетки. В то же время при росте и делении он должен раскрываться, чтобы в него могли встраиваться новые субъединицы. Вероятно, это происходит путем локального протеолиза, хотя конкретные сведения об этом отсутствуют.
Функции S-слоя. Ригидный S-слой дублирует муреиновый саккулус, но этим его функции не ограничиваются. В частности, он может выполнять роль:
— механопротекторной и формообразующей структуры;
— фильтра для экзогенных молекул;
— ловушки для катионов и секретируемых ферментов;
— фактора адгезии к гомологичным и гетерологичным клеткам, а также мертвым субстратам;
— рецептора для бактериофагов;
— матрикса, обеспечивающего прижизненную и постмортальную биоминерализацию.
Благодаря тому, что S-слой содержит поры с большим эксклюзивным диамером (30-45 кДа), он может иммобилизовывать ферменты и, таким образом, играть роль квази-периплазматического компартмента. У патогенных бактерий S-слой является фактором вирулентности. С одной стороны, он маскирует иммуногенные эпитопы и защищает клетку от иммуноглобулинов и других белков иммунной системы. С другой стороны, он выполняет роль адгезина и обеспечивает прикрепление бактерий к макрофагам (примерами служат Aeromonas salmonicida — патоген рыб, а также Campylobacter fetus — возбудитель сепсиса и менингита у новорожденных).
С помощью S-слоя многие виды бактерий механически защищаются от хищной бактерии Bdellovibrio bacterovorus, а другие виды путем неспецифической адгезии колонизируют мертвые субстраты.
Особо важны матричные свойства S-слоя цианобактерий. На нем формируются нерастворимые минералы кальция и магния, что на протяжении геологических периодов приводило и продолжает приводить к формированию осадочных пород.
Высокая упорядоченность S-слоев делает их перспективным материалом для нанотехнологии. Они могут использоваться в качестве биосенсоров, матриц для иммобилизации ферментов и антигенов, а также ультрафильтров.
8.5.3. Экзоплазматические ригидные слои у архей
Все археи, за исключением представителей рода Thermoplasma, обладают ригидной стенкой. Как правило, это паракристаллический поверхностный слой, или белковый S-слой. В редких случаях с ним сочетаются (или его подменяют) ригидные слои на гликозидной основе — псевдомуреин и кислый сульфатированный гетерополисахарид.
Археи могут окрашиваться по Граму как положительно, так и отрицательно, что зависит от толщины их ригидного слоя. Отрицательное окрашивание дают археи, образующие только S-слой. Положительное окрашивание дают палочковидные метаногены, образующие толстый (15-25 нм) псевдомуреиновый саккулус, и экстремально галофильные кокки со стенкой из сульфатированного гетерополисахарида.
8.5.3.1. Паракристаллический поверхностный слой
Археи чаще всего содержат единственный S-слой, хотя в некоторых случаях два S-слоя расположены один поверх другого. Протомеры единственного или нижнего S- слоя заякорены в CM. Halo bacterium salinarium, например, с этой целью использует гидрофобный С-концевой домен длиной в 21 аминокислоту.
Однако у нас нет оснований рассматривать S-слой у архей как надмембранный комплекс (см. раздел 8.5.1). В отличие от последнего, это паракристаллическая структура, которая высоко упорядочена и ригидна. У архей S-слой чаще всего выполняет роль клеточной стенки, тогда как у бактерий это бывает довольно редко (см. раздел 8.5.2). Характерно, что под воздействием антибиотика бацитрацина, ингибирующего гликозилирование белков S-слоя, клетки Halobacterium sp. теряют палочковидную форму. В свою очередь, клетки Sulfolobussp. содержат полуригид- ный S-слой и поэтому плеоморфны.
S-слои у архей дополнительно выполняют функции:
— молекулярного сита;
— внешней границы квази-периплазматического компартмента (см. ниже);
— фактора адгезии к живым клеткам и (не)органическим субстратам;
— матрицы для биоминерализации в результате осаждения солей кальция и магния.
Субъединицы S-слоя у архей устроены сложнее, чем у бактерий. Обычно они состоят из нескольких полипептидных доменов, часть которых имеет нитевидную форму. Главная особенность S-слоя архей состоит в том, что при помощи спейсеров между ним и СМ создается промежуток шириной 8-70 нм. Такие столбообразные домены поддерживают пористый купол из взаимосвязанных дистальных доменов, который образует наружную границу квази-периплазматического компартмента. Это функциональный аналог периплазматического компартмента грамотрицательных бактерий (см. раздел 8.4), квази-периплазматического компартмента бактерий из группы «mycolata» (см. раздел 8.5.2.2) и квази-периплазматического компартмента у бактерий, образующих пористый S-слой (см. раздел 8.5.2.3). Отверстия S-слоя у архей имеют очень большой диаметр (до 12 нм), и поэтому внутри квази-периплазматического компартмента удерживаются только белки молекулярной массой >35 кДа. К их числу относятся секретируемые протеазы, нуклеазы и гликозидазы.
Особенности состава и биогенез S-слоя. Характерная особенность S-слоя у архей — это доминирование неполярных аминокислот над полярными, причем среди последних преобладают аминокислоты с анионными радикалами.
Белки S-слоя у архей часто гликозилируются. Иногда гликановых цепей много, например, Н. salinarium содержит цепи трех типов, которые связаны с белком N-гликозидной связью через остатки Asn и Thr. Основу большой гликановой цепи Н. salinarium образует линейный кор (10-15 субъединиц, состоящих из N-ацетилглюкозамина, N-ацетилгалактозамина и алактуроновой кислоты), к которому добавляются остатки галактофуранозы, О-метил-галактуроновой кислоты и до 50 сульфатных групп. Гликановая цепь среднего размера (10 копий) содержит три сульфатированных остатка глюкуроновой кислоты и один остаток глюкозы.
Малая гликановая цепь (20 копий) состоит из нейтральных остатков галактозы и глюкозы.
Гликановый домен S-белка Methanothermus fervidus устроен гораздо проще и состоит из трисахарида маннозы, к которому присоединяются два остатка О-метил- маннозы и остаток N-ацетил-галактозамина. Как и при биосинтезе других глика- нов, остатки сахаров связываются с нуклеотидным переносчиком (УДФ), однако в качестве следующего, липофильного носителя используется не «прокариотный» С55- ундекапренилфосфат, а «эукариотный» С60-долихолилфосфат (см. раздел 8.4.1.1). В отличие от эукариотов, гликановая цепь собирается из повторяющихся субъединиц на липофильном носителе в цитоплазме, а затем переносится на предварительно экспортированный полипептид. В отдельных случаях процессинг гликана продолжается на гликопротеине.
Особенности структуры S-слоя. Степень упорядоченности S-слоя у архей варьирует от вида к виду. Такие палочковидные формы, как Pyrobaculum sp. и Thermoproteus sp. образуют почти идеальную паракристаллическую решетку. У плеоморфного Sulfolobus sp. решетка имеет многочисленные дефекты, в то время как у Desulfurococcus sp. упорядоченность практически отсутствует.
Форма субъединиц S-слоя, а также их доменный состав существенно различаются. Примером наиболее сложно организованных субъединиц, из которых образуется S-слой, служит «тетрабрахий» (англ. tetrabrachion; греч. tetra — четыре и brachion — рука; «четырехручник»), имеющийся у Staphylothermus marinus. По виду он напоминает парашютик одуванчика и состоит из заякоренного в СМ стебля длиной 70 нм и диаметром 5 нм. Посередине к стеблю прикрепляются две глобулярные частицы диаметром 10 нм, а на конце — четыре «руки» длиной 24 нм и диаметром 5 нм (рис. 54). Стебель состоит из четырех параллельных α-спиральных субъединиц общей молекулярной массой 150 кДа, а каждая рука — из четырех антипараллельных (3-цепей общей молекулярной массой 85 кДа. Глобулярная частица (150 кДа) — это «сериновая» протеаза, которая не имеет отношения к поддержанию структуры тетрабра- хия, а выполняет в квази-периплазмати- ческом компартменте функцию пищеварительного фермента.
Рис. 54. Клеточная оболочка археота Staphylothermus marinus. СМ — цитоплазматическая мембрана; Р — протеаза.
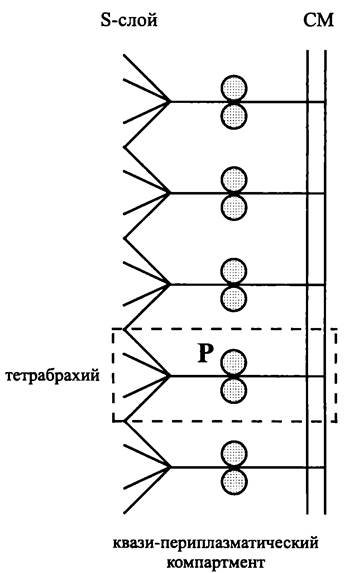
Устойчивость S-слоя в экстремальных условиях. S-слои у архей, занимающих экстремальные ниши, устойчивы при высоком содержании солей, температуре около 100°С, низких значениях pH и высоком гидростатическом давлении. Термостабильность белков S-слоя объясняется их посттрансляционной модификацией, прежде всего путем гликозилирования, а также образованием множественных гидрофобных связей и солевых мостиков.
8.5.3.2. Псевдомуреиновый саккулус
У мезофильных метаногенных архей из родов Methanobacterium, Methanobrevibacter и Methanosphaera клеточной стенкой служит псевдомуреиновый саккулус. Экстремально термофильные археи Methanopyrus kandleri и Methanothermus fervidus в дополнение к нему содержат паракристаллический поверхностный слой.
Псевдомуреин (англ. рseudomurein) — это археотный вариант пептидогликана. Подобно бактериальному муреину, он состоит из полигликозидных цепей, к которым присоединяются связанные друг с другом боковые олигопептидные цепи (рис. 55). Однако псевдомуреин в отличие от муреина не содержит мурамовой кислоты, D- аминокислот и D, L-диаминопимелиновой кислоты.
Рис. 55. Строение псевдомуреина. АглюN — остаток N-ацетилглюкозамина; ТалК — остаток N-ацетилталоэаминуроновой кислоты; глу, ала, лиз и ори — остатки глутаминовой кислоты, аланина, лизина и орнитина.
Гликановая часть псевдомуреина является сополимером N-ацетилглюкозамина и N-ацетил-талозамину роновой кислоты (α-L-2-N-ацетиламино-2-дезокси-талуроновой), которые соединены связями β1—> 3. Боковыми цепями служат три- или тетрапептиды. Они характеризуются тем, что:
— в них входят только L-аминокислоты (L-глутаминовая, L-аланин, L-лизин);
— они содержат «необычные» пептидные связи (за счет у-карбоксильной группы L-глутаминовой кислоты и ε-аминогруппы L-лизина).
Как и в случае муреина, встречаются разные варианты строения псевдомуреина. Например, N-ацетилглкжозамин в главной гликановой цепи может быть частично или полностью заменен N-ацетилгалактозамином. В боковой пептидной цепи место проксимальной глутаминовой кислоты может занять аспарагиновая кислота, а место аланина — треонин или серин; α-карбоксильная группа концевых остатков глутаминовой кислоты может связывать δ-аминогруппу орнитина, а α-аминогруппа лизина — у-карбоксильную группу глутаминовой кислоты.
Путь биосинтеза псевдомуреина кардинально отличается от пути биосинтеза муреина (см. раздел 8.4.3.1). Это касается, прежде всего, биосинтеза дисахаридного блока. Для его образования используется УДФ-активированная форма N-ацетилглюкозамина, а не обоих мономеров, как в случае муреина. Второй мономер, N-ацетилталозаминуроновая кислота образуется окислительным путем на уровне УДФ-активированного дисахарида, состоящего из N-ацетилглюкозамина и продукта его эпимеризации — N-ацетилгалактозамина.
В свою очередь, боковая пептидная цепь собирается уникальным способом (рис. 56). В качестве носителя используется не УДФ-активированное производное глюкозы (N-ацетил-мурамовая кислота), а активированная аминокислота (глутаминовая), которая через свою а-аминогруппу присоединяется к УДФ. После образования ди-, три-, тетра- и пентапептида он переносится на УДФ-активированный блок, состоящий из остатков N-ацетилглюкозамина и N-ацетилталозаминуроновой кислоты. На следующей стадии УДФ заменяется липофильным переносчиком. Его роль, как и при биосинтезе муреина, выполняет ундекапренилфосфат (см. рис. 35). В такой форме дисахарид-пентапептид транслоцируется на наружную поверхность СМ, где встраивается в псевдомуреиновый саккулус. По аналогии со сборкой муреина, источником энергии для транспептидазной реакции служит гидролиз пептидной связи между концевым L-аланином и L-лизином, а для трансгликозилазной реакции используется энергия расщепления пирофосфатной связи.
Рис. 56. Биосинтез блока-мономера и его встраивание в псевдомуреиновый саккулус. УДФ — уридиндифосфат; УМФ— уридинмонофосфат; АглюN — остаток N-ацетилглюкозамина; ТалК — остаток N-ацетилталозаминуроновой кислоты; глу, ала и лиз — остатки глутаминовой кислоты, аланина и лизина; Р — фосфатная группа; 1 — эпимераза; 2 — транспептидаза.Гребенкой обозначен ундекапренил.

Псевдомуреин более устойчив к биологическим воздействиям по сравнению с муреином. Поскольку он не содержит концевой последовательности Dала-Dала, его биосинтез не ингибируется D-циклосерином и такими антибиотиками, как пенициллин и ванкомицин (см. раздел 8.4.3.1). А так как боковые пептидные цепи содержат у- и ε-связи, он защищен и от действия пептидаз.
Несмотря на внешнее сходство с муреином, он не мог произойти от него в результате серии мутаций. Эти пептидогликаны имеют конвергентное происхождение, т. е. археи независимо от бактерий «изобрели» свой саккулус.
8.5.3.3. Гетерополисахаридный ригидный слой
Клеточная стенка экстремально галофильного археота Halococcus morrhuae имеет толщину 50-60 нм и состоит из гетерополисахарида. Этот полимер уникален: в числе прочих, он содержит два «необычных» мономера — N-ацетилгулозаминуроно- вую кислоту и N-глицил-глюкозамин. Все остатки уроновых кислот и часть остатков нейтральных сахаров сульфатированы.
Субъединица гетерополисахарида Н. morrhuae состоит из трех доменов, из которых первый и второй соединяются глициновым мостиком (рис. 57). Неясно, можно ли рассматривать такой гетерополисахарид как саккулус, или он состоит из нековалентно связанных субъединиц. Ведь для образования саккулуса необходимо, чтобы кроме интердоменных глициновых мостиков еще существовали мостики между отдельными субъединицами.
Рис. 57. Гетерополисахарид клеточной стенки археота Halococcus morrhuae. AгaлN — остаток N-ацетил галактозамина; глю, гал, гли и ман — остатки глюкозы, галактозы, глицина и маннозы; ГулК — остаток N-ацетилгулозаминуроновой кислоты; АглюN — остаток N-ацетилглюкозамина.
Ригидный слой других галококков (например, алкалифила Natronococcus occultus) состоит из уроновых кислот, N-ацетилглюкозамина и глутаминовой кислоты, однако его первичная структура еще не реконструирована.
8.5.4. Специализированные компартменты
В данную группу входят микрокомпартменты, которые сильно различаются по строению и функциям; Их часто называют придаточными структурами клетки (англ. cell appendage), чем подчеркивается их поверхностная локализация.
Придаточные структуры клетки можно разбить на две основные группы:
1. Покровы (англ. outermost investment);
2. Поверхностные органеллы (англ. surface organelle).
Как любой специализированный микрокомпартмент, покровы и поверхностные органеллы вносят вклад в дела «домашнего хозяйства». Однако для вегетативной, т. е. растущей и размножающейся клетки они не обязательны, и обладание ими лишь создает адаптационные преимущества.
Покровы и поверхностные органеллы имеют существенное значение для симбиотических бактерий, в особенности для патогенов, у которых капсула является одним из факторов вирулентности.
Покровы (общие сведения). Как вытекает из названия, эти придаточные структуры служат «верхней одеждой» прокариотной клетки. Покров может образовывать:
— отдельная клетка;
— неупорядоченный (гроздь) или упорядоченный (цепочка или кубический пакет) клеточный агрегат, а также система из клеточных агрегатов;
— трихом или группа трихомов.
Материалом для покрова обычно служат полисахариды, реже это белки. В исключительных случаях покровы состоят из полипептидов (капсулу В. anthracis образует поли-D-глутаминовая кислота) или других полимеров (полифосфатная капсула Neisseria meningitidis), а также могут иметь липидную природу («ламинарный» слой гетероциты цианобактерий).
Покровы различаются по морфологии, толщине и консистенции. Их традиционно подразделяют на капсулы, слизи и чехлы, хотя далеко не всегда между ними удается провести четкую грань.
Капсула (лат. capsula — коробочка) представляет собой слизистый слой, непосредственно прилегающий к клеточной поверхности. Ее внешняя граница хорошо видна при фазово-контрастном микроскопировании. Классическим путем выявления капсулы служит прижизненное негативное контрастирование тушью, частицы которой в нее не проникают. На электронно-микроскопическом уровне порой выявляется слабая структурированность капсулы — тонкие фибриллы или слои, что может быть методическим артефактом. Гидрофильный материал капсулы непрочно связан с клеточной поверхностью и легко переходит в раствор.
Чехол (англ. sheath) виден под световым микроскопом безо всякого контрастирования. Его толщина может быть различной, хотя обычно она меньше, чем у капсулы. Чехол имеет плотную консистенцию. На электронно-микроскопическом уровне в нем иногда выявляются отдельные слои. Внешняя граница чехла резко очерчена, а между ним и клеточной оболочкой существует зазор. Хотя покров такого типа чаще всего состоит из гидрофильных полимеров, они слабо растворяются в воде.
Как правило, чехол обладает упорядоченной структурой, и на нем еще в прижизненном состоянии кристаллизуются минеральные соли, что происходит при образовании микробных матов (см. разделы 1.8 и 7.3.5). Поэтому если клетка покидает чехол или умирает в нем, он сохраняется еще очень долго.
Слизь (англ. slime) по составу идентична капсуле. Однако она не повторяет контуров клетки и может далеко распространяться за ее пределы. Таким образом, особенность слизи только в ее неопределенной морфологии и отсутствии тесной связи с клеточной оболочкой.
Поверхностные органеллы (общие сведения). Это множественные белковые структуры, имеющие округлую или продолговатую форму и в отдельных участках прикрепленные к оболочке клетки. Они сильно различаются по строению и функциям.
Конкретными примерами поверхностных органелл служат целлюлосомы (англ. cellulosome), шипы (лат. spina) и экстрацеллюлярные газовые баллоны (англ. extracellular gas bag).
8.5.4.1. Капсулы
Капсулы у прокариотов встречаются гораздо чаще, чем чехлы. Они широко и незакономерно представлены в домене Bacteria. Напротив, в домене Archaea они довольно редки — в качестве примера можно привести рода Methanosacrina и Staphylothermus. В большинстве случаев капсула состоит из экзополисахаридов.
Экзополисахаридные структуры (англ, exopolysaccharide structures, EPS) служат внешней границей между бактериальной клеткой и ее непосредственным окружением. Они очень важны в экстремальных условиях, поскольку защищают клетку от стресса и физико-химических повреждений.
Функции EPS. Их можно разбить на три основные группы — аккумулирующие, защитные и адгезионные.
Аккумулирующие функции состоят в концентрации неорганических ионов и низкомолекулярных органических субстратов вблизи клеточной поверхности.
Защитные функции состоят в предохранении клетки:
— от высыхания;
— от антибактериальных агентов (кислот, щелочей, катионов тяжелых металлов, поверхностноактивных веществ, антибиотиков, бактериоцинов, гидролитических ферментов и т. д.);
— от компонентов иммунной системы макроорганизма (комплемента, антител, макрофагов);
— от бактериофагов;
— от хищных бактерий и фаготрофных протистов.
Адгезионные функции состоят в том, что EPS служат одним из факторов клеточной адгезии, а также цементирующим веществом и обеспечивают:
— взаимную агрегацию бактерий данного вида и их агрегацию с другими членами микробиоты;
— прикрепление бактерий к субстрату.
В результате адгезии образуются гомологичные или гетерологичные ассоциации прокариотов и ядерных микроорганизмов — многоклеточные агрегаты, микроколонии, хлопья и биопленки.
Строение EPS. В состав EPS входят больше десяти мономеров, к которым относятся гексозы (галактоза, глюкоза, манноза), пентозы (арабиноза, ксилоза, рибоза), дезоксигексозы (рамноза, фукоза) и гексуроновые кислоты (глюкуроновая, маннуроновая), а также их производные, содержащие в качестве заместителей амино-, ацетильную, метильную и сульфатную группы, остаток пирувата или олигопептидную группировку. Благодаря присутствию уроновых мономеров и кислотных заместителей большинство EPS имеет анионную природу.
Молекула EPS представляет собой гомополисахарид или гетерополисахарид. В первом случае она состоит из одинаковых моносахаров. Во втором случае она содержит регулярно повторяющиеся субъединицы из моносахаров 2-5 типов, объединенных в блок по 2-8. Дадим наиболее интересные примеры.
Альгинаты. Название этих бактериальных EPS происходит от аналогичного продукта, извлекаемого из морских водорослей. Они представляют собой линейные сополимеры D-маннуроновой кислоты и ее эпимера L-гулуроновой кислоты, которые соединены β1 —> 4 связями. Участки поли-D-маннуроновой кислоты и участки поли-L-гулуроновой кислоты разделяются спейсерами из чередующихся мономеров и иногда содержат ацетильные заместители. Среди бактерий, образующих альгинаты, следует в первую очередь назвать Azotobacter vinelandii и Р. aeruginosa (класс «Gammaproteobacteria»).
Декстран. Название этого EPS связано с тем, что он состоит из правовращающей глюкозы, или декстрозы (лат. dexter — правый). Он представляет собой нейтральный разветвленный гомополимер D-глюкозы со связями α1 —> 2, α1 —> 3 и α1 —> 6. Его продуцентом, в частности, является Leuconostoc mesentheroid.es (фила ВХIII Firmicutes).
Ксантан. Название этого EPS происходит от родового имени продуцента, фитопатогенной бактерии Xanthomonas campestris (класс «Gammaproteobacteria»). Его основную цепь образует линейный β1 —> 4 полимер D-глюкозы. Боковой субъединицей является ацетилированный трисахарид из остатков маннозы и глюкозы с присоединенным остатком пирувата.
Леван. Название этого EPS связано с тем, что он состоит из левовращающей фруктозы, или левулозы (лат. laevus — левый). Это нейтральный разветвленный гомополисахарид, который состоит из остатков фруктозы, соединенных β1 —> 2 и β2 —> 6 гликозидными связями. Его, в частности, образует Gluconobacter oxydans (класс «Alphaproteobacteria») при росте на сахарозе.
Целлюлоза. Особое место среди нейтральных EPS занимает бактериальная целлюлоза. По современным данным, ее могут синтезировать некоторые штаммы из родов Aerobacter, Acetobacter, Achromobacter, Agrobacterium, Alcaligenes, Azotobacter, Pseudomonas, Sarcina и Rhizobium (фила BXII Proteobacteria), а также многие цианобактерии.
Биосинтез и экспорт EPS. Известны четыре основных механизма биосинтеза EPS.
В первом случае этот процесс происходит снаружи клетки и катализируется гликозилтранс- феразой. Характерным примером служит образование декстрана с помощью декстрансахаразы: n х сахароза —> (глюкоза)n + n х фруктоза.
Во втором случае полимер образуется в периплазматическом компартменте из нуклеотид-активированных мономеров. В качестве примера можно привести биосинтез целлюлозы. С биохимической точки зрения это очень простой процесс - УДФ-глюкоза транспортируется в периплазматический компартмент, где синтаза надстраивает линейную затравку: УДФ-глюкоза + (β1 —> 4- глюкоза)n —> УДФ + (β1 —> 4-глюкоза)n+1. Готовый полимер выделяется наружу через особые поры (см. раздел 8.4.3.1).
В третьем случае в цитоплазматическом компартменте синтезируется повторяющийся блок полимера, по аналогии с образованием предшественников ЛПС и муреина (см. разделы 8.4.1.1 и 8.4.3.1). Для его экспорта в периплазматический компартмент используется липидный переносчик — ундекапренилфосфат (см. рис. 35). Примером служит биосинтез ксантана или капсульных EPS. Как и в случае олигосахаридных субъединиц ЛПС, для переноса повторяющегося блока EPS в периплазматический компартмент используется Wzy-транслоказный комплекс (см. раздел 8.4.1.1). Сборка полимера осуществляется на наружной поверхности СМ, и он выделяется через контакты Байера (см. раздел 8.4.1.4).
В четвертом случае полисахарид синтезируется в цитоплазматическом компартменте, а затем секретируется через СМ и ОМ. Типичным примером служит капсульный К5-антиген Е. coli. В его биосинтезе и экспорте участвуют продукты генов kfiA-D (полимеризация) и kpsC, M, S, T (транслокация через СМ). Секреция происходит с помощью мультиполипептидного комплекса молекулярной массой >600 кДа, состоящего из >10 субъединиц (см. раздел 8.6.1). Проксимальный конец связал с СМ и содержит субъединицу, гомологичную ABC-транспортерам. Дистальный конец находится в ОМ и представлен секретином Wza, который имеет ß-спиральную бочковидную структуру и образует выходное отверстие канала диаметром ~8 нм.
Судьба EPS. После биосинтеза и экспорта (если таковой имел место) EPS могут сохранять связь с клеточной оболочкой и тогда они образуют материал капсулы. В результате гипертрофии капсулы (с потерей формы, повторяющей контур бактерии, и ослаблением связи с клеткой по мере удаления от ее поверхности) образуется слизь. Наконец, полный разрыв EPS с клеткой приводит к появлению в окружающей среде сбрасываемых полисахаридов (см. ниже).
Практическое значение EPS. Ценность EPS, особенно альгинатов и ксантана определяется их реологическими (греч. rheo — течь) свойствами, т. е. способностью повышать вязкость и пластичность различных материалов.
Они широко применяются как уплотнители (в текстильной промышленности), пластификаторы (для производства дорогих сортов бумаги), гелеобразователи (в кондитерской промышленности) и наполнители (в фармацевтике). Кроме того, они используются в строительстве (для улучшения качества дорожных покрытий), в нефтедобывающей отрасли (препятствуют примешиванию подземной воды к выкачиваемой нефти), а также в целях ремедиации (адсорбируют катионы тяжелых металлов и радионуклиды).
Не растворимые в воде декстран и целлюлоза применяются как молекулярные сита и защитные покрытия медицинского назначения, а в сухом виде — как мембраны для аудиотехники.
Капсулы непатогенных бактерий. В одних случаях капсулы ненамного толще клеточной оболочки; в других они довольно массивны (больше диаметра клетки).
Как уже указывалось, структурной основой капсулы служат линейные или разветвленные полигликаны и полипептиды, состоящие из одинаковых или разных мономеров. Гибкие фибриллы капсульных полимеров ориентируются перпендикулярно поверхности клетки и сильно гидратированы — до 99% капсулы составляет вода.
Обводненность капсулы делает проблематичным сохранение этой структуры на электронно-микроскопических препаратах. Если полимер капсулы имеет кислотную природу (например, за счет карбоксильных групп), его цепи взаимодействуют друг с другом через электростатические мостики с помощью катионов Са2+ или Mg2+. Этим свойством широко пользуются морфологи, вводя в состав фиксаторов катионные красители, образующие межмолекулярные сшивки (например, рутений красный и альциан синий), а также диамины, лектины и антикапсульные антитела.
Основной функцией капсулы у непатогенных бактерий служит предохранение клетки от водного стресса на засушливых территориях или в водоемах с повышенной соленостью. В частности, у цианобактерий рода Nostoc устойчивость к высушиванию определяется капсулой, которая создает водный «буфер» между клетками и атмосферой и позволяет им расти в пустынях в виде корочек, смачиваемых только утренней росой.
Образование капсул у непатогенных бактерий индуцируется в стрессовых условиях, в частности при фосфорном голодании.
Сбрасываемые полисахариды непатогенных бактерий. Известно много случаев, когда обводненные капсульные полимеры теряют связь с клеточной оболочкой, в особенности, когда происходит их гиперпродукция. Тогда они выделяются в окружающую среду и начинают хаотически плавать в ней до тех пор, пока они не вступят в контакт с каким-нибудь (а)биотическим материалом.
Такие обводненные капсульные полимеры называются «внешней» слизью (анл. external slime), или «сбрасываемыми» полисахаридами (англ. released polysaccharides, RPS).
Капсулы патогенных бактерий. У зоопатогенных бактерий, особенно патогенов высших животных, капсула имеет важное значение для вирулентности и патогенности, поскольку они сталкиваются с иммунной системой в органах и тканях, которые в норме стерильны.
Капсулы оказывают комплексное защитное действие:
— они позволяют бактерии уклониться от фагоцитирования, поскольку EPS придают ее поверхности гидрофильность, упругость и электрический заряд;
— они служат барьером для проникновения к поверхности бактерии компонентов комплемента и бактерицидных белков, например, дефензинов;
— они препятствуют взаимодействию бактерии с антителами (бактерии не агглютинируются О-антисывороткой, поскольку «соматический» О-антиген маскируется «капсульным» К-антигеном (от нем. Kapsel — капсула);
— они сами не активируют комплемент и маскируют активирующие его компоненты клеточной оболочки, что препятствует неспецифической опсонизации бактерий полиморфноядерными лейкоцитами;
— они слабоиммуногенны, что препятствует специфической опсонизации (т. е. связыванию компонентов комплемента и антител с поверхностью бактерии, которое облегчает их поглощение макрофагами); тем не менее, они используются в качестве вакцин, и к ним вырабатываются специфические антитела;
— они избегают иммунного ответа, поскольку в отдельных случаях мимикрируют под молекулярные компоненты хозяина; примерами служат капсульный Kl- антиген Е. coli, который содержит сиаловую кислоту и мимикрирует под компонент гликокаликса в разных типах хозяйских клеток. Капсульный К5-антиген этой же бактерии представляет собой гетерополимер глюкозамина и N-ацетилглюкозамина, и мимикрирует под гепарин в составе аморфного вещества хряща.
(Липо)полисахаридная капсула энтеробактерий. Капсульные EPS имеются как у грамположительных (род Streptococcus), так и у грамотрицательных (Е. coli; рода Haemophilus, Klebsiella, Neisseria и др.) бактерий. Лучше всего они изучены на примере Е, coli, у которой выявлено >80 типов К-антигена.
При классификации капсульных полисахаридов используются такие критерии, как природа кислотного компонента, величина электрического заряда, наличие липидного домена на редуцирующем конце, температурный контроль биосинтеза и совместная экспрессия с ЛПС.
Различают две основные группы капсульных полисахаридов.
К I группе относятся менее крупные (<50 кДа) полисахариды, состоящие из остатков гексуроновых кислот. Фактически это специализированные липополисахариды, поскольку на редуцирующем конце они содержат липид А (см. раздел 8.4.1.1). Они встречаются у относительно немногих О-серотипов, синтезируются при температуре <30°С и не играют роли в патогенезе, а лишь способствуют выживанию бактерии вне хозяина. Примером такого полисахарида служат колановые кислоты (англ. colanic acid), которые образует Е. coli.
Ко II группе относятся более крупные (>50 кДа) полисахариды, состоящие из остатков гексуроновых кислот, сиаловой кислоты и КДО. Фактически это тоже спе- циализированые липополисахариды, поскольку на редуцирующем конце содержат фосфатидную кислоту. Они имеются у большинства О-серотипов и не образуются при температуре <20°С. Они важны для патогенеза, и их примерами являются К1- и К5-антигены Е. coli.
Гены для биосинтеза К-антигенов II группы расположены на хромосоме в районе локусов his и trp, а гены для их экспорта —в районе локуса serA. Соответствующий участок хромосомы подразделяется на центральный район 2 (kf i-гены), отвечающий за полимеризацию, и фланкирующие его районы 1 и 3 (fcps-гены), отвечающие за экспорт.
Гены для биосинтеза и экспорта К-антигенов I группы еще слабо изучены.
Полипептидная капсула Bacillus anthracis. Патогенная бактерия В. antrhracis вызывает сибирскую язву у рогатого скота и смертельно опасна для человека. В данном случае важным фактором вирулентности является поли-D-глутаматная капсула. За ее синтез отвечают плазмидные гены cap АВС. Капсула несет высокий отрицательный заряд, что не позволяет макрофагам фагоцитировать вегетативные клетки бактерий. Если эндоспора прорастает в присутствии СО2, то вокруг нее скапливается капсульный материал, обволакивающий вегетативную клетку при ее выходе из спорангия. Белок Dep (его ген идет следом за кластером cap АВС) кодирует деполимеразу, которая расщепляет поли-D-глутамат на фрагменты, блокирующие иммунную систему макроорганизма.
Полифосфатная капсула Neisseria meningitidis. Одним из основных компонентов капсулы N. meningitidis является конденсированный неорганический фосфат, который составляет ~50% запасов этого вещества (см. раздел 8.3.11.4). Возможно, что, благодаря своим, сильно выраженным полианионным свойствам капсула позволяет N. meningitidis избежать фагоцитирования макрофагами точно так же, как это удается В. anthracis. Во всяком случае, при мутации по гену полифосфаткиназы ррк опасный патоген теряет устойчивость к сывороточным белкам, и эту мутацию можно комплементировать при помощи плазмиды, несущей ген дикого типа.
Капсулы бактерий, ассоциированных с растениями. В данном случае состав и функции капсул могут существенно различаться.
У фитопатогенных бактерий капсула специальной роли не выполняет. Например, у возбудителей вилта и некроза (см. ниже) выживание в окружающей среде обеспечивается за счет таких свойств капсулы, как гигроскопичность, вязкость и электростатический заряд. При попадании внутрь растения она облегчает распространение инфекционного агента по тканям, защищает бактерии при размножении в межклетниках и предохраняет их от иммунных систем хозяина.
У симбионтов роль капсулы высокоспецифична и связана с сигналингом. Например, у клубеньковых бактерий капсула участвует в прикреплении к корневому волоску и запуске регуляторного каскада, что приводит к внедрению бактерий в ткани и подавлению иммунных систем хозяина.
Примером ЕРS у фитопатогенных бактерий является амиловоран. Его образует Erwinia amylovora — возбудитель вилта, или увядания томатов и других ценных сельскохозяйственных культур. В состав этого крупного гетерополимера (50-150 мДа) входят остатки галактозы, глюкуроновой кислоты и пирувата. В качестве других примеров можно назвать альгинаты Р. syringae, ксантан X. campestris и сложный набор гетерополисахаридов Р. solanacearum. Перечисленные бактерии вызывают у растений разнообразные некрозы.
Примером ЕРS у симбионтов является сукциногликан, который образуется многими штаммами рода Rhizobium. Он сильно варьирует по составу и представляет собой линейную цепь (в которой остатки глюкозы, глюкуроновой кислоты и галактозы соединяются связями β1—> 4) с боковыми цепями, образоваными 1-4 остатками глюкозы и галактозы. В главной и боковой цепях встречаются заместители — остатки ацетата, гидроксибутирата, пирувата и сукцината.
8.5.4.2. Чехлы
По широте распространения чехлы значительно уступают капсулам. Их могут образовывать как вегетативные клетки, так и разные варианты дифференцированных клеток (цисты, эндоспоры, гетероцисты и «везикулы» актинобактерий). В данном случае мы будем говорить только о чехлах вегетативных клеток. Чехлы покоящихся клеток (цист и эндоспор), гликолипидные чехлы метаболически специализированных клеток цианобактерий (гетероцист), а также гопаноидные чехлы «везикул» актинобактерий мы рассмотрим в III томе учебника.
Чехлы у хламидобактерий. Хламидобактериями называют членов кластера «Leptothrix-Sphaerotilus» (класс «Betaproteobacteria»), образующих трубчатый чехол. Обычно в нем помещается цепочка клеток, реже — продольный тяж из 2-3 цепочек. В ходе жизненного цикла клетки приобретают полярный жгутик, с помощью которого плавают, находят новую нишу и оседают на твердый субстрат. Для адгезии на полюсе, противоположном жгутику, выделяется клейкое вещество. Оно затвердевает, образуя «прикрепительный диск» (англ, holdfast). В этом месте закладывается чехол, обволакивающий цепочку делящихся клеток. Так происходит у L. lopholea и S. natans\ остальные хламидобактерии специального прикрепительного диска не образуют и вступают в адгезию с твердым субстратом при помощи полисахаридного чехла, содержащего оксиды и гидроксиды железа и марганца. Катионы Fe2+связываются кислыми полисахаридами, а затем окисляются абиогенно или ферментативным путем. Катионы Мn2+ вначале также связываются с чехлом, но их окисление происходит только при помощи ферментов.
Чехол хламидобактерий состоит из одного или нескольких слоев. Внутренний слой чехла L. discophora представляет собой матрицу из белковых фибрилл диаметром 6,5 нм, которые стабилизированы сульфгидрильными связями и дисульфидными мостиками. Чехол образует анастомозы с ОМ и утолщается в этих участках. Над
внутренним фибриллярным слоем расположен диффузный слой, образованный отрицательно заряженными полисахаридами, в состав которых входят уроновые кислоты и глюкозамин. Диффузный слой чехла S. natans содержит галактозу, глюкозу и фукозу, а также глюкуроновую кислоту.
Важнейшими функциями чехла у хламидобактерий являются:
— адгезионная, поскольку с его помощью происходит прикрепление к твердым поверхностям, что позволяет адаптивно заселять мелкие водные артерии;
— защитная, поскольку он непроницаем для фаготрофных протистов и хищных бактерий (например, Bdellovibrio bacteriovorus).
Чехол у хламидобактерий участвует в процессах биоминерализации, что приводит к образованию залежей железных и марганцевых руд.
Чехлы у цианобактерий. Чехлы образуются как у одноклеточных, так и трихомных цианобактерий и обычно имеют хорошо выраженную фибриллярную или паракристаллическую структуру.
У одноклеточной диазотрофной цианобактерии Gloeothece sp. растяжимый чехол подразделяется на внутренний и внешний. Первый плотно прилегает к ОМ одиночной клетки, а второй окружает группу из 4-16 клеток. Основу обоих чехлов составляет гетерополисахарид, состоящий из нейтральных сахаров — галактозы, глюкозы, маннозы, O-метилксилозы и ксилозы, а также из галактуроновой, глюкуроновой и маннуроновой кислот. Наружный чехол менее гидратирован и слабее сульфатирован, чем внутренний. Предполагается, что чехол Gloeothecesp. выполняет барьерную функцию и защищает нитрогеназу от экзогенного кислорода.
Особую роль играет чехол трихомной цианобактерии Scytonema sp. Он содержит сцитонемин - продукт конденсации фенольных и индольных (бензопиррольных) групп. Это индуцибельное соединение сильно поглощает «ближнее» (320-400 нм, UV-А), «среднее» (280-320 нм, UV-В) и «дальнее» (190-280 нм, UV-С) ультрафиолетовое излучение. Оно синтезируется на ярком солнечном свету и на 80-90% защищает клетки от облучения. Сцитонемин выявлен у многих одноклеточных и трихомных морских, пресноводных и почвенных штаммов разного географического происхождения. Скорее всего, это молекулярный реликт, который в Докембрии способствовал широкому расселению цианобактерий в отсутствии атмосферного озонового слоя.
Функцию ультрафиолетового экрана может выполнять и микрофибриллярная целлюлоза. Она обнаружена на периферии очень толстого, до 1 мкм, чехла у некоторых штаммов одноклеточных (Gloeocapsa sp.) и трихомных (Anabaena sp., Phomnidium sp., Nostoc sp. и Scytonema sp.) цианобактерий. По-видимому, этот биополимер секретируется через комплексы контактных пор (см. раздел 8.4.3.1).
Чехлы у архей. В отличие от бактерий, у архей чехлы бывают крайне редко, и они имеют особое строение.
Первый пример связан с метаногенами Methanosaeta thermophila и Methanospirillum hungatei, которые растут в виде неразветвленной цепочки из палочковидных клеток. Полуригидная стенка представлена S-слоем из негликозилированных белковых протомеров молекулярной массой ~100 кДа. Между соседними клетками находятся дискоидные белковые спейсеры (англ. spacer plug). Все это одето открытым на концах цилиндрическим чехлом, внешний диаметр которого составляет ~1 мкм, а длина ~150 мкм. Чехол образован системой колец или туго скрученной спиралью из димерных белковых субъединиц молекулярной массой около 50 кДа. Он исключительно устойчив к химическим реагентам, а также гидролитическим ферментам.
Стенка у таких архей довольно гибкая, и форму клетки определяет не она, а экзоскелет — чехол и фланкирующие спейсеры. Деление осуществляется уникальным способом. Материнская клетка перетягивается наружным «кольцом деления» (англ. division annulus), которое закладывается в промежутке между S-слоем и чехлом и закрывается, как ирисовая диафрагма, превращаясь в спейсер. Кольцо деления имеет вид спиральной ленты и состоит из белка.
Второй пример связан с представителями рода Methanosarcina, клеточную стенку которых образует ригидный S-слой. Каждая клетка окружена толстым (до 200 нм) слоистым чехлом, состоящим из гетерополимера «метанохондроитина». Клетки после деления не расходятся, а образуют округлые пакеты, окруженные общим фибриллярным чехлом. Подобно внутреннему чехлу, он содержит метано- хондроитин.
Строительный блок метанохондроитина состоит из двух остатков N-ацетилгалак- тозамина и остатка глюкуроновой или галактуроновой кислоты, которые соединены связями β1 —> 3 и β1 —> 4. Метанохондроитин синтезируется из активированных предшественников: УДФ-N-ацетилгалактозамина и УДФ-глюкуроновой кислоты. После образования УДФ-активированного трисахарида он переносится на ундекапренил- фосфат (см. рис. 35), транслоцируется в экзоплазматический компартмент и там присоединяется к затравке полимера.
Интересно, что по строению, а также способу биосинтеза метанохондроитин напоминает стеночную тейхуроновую кислоту (см. раздел 8.5.2.1). И в то же время он сходен с хондроитинсульфатом — одним из компонентов аморфного матрикса соединительной ткани, хотя отличается от него тем, что не сульфатирован и содержит не одну, а две молекулы N-ацетилгалактозамина на молекулу глюкуроновой кислоты.
8.5.4.3. Шипы и экстрацеллюлярные газовые баллоны
К группе поверхностных органелл относятся два типа специализированных компартментов — шипы и экстрацеллюлярные газовые баллоны.
Шипы. В отечественной литературе для обозначения этих поверхностных органелл также используется термин «спин» (лат. spina — шип). Они встречаются у грамотрицательных бактерий (в частности, у псевдомонад, метилотрофов и цианобактерий), но совершенно неизвестны у архей.
Шипы представляют собой жесткие полые конические или конусообразно-цилиндрические сруктуры. Среднее число шипов на клетку равно десяти, своим основанием они прикреплены к ОМ, ориентированы перпендикулярно клеточной поверхности и расположены нерегулярно. Шипы варьируют по размеру (диаметр 50-100 нм, длина 0,5-3 мкм). Стенка шипа имеет толщину ~7 нм и образована 2,5-105 спирально уложенными субъединицами белка спинина (~42 кДа), молекула которого состоит из двух антипараллельных (3-цепей.
Шипы образуются путем самосборки с дистального конца. Изнутри они полые; конусообразно-цилиндрические шипы имеют открытый конец, а конические шипы запечатаны.
В лабораторных условиях образование шипов индуцируется при повышении температуры и pH или при снижении ионной силы среды. Физиологическая роль шипов в природе неясна, особенно если учесть, что на их построение уходит очень много белка. Скорее всего, они мультифункциональны и обеспечивают повышенную пла-
вучесть, образование микроколоний, прикрепление к (а)биотическим субстратам и защиту от фаготрофных протистов.
Экстрацеллюлярные газовые баллоны. Этот редко встречающийся тип специализированных компартментов впервые описал в 1985 г. В. И. Дуда на примере бактерии Bacillusvesiculiferous, живущей при дефиците кислорода в кишечнике «сигнального» рачка Pacifastacus leniunsculus. Экстрацеллюлярные газовые баллоны представляют собой крупные газонакопительные структуры, прикрепленные к наружной поверхности клетки. Они имеют вид сферических, цилиндрических или игольчатых везикул величиной от нескольких десятков нанометров до нескольких десятков микрометров. Стенка экстрацеллюлярного газового баллона образована неунитарной мембраной толщиной ~2 нм.
Функцией экстрацеллюлярных газовых баллонов, по-видимому, служит запасание О2, который необходим бактерии для энергетического и конструктивного метаболизма. В этом плане они аналогичны кислородным вакуолям Beggiatoa-подобных бактерий (см. раздел 8.3.11.1), но отличаются от них локализацией и природой мембранной границы.
Механизм образования и эволюционное происхождение экстрацеллюлярных газовых баллонов неизвестны. Ясно, что это не блебинг-везикулы S-слоя, поскольку сквозь его поры легко проходят гидрофильные молекулы. Заманчиво предположить, что мембрана экстрацеллюлярных газовых баллонов состоит из белка, аналогичного или гомологичного полипетиду газовой везикулы (см. раздел 8.3.11.2). В связи с этим встал бы вопрос о его секреции и доставке на наружную поверхность клетки (см. раздел 8.6.1).
8.5.4.4. Целлюлосомы
Целлюлоза, главный структурный компонент клеточной стенки растений, ежегодно синтезируется в количестве ~4 • 1011 т. Не растворимая в воде целлюлоза является одним из наиболее рекальцитрантных обновляемых субстратов. Тем не менее, в природе она постоянно разрушается с помощью грибных и бактериальных ферментов, что является очень ответственным звеном в глобальном цикле углерода. Кроме того, целлюлолитические бактерии служат перспективными поставщиками гексоз и пентоз, используемых в качестве стартового сырья при бродильном производстве этанола.
В природе целлюлоза разрушается как в аэробных, так и в анаэробных условиях.
Аэробный процесс, приводящий к полному окислению целлюлозы до СО2 и Н2О, может осуществляться отдельным видом грибов, например, белой плесенью Phanerochaetechrysosporium или отдельным видом бактерий, например, Cellulomonas cellulosolvens и Thermomonospora curvata (фила BXIV Actinobacteria), а также Cellvibrio vulgaris (класс «Gammaproteobacteria»).
Анаэробный процесс заканчивается образованием СО2 и СН4. Его осуществляет уже не отдельный вид, а целое микробное сообщество:
— бактерии, гидролизующие целлюлозу до целлодекстринов и глюкозы;
— бактерии, сбраживающие глюкозу до Н2/СО2, спиртов и органических кислот;
— автотрофные и гетеротрофные бактерии, синтезирующие из этих субстратов ацетат;
— автотрофные археи, образующие СН4 из СО2/Н2;
— гетеротрофные археи, образующие СН4 из ацетата.
Молекула целлюлозы состоит из 5 • 102 — 1,5 • 104 остатков глюкозы (протомеров), связанных β1 —> 4 гликозидными связями. Поскольку каждый протомер развернут по отношению к своему соседу на 180°, повторяющейся единицей молекулы целлюлозы является дисахарид целлобиоза. С помощью водородных связей и сил Ван дер Ваальса, действующих между пиранозными кольцами, молекулы целлюлозы ориентируются параллельно друг другу, образуя не растворимые в воде кристаллические микрофибриллы, которые содержат 10-40% аморфных участков. Микрофибриллы целлюлозы объединяются в макроскопические образования — волокна.
Целлюлоза редко встречается в чистом виде (например, во вторичной стенке волосков на семенах хлопка). Обычно ее фибриллы погружены в аморфный матрикс, состоящий из гемицеллюлоз, лигнина и пектинов, что делает этот полимер малодоступным для гидролитических ферментов.
Гемицеллюлозы (ксиланы и маннаны) по валовому накоплению являются вторым в природе полимером после целлюлозы. Линейная цепь ксиланов образована остатками ксилозы. Боковыми заместителями являются арабиноза, уксусная кислота и (метил)глюкуроновая кислота, а также ароматические кислоты — р-кумаровая и феруловая. Лигнин состоит из модифицированных остатков ароматических спиртов («монолигнолов») — кониферилового, р-кумарилового и синапилового. Он связан с боковыми цепями гемицелюллоз через остатки р-кумаровой и феруловой кислот. Пектины являются линейными гомополимерами галактуроновой кислоты, карбоксильные группы которой этерифицированы метанолом.
Гидролиз целлюлозы служит примером гетерофазной реакции и аналогичен гидролизу липидов с помощью липаз. Он осуществляется набором внеклеточных целлюлаз, атакующих внутренние гликозидные связи цепи целлюлозы с образованием целлодекстринов (эндоглюканазы). Кроме того, целлюлазы отщепляют концевой дисахарид целлобиозу (экзоглюканазы, или целлобиогидролазы), а также гидролизуют низкомолекулярные целлодекстрины и целлобиозу до мономерной глюкозы (β-глюкозидазы).
Гемицеллюлозы разрушаются грибными и бактериальными ксиланазами и манназами, гидролизующими линейные цепи, а также вспомогательными ферментами (арабинофуранозидазами, ацетилксилан-эстеразами, глюкуронидазами и ферулатэстеразами), которые атакуют боковые цепи. Рекальцитрантный лигнин разрушается с помощью грибного лигнолитического комплекса, состоящего из лакказ (медьсодержащих оксидаз) и пероксидаз.
Пектиназы (деполимеразы и эстеразы) действуют на линейную цепь полимера, а также отщепляют от нее боковые метильные группы.
Бактерии, разрушающие целлюлозу одновременно с гемицеллюлозами и пектинами, синтезируют комплекс соответствующих гидролитических ферментов. Продукты распада целлюлозы — целлодекстрины, целлобиоза и глюкоза — используются в качестве субстратов для роста аэробных и анаэробных микроорганизмов. Основная часть этих субстратов (90-95%) разлагается в аэробных условиях с образованием Н2О и окисленного конечного продукта, СО2. Остальные 5-10% разлагаются в анаэробных условиях с образованием Н2О и восстановленного конечного продукта, СН4. В обоих случаях происходит минерализация глюкозы — процесс, который является важным звеном в глобальном круговороте углерода.
Для ферментативного разрушения целлюлозы микроорганизмы используют три стратегии:
— выделение в окружающую среду набора целлюлаз;
— выделение в окружающую среду одного фермента с комплексной целлюлолитической активностью;
— компартментализация целлюлаз в целлюлосоме, расположенной в экзоплазматическом компартменте.
Целлюлосомы были впервые описаны в 1983 г. израильскими биохимиками Ламедом (R. Lamed) и Байером (Е. А. Bayer) у анаэробной термофильной бактерии Clostridium thermocellum. В настоящее время эти надмолекулярные комплексы обнаружены только у бактерий из филы ВХIII Firmicutes (Acetivibrio cellolyticus, Butyrivibrio fibrisolvens, Clostridium acetobutylicum, C. cellobioparum, C. cellulolyticum, C. cellulovorans, C. josui, C. papyrosolvens, C. thermocellum и Ruminococcus albus), а также у бактерий из филы ВХХ Bacteroidetes(Bacteroides cellulosolvens). Нишами для этих бактерий служат почва, компостные кучи, сточные воды деревообрабатывающих и бумажных фабрик, а также рубец жвачных животных.
Роль целлюлосомы при биодеградации целлюлозы заключается в том, что этот
микрокомпартмент обеспечивает адгезию бактерий к субстрату, оптимальное взаимодействие целлюлаз и канализированное поступление продуктов гидролиза к поверхности клетки.
Целлюлосома — самый крупный из внеклеточных ферментных комплексов. В изолированном виде она представляет собой сферическую частицу диаметром ~20 нм. На поверхности клетки целлюлосомы образуют протуберанцы длиной 50- 100 нм.
Целлюлосома имеет сложный полипептидный состав (14-26 компонентов молекулярной массой 20-200 кДа) и очень большую молекулярную массу (2 • 103 - 6,5 • 103 кДа). Это исключительно стабильная структура, не диссоциирующая в растворе с высокой ионной силой и под действием детергентов.
После первичного контакта с фибриллами целлюлозы целлюлосомы агрегируют в полицеллюлосомы, которые служат коридорами для диффузии растворимых продуктов гидролиза. Полицеллюлосома имеет молекулярную массу 5 • 104 -105 кДа и состоит из тех же самых субъединиц, что и целлюлосома.
Целлюлазные субъединицы одномоментно производят множественные разрезы внутри молекулы субстрата. Целлюлосома гидролизует как аморфную, так и кристаллическую целлюлозу в основном до целлобиозы, т. е. она проявляет и эндоглю- каназную, и экзоглюканазную активность.
В некоторых случаях (примером служит С. cellulolyticum) целлюлосомы вместе с целлюлазами содержат гемицеллюлазы и пектиназы. Для таких гетероферментных компартментов предложено название «целлволдазы» (от англ. cell wall — клеточная стенка). Кроме того, в состав целлюлосом могут дополнительно входить хитиназы, что позволяет использовать хитин (структурный аналог целлюлозы, в котором гидроксильные группы в положении 2 замещены аминогруппами). Структурную основу целлюлосомы составляет большой гликопротеин CipC (от англ. cellulosome integrating protein; 210-250 кДа). У него есть второе название — «скефолдин» (англ. scaffold — каркас). Наиболее изученный скефолдин С. thermocellum содержит функционально специализированные домены, разделенные спейсерными участками (рис. 58). Известны три основных типа таких доменов.
Рис. 58. Целлюлосома. СМ — цитоплазматическая мембрана; М — муреиновый саккулус; S — поверхностный белковый слой; СiрС— скефолдин; SdbА — скефолдинсвязывающий белок.
Во-первых, это восемь копий гидрофобного «когезина» Cohi (англ. cohesion — сцепление). Отдельный когезин состоит из 138 аминокислотных остатков и служит для связывания целлюлазной субъединицы через «докерины» (англ. dock — состыковывать) Doci. Для этого у нее существуют продублированные последовательности размером в 22 аминокислоты. Когезины разных бактерий взаимно гомологичны, и их число равно 5-9.
Во-вторых, это «докерин» Doch, фиксирующий целлюлосому на теле клетки. Его партнером является «когезин» CohII — дистальный домен белка SdbA (от англ. scaffoldin dockerin binding). Благодаря тому, что на С-конце белка SdbA содержатся домены, гомологичные доменам S-слоя (англ. S-layer homology domains, SLH), он интегрирован в клеточную стенку. Помимо белка SdbA на поверхности бактерии расположены белки 01рА/В. У них имеются собственные когезины обоих типов, и они также участвуют в связывании скефолдина и целлюлаз.
Втретьих, это одиночный домен СВМ (от англ. carbohydrate binding module). С его помощью целлюлосома прикрепляется к фибриллам целлюлозы преимущественно в кристаллических участках. В некоторых случаях, например, у мезофильной целлюлолитической бактерии R. albus, для прикрепления к целлюлозе дополнительно используются адгезионные фимбрии (см. раздел 8.6.3), состоящие из пилина СbрС (от англ. cellulose binding protein).
Гены, кодирующие компоненты целлюлосомы, образуют кластер размером ~26 т. п. н., либо рассеяны по всему геному.
8.5.4.5. Экзоплазматические микрокристаллы
У некоторых цианобактерий в экзоплазматическом компартменте образуются микрокристаллы минералов, в частности кальцита СаСОз. Матрицей для начала кристаллизации служат узлы паракристаллической решетки S-слоя (см. раздел 8.5.2.3), либо регулярно расположенные полисахаридные или белковые фибриллы чехла (см. раздел 8.5.4.2). Прижизненная или постмортальная минерализация способствует формированию микробных матов, на основе которых возникают ископаемые строматолиты (см. разделы 1.8 и 7.3.5).
Опосредованная цианобактериями кристаллизация минералов существенно влияет на глобальные циклы биогенных элементов. В морских микробных матах на трихомах цианобактерий повсеместно происходит отложение гипса СаSO4 • 2Н2O, фосфата кальция Са3(РO4)2 и галита NaСI. Кальцифицированные трихомы образуют и аэрофитные (растущие на увлажняемых наземных субстратах, а не в водной толще) цианобактерии, например, штаммы Scytonema sp.
Формирование экзоплазматических микрокристаллов не сводится к простому физико-химическому процессу, а зависит от состава и структуры чехла, а также от способности цианобактерий повышать pH экзоплазматического компартмента в результате ассимиляции «растворенного неорганического углерода»
(СO2/НСO3-/СO32-).
8.6. Специализированные компартменты со смешанной локализацией
В этом разделе мы рассмотрим микрокомпартменты, расположенные не в каком- нибудь одном, а сразу в двух или даже трех генеральных компартментах.
Характеризуя ОМ и муреиновый саккулус, мы уже останавливались на компартментах этого типа. Речь шла о контактах Байера (см. раздел 8.4.1.4), а также о микроплазмодесмах и «комплексах контактной поры» (см. раздел 8.4.3.1).
Теперь же мы обратимся к примерам тех компартментов со смешанной локализацией, которые выполняют широкий спектр экспортных функций. Они представляют собой раскрывающиеся каналы в клеточной оболочке, через которые экспортируются белки (исключением служит импорт чужеродной ДНК, обеспечивающий генетическую трансформацию).
У грамположительных бактерий и архей белок транслоцируется только через одну мембрану — СМ. У грамотрицательных бактерий при экспорте белка возможны две ситуации. Если таргетинг предусматривает взаимодействие белка с наружной поверхностью СМ или секрецию в периплазматическое пространство, экспорт происходит без участия ОМ. Однако, если таргетинг предусматривает ассоциацию с внешней поверхностью клетки или секрецию во внешнюю среду, белок необходимо дополнительно транслоцировать через ОМ.
Во всех случаях для прохождения белка через клеточную оболочку требуется канал (секреторная пора). В связи с этим перед нами встают три взаимосвязанных вопроса:
— на основе каких молекулярных структур формируется секреторная пора;
— какая из форм энергии, ассимилированной клеткой, используется для работы по транслокации белка;
— в каком конформационном состоянии белок проходит через секреторную пору.
При ответе на эти вопросы мы будем говорить не только о системах секреции
белка, но и о системах подвижности прокариотной клетки. Казалось бы, между ними не должно быть ничего общего. Однако теперь стало известно, что системы секреции белка и жгутиковый аппарат содержат взаимно гомологичные субъединицы, а это указывает на их общее эволюционное происхождение.
8.6.1. Системы секреции белка
Белки синтезируются на цитоплазматических рибосомах и в ходе секреции транслоцируются через СМ.
Белки, которые транслоцируются через СМ, как правило, содержат N-концевой сигнальный пептид, или сигнальную последовательность. Она распознается специфическими рецепторами экспортного комплекса. В ходе транслокации белка или при попадании на наружную поверхность СМ сигнальный пептид удаляется сигнальной пептидазой (англ. signalpeptidase, Sep). У бактерий, архей и эукариотов сигнальные пептиды взаимно гомологичны. Они состоят из 20-30 аминокислотных остатков и содержат три домена — N-концевой гидрофильный домен, центральный гидрофобный домен и С-концевой домен, заканчивающийся сайтом для расщепления сигнальной пептидазой.
У бактерий белки экспортируются в основном посттрансляционно, т. е. после завершения биосинтеза. Предназначенный для транслокации белок может иметь альтернативную конформацию — свернутую или развернутую.
В первом случае белок приобретает третичную структуру еще в цитоплазме (хотя на внешней стороне СМ от него отрезается сигнальный пептид). Ясно, что для экспорта такого белка необходима крупная пора.
Во втором случае белок поддерживается в развернутой конформации молекулярным шапероном (см. раздел 8.3.10.2), «продевается» N-концом вперед через транспортер и только на выходе приобретает свернутую конформацию.
У прокариотов известен ряд систем для секреции белка. В соответствии со значением для жизнедеятельности и широтой распространения, эти системы можно разбить на две основные группы — генеральные и специализированные. Конкретные варианты специализированных систем обозначают цифрами I-IV.
Генеральных систем две — Tat-система и Sec-система. Первая из них секретирует белок в свернутой конформации, вторая в развернутой.
Генеральная секреторная Tat-система. У белков, которые транспортируются с помощью этой системы, сигнальный пептид содержит между N-концевым доменом и гидрофобным доменом консервативный мотив с двумя соседними остатками Arg. Отсюда ее название, Tat-система (от англю twin-arginine translocation, «транслокация спаренных аргининовых остатков»).
Генеральная Tat-система (рис. 59) была впервые обнаружена в хлоропластах. К настоящему времени известно, что она имеется у многих представителей фил ВХ Cyanobacteria, BXIIProteobacteria, ВХIII Firmicutes и BXIV Actinobacteria, а также у ряда представителей фил AI Crenarchaeota и АII Euryarchaeota. Наряду с этим есть филы, где она вообще не встречается (например, фила BXVI Chlamydiae).
Особенность генеральной Tat-системы заключается в том, что она:
— секретирует белки в свернутой конформации;
— сигнальный пептид содержит консервативный мотив Ser(Thr)-Arg—Arg-X- Phe-Leu-Lys;
— белок транслоцируется за счет протондвижущей силы (Pmf).
Сначала считали, что субстратами для Tat-системы служат только субъединицы электрон-транспортных цепей, связанные с наружной поверхностью СМ и содержащие редокс-активные центры (белки Риске, гидрогеназы, нитритредуктаза, фор- миатдегидрогеназа). Кофакторы этих белков — ФАД, НАДФ, гемы, железосерные центры, ионы металлов переходной валентности, молибдоптерины и т. д. (см. II том учебника) очень лабильны и связываются с апопротеинами только в цитоплазме. Для этого апопротеины должны предварительно приобрести третичную структуру, а в некоторых случаях из нескольких субьединиц должен образоваться мультимерный комплекс.
Теперь установлено, что функции Tat-системы в секреции белка этим не ограничиваются. Например, с ее помощью могут экспортироваться фосфолипаза С, щелочная фосфатаза PhoD, N-ацетилмурамил-L-аланин-амидаза, а также факторы вирулентности некоторых патогенных бактерий, например, Р. aeruginosa.
Генеральная Tat-система образуется конститутивно. Ее кодируют tat-оперон, в состав которого входят гены tatABCD, и моноцистронный оперон tatE. Взаимно гомологичные белки TatA, TatB и TatE состоят из двух α-спиральных доменов — трансмембранного и амфипатического; интегральный белок TatC содержит шесть α-спиральных доменов. Все субъединицы Tat-системы представлены несколькими копиями.
Как именно работает Tat-система, точно еще неизвестно. Возможно, что субъединицы TatB и TatC служат ее каркасом, а также рецептором, тогда как субъединицы TatA образуют раскрывающуюся гидрофильную пору диаметром до 70А, через которую и проходит секретируемый белок. Необходимо еще раз подчеркнуть,
что источником энергии для Tat-системы служит Pmf а не гидролиз нуклеозидтри- фосфатов (АТФ и/или ГТФ).
Некоторые бактерии секретируют с помощью Tat-системы до 20% белков, а ар- хеот Halo bacterium sp. NRC-1 (фила All Euryarchaeota) — большинство белков.
Поскольку археи синтезируют уникальные мембранные липиды (см. раздел 8.3.3.2) и часто занимают экстремальные ниши (см. главы 3 и 7), неудивительно, что их секреторные системы во многом отличаются от бактериальных. Например, субъединица TatC2 у Halobacterium sp. NRC-1 имеет аномальное строение (состоит из 14 доменов и, видимо, является продуктом слияния двух субъединиц TatC с двухдоменным линкером), тогда как у экстремально термофильного метаногена Methanopyrus kandleri эта субъединица вообще отсутствует.
Генеральная секреторная Sec-система. Второе название этой системы — «транслоказа» (рис. 59).
Секретируемый белок презентируется Sec-системе в развернутой форме. Для этого используется цитоплазматический шаперон SecB — тетрамер субъединиц молекулярной массой 15 кДа.
Основу транслоказы, изученной на примере Е. coli, составляет проводящий канал, или «транслокон». Он образован трансмембранными субъединицами SecY (49 кДа; 10 трансмембранных α-спиралей), SecE (14 кДа; три трансмембранные спирали) и SecG (11,5 кДа; две трансмембранные спирали). Четыре тримера субъединиц SecYEG образуют кольцо с внутренним каналом диаметром 1,5-5 нм.
Секретируемый белок проталкивается через транслокон димером амфипатических субъединиц SecA (молекулярная масса 102 кДа), которые выполняют рецепторную роль и обладают АТФазной активностью.
На периплазматической стороне транслоказы находятся вспомогательные субъединицы SecD и SecF. Вместе с субъединицами YajC и YidC они стабилизируют интегральные субъединицы SecA и способствуют транслокации.
При работе транслоказы Sec используются сразу два источника энергии — АТФ и Pmf.
Sec-система грамположительных бактерий сходна по составу и механизму действия с Sec-системой грамотрицательных бактерий, но они используют негомологичные молекулярные шапероны.
Кроме того, хотя у грамположительных бактерий нет ОМ, секреция не заканчивается после выхода белка на наружную поверхность СМ. Формирование третичной структуры белка и его выход во внешнюю среду зависят от интактного состояния клеточной стенки (в частности, от накопления в ней катионов Са2+). Поскольку грамположительные бактерии находят широкое применение в биотехнологии при производстве ферментов и гетерологичных белков, этот аспект их секреции важен в практическом отношении.
Субъединцы SecY, SecE и SecG транслоказы архей гомологичны бактериальным. Однако в археотной транслоказе отсутствует субъединица SecA, роль которой передается субъединицам SecD и SecF. Кроме того, у архей отсутствует «таргетинговый» шаперон SecB. В то же время у них выявлены компоненты SRP-пути (см. ниже), т. е. в очередной раз проявляется их сходство с эукариотами.
SRP-вариант генеральной секреторной Sec-системы. Как мы уже отмечали, у бактерий большинство секретируемых белков доставляется к транслокону шапероном SecB. Однако некоторые секретируемые белки высокогидрофоб- ны и служат интегральными компонентами СМ. В данном случае для их презентации Sec-системе используется не шаперон, а так называемый «SRP-комплекс» (рис. 59).
Рис. 59. Системы секреции белка. СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; Рmf— протондвижущая сила; SRР — «сигналраспознающая» частица; R —рибосома.
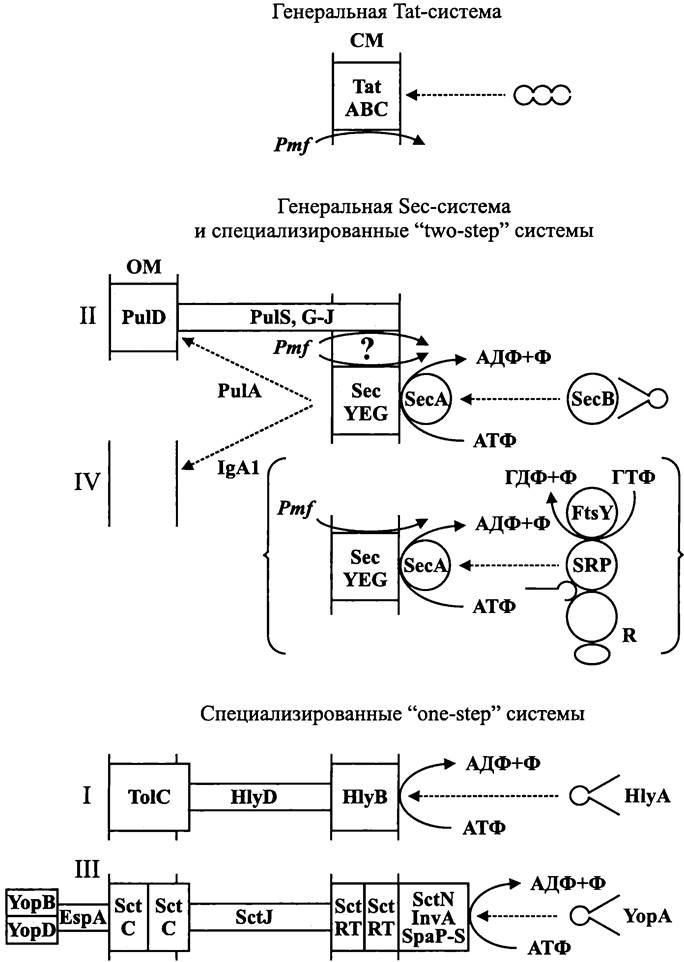
Его роль заключается в том, чтобы не позволить гидрофобным белкам преждевременно агрегировать в цитоплазме. Эукариотный гомолог этого комплекса, SRP54 обеспечивает мембранный таргетинг белков и их котрансляционную интеграцию в эндоплазматический ретикулум.
SRP (от англ. signal recognition particle; «сигналраспознающая» частица) представляет собой растворимый рибонуклеопротеиновый комплекс. Его каркас образует молекула 4,5SРНК, с которой связан полипептид Р48 молекулярной массой 48 кДа, или Ffh (от англ. fifty-four homolog, «гомолог эукариотного SRP-белка молекулярной массой 54 кДа»). Полипептид Р48 распознает сигнал, определяющий мембранный таргетинг секретируемого белка — высокогидрофобную сигнальную последовательность в незавершенной полипептидной цепи. В результате взаимодействия полипептида Р48 с рибосомой процесс элонгации приостанавливается (см. раздел 8.3.8).
Цикл работы SRP48 состоит из нескольких стадий. Вначале образуется бинарный комплекс, состоящий из рибосомы и SRP48. Затем к нему подключается растворимый белок FtsY, обладающий ГТФазной активностью (рис. 59). Такой тройственный комплекс с помощью белка FtsY фиксируется на СМ. Связанная с рибосомой полипептидная цепь (англ, ribosome-bound nascent polypeptide chain, RNC) высвобождается в канал транслокона SecYEG. После этого за счет энергии гидролиза ГТФ комплекс, состоящий из рибосомы, частицы SRP48 и белка FtsY, покидает мембрану, и цикл вновь повторяется.
Транслокация белка может осуществляться до или после терминации трансляции. В первом случае рибосома за счет энергии гидролиза ГТФ проталкивает полипептидную цепь через мембрану. Во втором случае это делает субъединица SecA, которая использует энергию гидролиза АТФ. Расходуется ли в обоих случаях Pmf — неизвестно.
Специализированная Sec-зависимая секреторная система II. Грамотрицательные бактерии могут использовать две специализированные секреторные системы, которые называются «Sec-зависимыми» — это система II и система IV.
Они действуют в два этапа (англ. two-step). На первом этапе белок при помощи генеральной Sec-системы транслоцируется на наружную поверхность СМ. При этом удаляется сигнальная последовательность, он освобождается в периплазматический компартмент, где шапероны поддерживают его в развернутой конформации. На втором этапе белок транслоцируется через ОМ и приобретает свернутую конформацию на ее наружной поверхности.
«Two-step» секреторная система II очень широко распространена у грамотрицательных бактерий. Самым известным результатом ее работы является экспорт пуллуланазы PulA — фермента, отрезающего боковые ветви от разветвленного амилопектина (который наряду с линейной амилозой входит в состав крахмала).
У Klebsiella oxytoca (сем. Enterobacteriaceae) в состав данного экспортера входят восемь основных субъединиц (PulCDF, PulK-O), которые кодируются pul-опероном.
Субъединица PulD, или «секретин» образует гидрофильную пору в ОМ. Субъединицы PulCF и PulK-O также расположены в СМ. Парадокс секреторной системы II заключается в том, что белок транслоцируется в периплазматический компартмент при помощи генеральной Sec-системы в развернутой форме, но где и как происходит его фолдинг, непонятно. Неизвестна также роль вспомогательных субъединиц (PulS и PulG-J), образующих мостик между двумя мембранами, хотя существует
предположение, что по нему от СМ передается энергия, которую необходимо затратить для выхода белка через секретиновую пору.
Секретины относятся к суперсемейству белков, формирующих мультимерные поры в ОМ. Через них экспортируются разнообразные субстраты — экзоферменты и токсины. Кроме того, через них из клетки выходят зрелые частицы нитчатых бактериофагов.
Все секретины образуют высокостабильные 12-субъединичные комплексы в виде кольца с центральным отверстием диаметром ~9,5 нм.
Между секретинами системы II (например, секретином PulD Klebsiella oxytoca и секретином XcpQ Р. aeruginosa) и системы III (например, секретином SctC Shigella lexneri и сектерином YscC Yersinia pestis) существует взаимная гомология. Это свидетельствует об общем эволюционном происхождении периферических доменов в двух секреторных системах, различающихся по механизму действия.
После выделения секретируемого белка в периплазматический компартмент он приобретает свернутую конформацию, близкую к нативной. В этом совместно участвуют периплазматические молекулярные шапероны, периплазматические тиол:дисульфидредуктазы Dsb-типа (см. раздел 8.4.2) и внутримолекулярные шаперонные домены секретируемого белка. По-видимому, третичная и даже четвертичная структура субстратного белка несет сигнал для прохождения через очень крупную пору секретина (в частности, через секретин EpsD экспортируется мультимерный токсин Vibrio cholerae).
Другие примеры секреторной системы II — это продукты out-оперона фитопатогенных бактерий рода Erwinia (секреция пектиназ и целлюлаз), xср-оперона Р. aeruginosa (секреция аминопептидазы, эластазы и экзотоксина А) и хрс-оперона Xanthomonas campestris (секреция пектиназы). Таким образом, с помощью данной системы грамотрицательные бактерии секретируют разнообразные гидролитические ферменты.
Специализированная Sec-зависимая секреторная система IV. Эта система была впервые обнаружена у патогенной бактерии Neisseria gonorhhoeae, которая секретирует протеазу IgA1. Позднее оказалось, что с ее помощью секретируются и другие факторы патогенности, в том числе адгезины (рис. 59).
Секреторную систему IV называют «автотранспортером» (англ. autotransporter), поскольку белок транслоцируется через ОМ самостоятельно, без помощи переносчиков. Дело в том, что С-концевой домен секретируемого белка сходен по структуре с поринами (см. раздел 8.4.1.2). Благодаря этому он спонтанно встраивается в ОМ и образует в ней бочковидную пору из 10-14 антипараллельных ß-цепей, через которую «продевается» секретируемый домен той же самой белковой молекулы. Так как пора имеет небольшой диаметр, секретируемый домен должен находиться в развернутой конформации, за что отвечает молекулярный шаперон (его роль может выполнять периплазматический шаперон или один из собственных внутренних доменов секретируемого белка). После выхода через пориновый домен на наружную поверхность клетки а-спиральный домен «вытягивает» его за собой и либо сохраняет с ним ковалентную связь, либо отрезается протеазой OmpT-типа (см. раздел
8.4.1.2) .
Специализированная секреторная система I. У грамотрицательных бактерий известны две специализированные секреторные системы, которые называют «Sec-независимыми» — это система I и система III.
В данном случае понадобится уточнение. Независимость от генеральной Sec-си- стемы выражается в том, что она не участвует в секреции субстратного белка, однако с ее помощью экспортируются субъединицы специализированой секреторной системы I, и в этом смысле зависимость сохраняется.
Специализированные Sec-независимые секреторные системы действуют одноэтапно (англ. one-step), и белок транслоцируется в развернутой конформации. Источником энергии для секреции служит гидролиз АТФ.
Особенность специализированной системы I заключается в том, что белок секретируется непосредственно в окружающую среду, без промежуточного высвобождения в периплазматический компартмент. По механизму действия секреторная система I напоминает мост, соединяющий цитоплазматический компартмент с экзо- плазматическим.
Примером специализированной секреторной системы I служит экспортер гемолизина HlyA Е. coli. Данная система относительно проста и состоит из субъединиц трех типов — Hlу В, HlyD и ТоLС.
Интегральный белок НlуВ присутствует в СМ в димерной форме. Он содержит мотив, характерный для представителей суперсемейства «транспортных» АТФаз ABC-типа (от англ. ATP-binding cassette), и обладает АТФазной активностью. Периферический белок HlyD, или «адгезионный» белок (англ. fusion protein) образует проводящий канал между СМ и ОМ. Установлено, что он димеризуется in situ. Интегральный белок ТЫС, или «выделительный» порин (см. раздел 8.4.1.2) состоит из β- спиральных цепей. Его тример образует гидрофильную пору внутри ОМ и частично выступает в периплазматический компартмент, где контактирует с «адгезионным» белком. Канал, проходящий через все три домена секреторной системы I, раскрывается только после связывания субстратного белка с субъединицами HlyBD. После завершения секреции канал опять закрывается.
Сигналом для мембранного таргетинга служит неудаляемый С-концевой участок полипептидной цепи, причем в роли рецептора выступает субъединица НlуВ.
С помощью разных вариантов специализированной секреторной системы I, предназначенных для отдельных групп субстратов, могут экспортироваться:
— экзоферменты (например, аденилатциклаза у возбудителя коклюша Bordetella pertussis, а также экзопротеазы у зоопатогенной бактерии Р. aeruginosa и фитопатогенной бактерии Erwinia chrysanthemi);
— токсины (в частности, лейкотоксин у Pasteurelia haemolytica);
— субъединицы поверхностного S-слоя;
— липоолигосахаридные Nod-факторы у симбиотических клубеньковых бактерий из сем. Rhizobiaceae.
Специализированная секреторная система III. Данная система играет центральную роль в патогенности многих грамотрицательных бактерий.
Ее важнейшая особенность состоит в том, что секретируемый белок доставляется непосредственно в цитоплазматический компартмент эукариотного хозяина — без промежуточного высвобождения в периплазматический компартмент бактерии и выхода в межклеточное пространство. Секретируемый белок одноэтапно транслоцируется через СМ, периплазматический компартмент, ОМ и СМ эукариотной клетки. Таким образом, по механизму действия секреторная система III напоминает шприц с иглой, и поэтому ее образно называют «инъектосомой» (англ. injectosome).
Субстратами для секреторной системы III служат «эффекторы» и «транслокаторы». Те и другие содержат уникальные неудаляемые сигнальные последовательности, расположенные в разных участках полипептидной цепи.
Эффекторы, хотя и сильно различаются по структуре и вызываемому ими клеточному ответу, обязательно имеют гомологичные участки с эукариотными белками. Оказавшись в цитоплазматическом компартменте эукариотной клетки, эффектор, примером которого служит белок возбудителя чумы YopA (от англ. Yersinia outer
protein), модифицирует регуляторные каскады, разрушает цитоскелет и вызывает апоптоз.
Транслокаторы играют вспомогательную роль и образуют пору в СМ эукариотной клетки, через которую эффекторы проходят в ее цитоплазматический компарт- мент.
Секреторная система III имеет сложное строение и содержит до 25 типов субъединиц. Биосинтез и сборка ее компонентов, а также секреция белка контролируются внешней средой. В частности, условием секреции эффектора YopA является низкое содержание катионов Са2+ и температура выше 37°С. Кроме того, условием ее запуска является контакт с клеткой-мишенью, а для этого требуются специальные адгезины.
Для активации транскрипции компонентов секреторной системы III используются регуляторные белки семейства AraC/XylS. В свою очередь, в регуляции транскрипции секретируемых белков участвуют мультифункциональные молекулярные шапероны из семейства TTS (от англ. type three secretion; 15 кДа). Кроме того, они поддерживают секретируемый белок в развернутой конформации, а также выполняют роль стабилизирующих факторов.
Секреторная система III состоит из трех основных частей — базального тела, «иглы» и транслокаторного домена (рис. 59). Она охарактеризована на примере Salmonella entéricaсеротип Typhimurium, Shigella flexneri, Yersinia enterocolitica, Y. pestis и Y. paratruberculosis, а также энтеропатогенных штаммов Е. coli.
Базальное тело интегрировано в клеточную оболочку и включает в себя:
— проксимальный домен, который находится в СМ (два кольца диаметром 40 нм, образованные субъединицами SctR и SctT); на его внутренней стороне находится экспортный аппарат, состоящий из субъединиц InvA и SpaP-S, а также из субъединицы SctN, относящейся с суперсемейству «транспортных» АВС-АТФаз;
— осевой цилиндр, который пересекает периплазматический компартмент; внутри него проходит канал диаметром 2 нм (субъединицы SetJ);
— дистальный домен, или секретин, который пронизывает муреиновый саккулус и встроен в ОМ (два кольца диаметром 20 нм, образованные субъединицами SctC).
Девять генов, кодирующих субъединицы базального тела и его сборку, высококонсервативны, причем восемь из них гомологичны генам, участвующим в биогенезе жгутика (см. ниже).
Игла служит продолжением базального тела и состоит из белка EspA. Эта трубчатая структура имеет диаметр ~8 нм и длину до 80 нм. Она выполняет роль адгезионного мостика между бактерией и клеткой-мишенью. Кроме того, она является наружной частью канала, по которому передаются транслокаторы и эффекторы.
Транслокаторный домен расположен на конце иглы. Он представляет собой мультимерный комплекс, образованный двумя белками (IрА и IрС у Sh. flexneri, YopB и YopD у Y. enterocolitica и т. д.).
Помимо патогенов человека и животных, секреторная система III имеется у некоторых фитопатогенных бактерий, которые также доставляют белковые факторы вирулентности непосредственно в клетку хозяина. В данном случае инъектосоме приходится преодолевать клеточную стенку, которая по толщине (>100 нм) значительно превышает клеточную оболочку млекопитающих. Поэтому конец иглы наращиваются за счет присоединения к нему фимбрии Нгр-типа (см. раздел 8.6.3).
Секреторная система III типа служит не только для экспорта белковых факторов вирулентности. Она используется в аппарате Mrf (от англ. mating pair formation),
который формирует партнерские пары при конъюгации. Наконец, при генетической трансформации патогенных бактерий родов Campylobacter и Helicobacter она может захватывать «голую» ДНК, происходящую из клетки гетерологичной бактерии.
Специализированная секреторная система III как гомолог и функциональный аналог базального тела бактериального жгутика. В конце 1990-х годов выяснилось, что специализированная секреторная система III проявляет сходство с широко распространенной локомоторной системой — бактериальным жгутиком, точнее, его базальным телом (см. раздел 8.6.2).
Доказано не только то, что эти системы имеют внешне сходное строение, но и то, что их субъединицы кодируются гомологичными генами. В настоящее время обе структуры рассматриваются как варианты специализированного компартмента — секреторной системы III.
Обеспечивающая вирулентность секреторная система III и входящая в жгутиковый аппарат секреторная система III не исключают друг друга. В частности, патогенные протеобактерии Bordetella sp. (сем. Alcaligenaceae) и Р. aeruginosa (сем. Pseudomonadaceae), а также виды родов Escherichia, Salmonella, Shigella и Yersinia (сем. Enterobacteriaceae) обладают генами обеих систем.
В то же время известны случаи, когда у патогенных бактерий имеется только «вирулентная» секреторная система III. Примером служат облигатные внутриклеточные паразиты из рода Chlamydia. У них на поверхности клетки расположены в гексагональном порядке 13-30 «рожек» (англ. projections) длиной 70-90 нм и диаметром 5-10 нм. Эти структуры морфологически сходны с секреторной системой III. В геноме хламидий обнаружены гомологи генов, кодирующих большинство ее субъединиц. Предполагается, что «рожки» служат каналами для транспорта метаболитов из клетки хозяина в клетку паразита и, возможно, каналами для взаимного сигналинга.
У некоторых грамотрицательных патогенов, например, Helicobacter sp. (сем. «Helicobacteraceae») и Vibrio sp. (Vibrionaceae), а также у всех грамположительных патогенов секреторная система III представлена только «жгутиковым» вариантом. Для экспорта факторов вирулентности они используют другие секреторные системы.
Наконец, жгутиковый аппарат Y. enterocolitica обладает уникальной способностью секретировать в среду фактор вирулентности — липопротеин YlpA.
В отличие от жгутиковых генов, которые кодируются хромосомой, гены «вирулентной» секреторной системы III в большинстве случаев находятся на плазмидах в островках патогенности (см. раздел 8.6.3.3) и могут «горизонтальным» путем передаваться от одной бактерии к другой.
При совместном кодировании обеих систем они обычно имеют противоположные векторы регуляции и одновременно не экспрессируются. Как правило, на инвазивной стадии инфекции бактерии образуют только «жгутиковую» секреторную систему III. Логика этого явления состоит в том, что необходимость в «вирулентной» системе III возникает только когда патоген вступает в прямой контакт с клеткой хозяина. Регуляторным сигналом, оказывающим негативное влияние на экспрессию системы подвижности, служит повышение температуры до 37° С.
8.6.2. Жгутики
Прокариотный жгутик, или «флагелла» (лат. flagellum — бич) представляет собой многофункциональный специализированый компартмент со смешанной локализацией. Однако его главная задача — быть локомоторной органеллой.
В любой нише мы встречаем как пространственные, так и темпоральные градиенты физических и химических параметров среды. Многие, хотя далеко не все, прокариоты обладают подвижностью, что позволяет им изменять свое положение в окружающей среде и попадать в более благоприятные условия. Помимо этого, активная подвижность дает возможность ряду бактерий скапливаться в многоклеточные агрегаты, внутри которых образуются покоящиеся клетки, что позволяет переносить неблагоприятные, а иногда и экстремальные условия окружающей среды.
8.6.2.1. Типы подвижности у прокариотов
Между подвижностью бактерий и архей имеется большая разница. В частности, у бактерий больше способов подвижности. При этом самый распространенный локомоторный аппарат, жгутик, имеет разное происхождение у бактерий и архей. При внешнем сходстве с бактериальным, жгутик архей устроен иначе; различия также касаются механизма биогенеза и кинематики. Короче, перед нами еще один пример того, как архей мимикрируют под бактерий.
Для описания типов подвижности у прокариотов нам потребуется специальная терминология.
Термином «подвижность» (англ. motility) обозначают изменение пространственного расположения отдельной клетки или трихома, а также группы клеток, соприкасающихся друг с другом.
Клеточная подвижность обеспечивается внутренними органеллами клетки или взаимодействием поверхностных структур с окружающей средой. Подвижность не означает любое перемещение в окружающей среде, которое, в частности, может происходить за счет броунова движения, конвекции или водного потока.
Общая схема типов подвижности у прокариотов приводится на рис. 60.
Рис. 60. Классификация типов подвижности у прокариотов.
Активная и пассивная подвижность. При активной подвижности клетка совершает работу по собственному перемещению, на что расходуются внутренние запасы энергии.
Пассивная подвижность не требует от клетки энергетических затрат. Она может осуществляться в результате спонтанной разборки или сборки филаментозных структур, соединяющих клетку с субстратом, а также в результате изменения плавучести клетки.
Подвижность в жидкой фазе и на границе раздела жидкой и твердой фаз. Насколько успешно тот или иной тип подвижности может быть использован бактерией, зависит от структуры внешней среды.
В случае, когда клетка перемещается в жидкой среде и локомоторными органеллами служат экзоплазматические жгутики, периплазматические жгутики или мембранные структуры, мы говорим о «плавании» (англ. swimming).
В случае, когда клетка или трихом перемещается по границе раздела жидкой и твердой фаз, речь идет о «скольжении» (англ. gliding).
Индивидуальная и коллективная подвижность. При индивидуальной подвижности (англ. adventurous motility; «авантюристическая подвижность») особь плавает в жидкой фазе, перемещается рывком или скользит, находясь в контакте с твердой фазой.
При коллективной подвижности (англ. social motility) колония клеток движется по увлажненной поверхности твердого субстрата. Если такое перемещение осуществляется с помощью жгутиков, оно называется «роением» (англ. swarming), а если с помощью фимбрий IV типа или с помощью мембранных структур — коллективным скольжением (англ. social gliding).
Подвижность с помощью жгутиков (активная). Плавание при помощи экзоплазматических жгутиков широко распространено у бактерий и архей, тогда как роение наблюдается только у бактерий. Плавание благодаря периплазматическим жгутикам является уникальным свойством представителей филы BXVII Spirochaetes.
Строение и биогенез жгутиков, а также обеспечиваемые ими типы подвижности мы подробно рассмотрим в разделах 8.6.2.2 и 8.6.2.3.
Подвижность с помощью мембранных структур (активная). Данный тип подвижности основан на усилии, которое передается от ОМ и/или СМ на жидкий или твердый субстрат.
В случае жидкого субстрата это означает плавание без участия жгутиков. Такой тип подвижности встречается крайне редко. Впервые он был описан в 1985 г. у одноклеточной цианобактерии рода Synechococcus, входящей в состав пикопланктона тропической зоны Мирового океана. При отсутствии морфологически выраженных локомоторных органелл эти бактерии перемещаются со скоростью ~10 мкм • с-1, что соответствует скорости скольжения, однако в несколько раз ниже скорости плавания. Во второй половине 1990-х годов было показано, что данный тип подвижности обеспечивается поверхностным белком SwmА, который содержит характерный мотив в виде Са2+-связывающего нонапептида. Интересно, что многочисленные повторы Са2+-связывающего нонапептида имеются и в молекуле осциллина (см. ниже).
Роль белка SwmA в квази-плавании Synechococcus sp. остается загадкой, поскольку мутанты, у которых он отсутствует, лишаются способности к перемещению, но могут вращаться.
Другой пример плавания без участия жгутиков относится к спирально извитым бактериям из рода Spiroplasma (фила ВХIII Firmicutes, класс «Mollicutes»), у которых нет ни ОМ, ни ригидного слоя. В невязкой жидкости эти бактерии вращаются, а в гелеобразной среде совершают поступательные движения. Механизм подвижности, в которой участвуют ундулирующая (лат. iinda — волна) ОМ или ундулирующая СМ, точно неизвестен, хотя предполагается, что он связан с поверхностными фибриллами, конформационное изменение которых передается на СМ.
По поверхности твердого субстрата подвижность происходит благодаря комплексу СМ/ОМ. Примером служат «авантюристически» скользящие клетки Myxococcus xanthus (класс «Deltaproteobacteria»). Хотя механизм такой подвижности точно не известен, предполагается, что в нем участвует «Ton-зависимая» система, которая изменяет конформацию ОМ за счет энергии Pmf (см. II том учебника). Показано, что за подвижность отвечает ген aqIU; его продукт входит в состав ОМ и на 35% гомологичен белку ТolС Е. coli — компоненту специализированной секреторной системы I типа. Напомним, что эта система образует мост между СМ и ОМ, и за счет энергии гидролиза АТФ белок выходит через отверстие порина (см. раздел 8.6.1).
В конце 1990-х годов была предложена модель для объяснения механизма «авантюристического» скольжения бактерий из родов Cytophaga и Flavobacteium (фила ВХХ Bacteroidetes). Согласно этой модели, в составе ОМ есть белки, обеспечивающие адгезию к твердому субстрату. В свою очередь, белки СМ используют энергию Pmf для того, чтобы сместить ОМ вдоль специальных локусов муреинового саккулуса. Это напоминает машину на гусеничном ходу. Естественно, что для эффективного скольжения необходима координация работы «гусеничных катков», или отдельных систем, преобразующих хемиосмотическую энергию в механическую.
Подвижность с помощью эндоскелета (активная). Данный тип подвижности представляет собой один из вариантов скольжения и связан с механическим взаимодействием эндоскелета с твердым субстратом, например, поверхностью эпителиальной ткани. Он описан у микоплазм, в частности — Mycoplasma pneumoniae, и пока мало изучен.
Подвижность с помощью эндоскелета осуществляет «терминальная органелла». Это вырост СМ, внутри которого находится цитоскелет, состоящий из HMW-белка и вспомогательных белков (см. раздел 8.3.9.1).
Подвижность с помощью реактивной струи (активная). Данный тип подвижности был впервые изучен в конце 1980-х годов у осциллаториевой цианобактерии «Phormidiumuncinatum». Путем выброса реактивной струи трихомы могут скользить со скоростью до 10 мкм-с-1.
Механизм этого типа выглядит следующим образом. Через «комплекс контактной поры» (см. раздел 8.4.3.1) из цитоплазматического компартмента со скоростью 1,2 • 104 - 4 • 104 кДа • с-1 на поверхность клетки выдавливается, по мере ее биосинтеза, слизь, состоящая из экзополисахарида (см. раздел 8.5.4.1).
Поверх ОМ и S-слоя по оси трихома пролегают спиральные каналы, образованные фибриллами гликопротеина осциллина (англ. oscillin, 66 кДа). В области перегородки между соседними клетками находятся два кольца выходных отверстий «комплекса контактной поры». Отверстия одного кольца направлены в сторону условного переднего конца трихома, а отверстия второго кольца в сторону условного заднего конца трихома. Отверстия открываются в осциллиновые каналы, борта которых используются как резьба винта, а слизь выполняет роль реактивной струи. Направление перемещения трихома зависит от того, через какую серию отверстий выдавливается слизь.
Подвижность за счет полимеризации актина (пассивная). В данном случае патогенная бактерия, проникшая в эукариотную клетку, индуцирует полимеризацию хозяйского актина (рис. 61). Данный тип подвижности является важнейшим фактором вирулентности факультативных внутриклеточных паразитов Listeria monocytogenes (фила ВХIII Firmicutes) и Shigellaflexneri (класс «Gammaproteobacteria»), а также облигатных внутриклеточных паразитов из рода Rickettsia. Способностью полимеризовать актин обладают даже некоторые вирусы.
Рис. 61. Система для полимеризации хозяйского актина, используемая в целях подвижности факультативным внутриклеточным паразитом Listeria monocytogenes.
N и С — терминальные домены поверхностного бактериального белка ActA; VASP — профилинсвязывающий хозяйский белок; Агр2 и Агр3 — вспомогательные хозяйские белки; Р и В — «заостренный» и «зазубренный» концы актиновой нити.

Внутриклеточные паразитические бактерии подразделяются на тех, которые размножаются в вакуолях хозяина (рода Chlamydia, Coxiella, Legionella, Mycobacterium, Salmonella и др.), и тех, которые размножаются в его цитоплазме (рода Listeria, Shigella, Rickettsia и др.). Кроме того, внутриклеточные паразиты могут поселяться в фагоцитирующих или в нефагоцитирующих клетках. В первом случае они остаются в фагосоме или разрушают ее мембрану и переходят в цитоплазму.
Некоторые патогенные бактерии, в частности Listeria и Shigella способны проникать в нефагоцитирующие клетки. Для этого у них существуют особые приспособления, которые влияют на цитоскелет хозяина и индуцируют эндоцитоз. В частности, L. monocytogenes образует белок интерналин IntA (от англ. internalin), который связывается с поверхностью энтероцитов. Роль рецептора интерналина играет Са2+-зависимый белок Е-кадгерин (англ. cadherin). В норме он входит в состав опоясывающих десмосом (гомотипическая адгезия), которые обеспечивают поляризацию эпителиальных клеток и общую архитектонику ткани.
После эндоцитоза паразит с помощью очень активных фосфолипаз разрушает мембрану фа- голизосомы и переходит в цитоплазму хозяина, где он размножается и полимеризует вокруг себя хозяйский актин. Затем оболочка из актиновых нитей превращается в «хвост кометы» длиной 10- 40 мкм, на котором бактерия, как на лифте, продвигается по хозяйской цитоплазме со скоростью 6-60 мкм • мин-1, а затем внутрь соседней клетки проникает протуберанец длиной до 25 мкм. Его окончание вместе с находящейся в нем бактерией превращается в вакуоль, окруженную двумя концентрическими мембранами. Мембраны разрушаются под воздействием фосфолипаз и т. д. Таким образом, бактерия колонизирует ткани, не покидая цитоплазму хозяина, что позволяет избежать действия его защитных систем.
Актин представляет собой глобулярный белок, или G-актин (от англ. globular) с молекулярной массой 43 кДа, способный полимеризоваться в длинные нити F-актина (от англ. filamentous). Актиновая нить анизотропна — ее сборка блокируется на «заостренном» конце (англ. pointed end) и происходит на «зазубренном» конце (англ. barbed end).
Полимеризация хозяйского актина зависит от поверхностных белков ActА (у L. monocytogenes) или IcsA (у S. flexneri), которые накапливаются на одном из полюсов клетки.
N-концевой домен белка ActА содержит актин-связывающие сайты (рис. 61). В его центральном домене имеются повторы Pro-Glu, которые узнают хозяйский белок VASP (от англ. vasodilator- stimulated phosphoprotein; ~50 кДа). В свою очередь, комплекс ActA/VASP связывает хозяйский белок профилин (англ. profilin; 16 кДа). Роль профилина, по-видимому, заключается в том, что он поставляет G-актин на «зазубренный» конец F-актиновой нити при условии, что последний не экранируется кэп-белком. Во взаимодействии белка ActA с мономерным актином, по-видимому, участвуют хозяйские белки Агр2/Агр3 (от англ. actin-related protein). Смысл его заключается не в том, что ActA непосредственно полимеризует актин, а в том, что этот белок служит площадкой для вспомогательных белков (VASP, профилина и Агр2/Агр3), а также декэппирует конец актиновой нити, т. е. поддерживает ее в «зазубренном» состоянии.
После завершения полимеризации F-актин кэппируется, между отдельными нитями с помощью α-актинина образуются поперечные сшивки, и из них образуется «хвост кометы».
Подвижность в результате изменения плавучести (пассивная). Данный тип подвижности позволяет клеткам перемещаться вертикально в толще водной среды. В результате они попадают в горизонт, наиболее подходящий для жизнедеятельности.
Известны следующие альтернативные типы подвижности за счет изменения плавучести — флотация и погружение. В первом случае прокариотная клетка образует газовые везикулы, становится легче воды и всплывает (см. раздел 8.3.11.2). Во втором она накапливает полиглюкозидные гранулы, становится тяжелее воды и погружается (см. раздел 8.3.11.4).
Миграции в противоположных направлениях происходят с помощью регуляторной петли обратной связи в результате уменьшения количества этих органелл.
Подвижность, осуществляемая за счет втягивающихся фимбрий IV типа (пассивная). Строение и биогенез фимбрий IV типа, а также обеспечиваемые ими типы подвижности будут рассмотрены в разделе 8.6.3.
8.6.2.2. Жгутики бактерий
Используя жгутики, бактерии индивидуально плавают в жидком субстрате или переходят к коллективному роению по увлажненной поверхности твердого субстрата.
Подсчитано, что жгутики образуют до 80% известных видов бактерий. Это указывает на высокую биологическую целесообразность данных структур. Прежде всего, передвигаясь с их помощью, бактерии могут проникнуть в область с более благоприятными условиями существования. Наряду с фимбриями жгутики способствуют образованию биопленок (см. ниже). Благодаря жгутикам Geobacter metallireducens вступает в контакт с илом или минеральными частицами, где содержатся слабо растворимые катионы Fe3+, используемые в качестве акцептора электронов при анаэробном дыхании. Жгутики облегчают колонизацию хозяина симбиотическими бактериями (примером служат клубеньковые бактерии из рода Rhizobium). Для инвазивных патогенов жгутики являются одним из факторов вирулентности, поскольку позволяют проникнуть в тело хозяина, перемещаться в вязком межклеточном веществе соединительной ткани и проходить через щели между клетками эндотелия.
Обладание жгутиками имеет и негативную сторону. Они состоят из разных субъединиц, на биосинтез которых уходит много строительного материала, а для сборки и вращения жгутиков требуется много энергии. Неслучайно при отсутствии конкурентного давления бактерии теряют подвижность и/или перестают образовывать жгутики. Неслучайно также, что у патогенных бактерий сигналом к прекращению биогенеза жгутиков служит повышение наружной температуры: «Замри — ты находишься внутри макроорганизма». Кроме того, жгутики высоко иммуногенны, что ставит бактерию под удар защитных сил макроорганизма. Жгутики — это ахиллесова пята бактериальной клетки. Этим пользуются на практике при активной иммунизации с помощью препаратов жгутиков, а также при пассивной иммунизации антижгутиковыми антисыворотками или антителами. В последнее время с помощью генноинженерных методов создают гибридные жгутиковые вакцины, когда в молекулу флагеллина вводят эпитопы других инфекционных агентов, в частности слабо имммуногенного вируса гриппа.
Жгутик как специализированный компартмент со смешанной локализацией; устройство и работа жгутика. Хорошо известно, что эукариотный жгутик, или «ундулиподия» производит биения, посылает бегущую волну или описывает телесный угол, используя энергию гидролиза АТФ. В структурном плане он представляет собой вырост клетки с цитоскелетом внутри.
В отличие от него, прокариотный жгутик вмонтирован в клеточную оболочку и свободно вращается в ней за счет энергии трансмембранного градиента катионов Н+ или Na+. Такие биокинематические системы типа «колеса», состоящие из двух независимых частей, статора и ротора, известны только в мире прокариотов (другими примерами являются FoFi-АТФаза и, возможно, фимбрия IV типа).
Жгутик попеременно вращается по часовой стрелке или против часовой стрелки. Частота вращения постоянна для конкретного объекта и находится в пределах 250- 1700 Гц (1,5 • 104-105 оборотов • мин-1). С помощью жгутиков Е. coli и многие другие бактерии плавают со скоростью около 50 мкм • с-1 и, следовательно, за секунду преодолевают расстояние, равное 20 длинам их тела. Такую же относительную скорость (20 м • с-1) развивает только тунец Tunnus albacus. Для того, чтобы сравниться по относительной скорости с самыми быстрыми вибрионами, плавающими со скоростью до 200 мкм • с-1, человеку пришлось бы перемещаться со скоростью самолета (600 км • ч-1).
Основная информация о строении и работе бактериального жгутика получена на примере Е. coli и Salmonella entérica серотип Typhimurium. «Плавательные» гены у этих бактерий традиционно относят к трем основным группам:
— flа-гены (от англ. flagellum — жгутик), кодирующие субъединицы и биогенез жгутика; в настоящее время по последовательному положению на генной карте их обозначают как flg-гены, flh-, fli- и flj-гены;
— mot-гены (от англ. motility — подвижность), отвечающие за вращение мотора жгутика;
— che-гены (от англ. chemotaxis —хемотаксис), обеспечивающие плавание как адаптивную реакцию.
Наибольшее число генов (около 50) входит в группу fia. Из них около половины кодируют субъединицы жгутика. Гены flgM, flhCD и fliA регулируют биогенез жгутика.
Грамотрицательные бактерии чаще всего имеют один или несколько «экзоплаз- матических» жгутиков (англ. exoplasmic flagella). Легко заметить, что этот термин неточно отражает локализацию жгутиков, поскольку они расположены одновременно в цитоплазматическом, периплазматическом и экзоплазматическом компартментах.
Грамотрицательные бактерии из филы BXVII Spirochaetes обладают «периплазматическими» жгутиками (англ. periplasmic flagella). И этот термин неточен, поскольку такие жгутики располагаются одновременно в цитоплазматическом и периплазматическом компартментах. Особым вариантом периплазматических жгутиков являются «зачехленные» жгутики (англ. sheathed flagellum) вибриоидных бактерий Bdellovibrio bacterioviorus, Vibrio cholerae, V. parachaemolyticus и Helicobacter sp. Они покрыты выростом ОМ, отличающимся по составу от ее остальных участков.
Наконец, экзоплазматические жгутики грамположительных бактерий расположены одновременно в цитоплазматическом и экзоплазматическом компартментах.
Бактериальный жгутик, строение которого мы рассмотрим на примере жгутика грамотрицательных бактерий, состоит из трех частей (рис. 62, А-В):
— «нити», или жесткой филаментозной стуктуры длиной до 15 мкм и диаметром ~20 нм, которая расположена за пределами цитоплазматического компартмента (рис. 62, А);
— базального тела, которое встроено в СМ и имеет дополнительную опору; ее роль у грамотрицательных бактерий играет муреиновый саккулус совместно с ОМ, а у грамположительных бактерий только муреин-тейхоатный саккулус (рис. 60, В);
— «крюка» длиной ~55 нм и диаметром ~30 нм, или гибкого сочленения между нитью и базальным телом (рис. 60, Б).
В состав базального тела, в числе прочего, входит секреторная система. Она относится к специализированным секреторным системам III (см. раздел 8.6.1) и при сборке жгутика экспортирует субъединицы базального тела, крюка и нити (рис. 62, Г).
Рис. 62. Строение бактериального жгутика. А — нить; Б — крюк; В — базальное тело; Г — секреторный аппарат; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; М — муреиновый саккулус.

Клетка, имеющая жгутик(и), получает двигательный импульс в результате крутящего взаимодействия нити с субстратом — напрямую в случае экзоплазматических жгутиков или через ОМ в случае периплазматических жгутиков. Хотя нить многократно превышает по размеру все остальные части жгутика, она не служит источником двигательного импульса, который исходит от работающего молекулярного мотора.
Мотор жгутика включен в состав базального тела, которое погружено в клеточную оболочку и частично выдается в цитоплазматический компартмент. Ротор мотора вращается за счет энергии протонного градиента (Pmf) или, в случае морской бактерии Vibrio alginolyticus и алкалифильных штаммов Bacillus sp., за счет энергии градиента катионов Na+ (Smf). Энергия АТФ вообще не используется для работы жгутика. Тем не менее, она необходима для систем его сборки, а также для систем, регулирующих направление его вращения попеременно по часовой стрелке или против часовой стрелки.
Нить. Рабочей частью жгутика является жесткая нить (англ. filament). Она имеет вид спирализованного цилиндра с внутренним каналом диаметром ~3 нм. Нить
состоит из ~2-104 идентичных субъединиц флагеллина (FliC; ~50 кДа). В очень редких случаях она образована сразу несколькими типами флагеллина — например, у Helicobacter pylori(два флагеллина, расположенные на противоположных концах нити), Caulobacter crescentus и Rhizobium meliloti (два флагеллина, расположенные на противоположных концах нити), а также у Campylobacter coli (два флагеллина, расположены поочередно).
Недавно выяснилось, что у ряда бактерий (Aeromonas caviae, Campylobacter jejuni, C. crescentus, H. pylori и P. aeruginosa) флагеллины гликозилированы.
В результате рентген-кристаллографического исследования при уровне разрешения 2Â, проведенного в начале 2000-х годов, было показано, что субъединицы флагеллина по форме напоминают бумеранг. Одно плечо образует внутреннюю выстилку, а другое — стенку канала нити. Флагеллины бактерий взаимно гомологичны, причем наиболее консервативным участком является центральный домен молекулы.
Флагеллиновые субъединицы имеют спиральную укладку, по 11 субъединиц на оборот спирали. Нить свернута в суперспираль, диаметр и длина волны которой зависят от природы объекта. Флагеллиновые субъединицы образуют 11 параллельных протофиламентов, проходящих под небольшим уклоном к оси нити. Флагеллин упаковал двумя способами — в виде «коротких» R-npo- тофиламентов (от англ. right — правый) и «длинных» L-протофиламентов (от англ. left — левый), в результате чего нить приобретает закрученную форму, причем «короткие» протофиламенты идут по внутренней стороне большой спирали.
Форма нити определяется аминокислотной последовательностью флагеллина, ионной силой и pH раствора. Кроме того, она зависит от скручивающего усилия, приложенного к нити со стороны мотора. При вращении мотора против часовой стрелки нить принимает форму левозакрученной спирали. Когда мотор переключается на вращение по часовой стрелке, нить изменяет свою конформацию и превращается в правозакрученную спираль. Именно этим определяются два типа движения политрихиальных бактерий в жидкой среде — «плавание» и «кувыркание», поскольку левозакрученные спирали при вращении против часовой стрелки сплетаются в жгут и работают согласованно, а правозакрученные спирали при вращении по чатовой стрелке расплетаются и тянут клетку в разные строны.
Канал нити очень узкий, и молекулы флагеллина проходят через него по очереди. При этом они находятся в частично развернутой конформации. На дистальном конце нити расположен кэп (рис. 62,А; вставка). Он имеет форму диска и состоит из пяти субъединиц FHD (50 кДа). Дистальный домен пентамера имеет вид плоской пятиугольной площадки, а проксимальный домен — пяти гибких ножек. Благодаря этому в кэпе образуется пять окошек, через которые субъединицы флагеллина выходят наружу. Кэп придает им свернутую конформацию. Таким образом, кэп играет уникальную роль — вращающегося экзоплазматического шаперонина. За счет освобожденной энергии кэп «переступает» вверх по спирали нити, пропускает следующую молекулу флагеллина и т. д.
Крюк. Нить соединяется с базальным телом при помощи крюка (англ, hook), состоящего из субъединиц FlgE, вспомогательных белков FlgKL, а также кэппирующего белка FlgD. По завершении сборки крюка кэп удаляется. В отличие от нити, протофиламенты крюка при его вращении непрерывно меняют конформацию, переходя от «длинной» упаковки к «короткой». Крюк играет роль гибкого сочленения, что позволяет жесткой нити изменять свое положение по отношению к поверхности клетки. Благодаря этому отдельные жгутики могут сплетаться в жгут.
Базальное тело. Базальное тело (англ. basal body) встроено в клеточную оболочку. В случае экзоплазматических жгутиков грамотрицательных бактерий основными частями базального тела являются (рис. 62):
— «ось» (англ. rod; субъединицы FlgBCFG и FliE), которая в своей дистальной части связана с крюком и нитью, а в проксимальной части с MS-кольцом;
— «L-кольцо» (от англ. lipopolysaccharide; субъединицы FlgH), которое интегрировано в ОМ и выполняет роль поддерживающей втулки для вращающейся оси,
— «P-кольцо» (от англю peptidoglycan; субъединицы Flgl), которое интегрировано в муреиновый саккулус и также выполняет роль поддерживающей втулки для вращающейся оси,
— «MS-кольцо» (от англ. membrane stator; 26 субъединиц FliF), на котором смонтированы проксимальные субъединицы оси (FliE), субъединицы ротора (FHG), переключатель направления вращения (37 субъединиц FUM и 110 субъединиц FHN), а также экспортный аппарат;
— «ротор» (англ. rotor; 44 субъединицы FliG), расположенный в основании SM- кольца;
— «С-кольцо» (от англ. cytoplasmic; субъединицы FliMN), которое связано с ротором и играет роль переключателя направления вращения.
Экзоплазматические жгутики грамположительных бактерий не имеют L- и Р- колец. Роль поддерживающей втулки для вращающегося стержня у них выполняет мощный муреин-тейхоатный саккулус.
Заметим, что название «MS-кольцо» не отражает роли данной структуры в работе мотора, поскольку в действительности статором сужит комплекс мембранных белков MotAB.
Субъединицы MotAB уложены по кругу внутри СМ и образуют 10-16 «полюсов». В молекуле белка MotB имеются четыре интегральные α-спирали и большой периферический С-концевой домен, нековалентно связанный с муреиновым саккулусом; благодаря ему белок MotА заякоривается в ригидном слое клеточной стенки.
Статорные субъединицы MotAB приводят во вращение ротор FHG, используя энергию Pmf. Предполагается, что скручивающее усилие возникает следующим образом.
Когда протоны движутся в цитоплазму по хемиосмотическому градиенту, они нейтрализуют отрицательный заряд карбоксильных групп Asp32 в димере субъединиц MotB. Вследствие этого изменяется конформация полипептидных цепей в тетрамере субъединиц MotA. В свою очередь, это влияет на электростатическое взаимодействие между субъединицами MotAи С-концевыми доменами роторных субъединиц FliG, которые связаны с проксимальной частью MS-кольца (рис. 62).
По аналогии с электромотором, «гладкое» вращение ротора жгутика складывается из отдельных шагов, причем каждый раз комплекс MotAB взаимодействует с противостоящей ему субъединицей FliG. На каждом шагу в цитоплазму переносятся два протона, а за полный оборот жгутика в цитоплазматический компартмент переходят ~1,2103 протонов.
У морской бактерии V. alginolyticus, использующей для вращения жгутика Smf, роль статорных субъединиц выполняют гомологи MotAB — белки РотАВ.
С-кольцо состоит из субъединиц FliMN. Фосфорилированный регулятор ответа CheY-P связывается с субъединицей FHM, и жгутик переходит от вращения против часовой стрелки к вращению по часовой стрелке.
Регуляция биосинтеза и сценарий сборки бактериального жгутика. Биогенез жгутика «дорого обходится» бактерии и поэтому находится под контролем регуляторных систем, реагирующих на условия окружающей среды. Жгутиковые гены локализуются в хромосоме и объединяются в опероны, 15 из которых отвечают за биосинтез, а 17 — за функционирование жгутика.
Биосинтез жгутикового аппарата контролируется регуляторным каскадом, который охарктеризован у Е. coli и Salmonella enterica серовар Typhimurium. Он состоит из трех классов генов.
К генам I класса принадлежит мастер-оперон flhDC. Его продукт, белок FшрD2C2 активирует транскрипцию fli-генов более низкой иерархической ступени.
Ген II класса fliA кодирует белок RpoF (σ28), активирующий транскрипцию жгутиковых генов.
Гены III класса, или собственно жгутиковые гены позитивно регулируются сигма-фактором σ28 и негативно регулируются антисигма-фактором FlgM.
Белок FlgM сохраняется до конца сборки базального тела и крюка, а перед встраиванием субъединиц FlgLK удаляется из клетки. Это позволяет сигма-фактору σ28 активировать транскрипцию генов III класса, которые кодируют флагеллин, кэп и белки, отвечающие за хемотаксис.
Для экспорта субъединиц жгутика и белков, регулирующих их сборку, используются сразу две системы экспорта белка — генеральная Sec-система и «жгутиковая» секреторная система III.
Первая из них экспортирует те субъединицы, которые являются интегральными компонентами СМ, а также субъединицы периплазматических частей жгутика.
Остальные субъединицы экспортируются с помощью секреторной системы III типа по каналу, проходящему через MS-кольцо, крюк и нить.
Сборка жгутика проходит в 12 этапов.
1. На первом этапе с помощью генеральной Sec-системы экспортируются субъединицы MS- кольца (FHF).
2. На втором этапе с ее же участием переносятся субъединицы экспортного аппарата жгутика (FlhAB, FliOPQR и FliHIJ). В обоих случаях сигнальный пептид удаляется сигнальной пептидазой.
3. На третьем этапе путем самосборки в базальное тело встраиваются субъединицы статор- роторной системы (FliG, MotAB) и переключателя направления вращения (FHMN).
4. На четвертом этапе собирается ось, и здесь к экспорту впервые подключается секреторная система III. Сборка оси (субъединицы FHE, FlgBC, FlgF и FlgG) происходит от проксимального конца к дистальному. Вспомогательная субъединица FlgJ играет роль мурамидазы и проделывает в муреиновом саккулусе сквозное отверстие для растущей оси.
5. На пятом этапе собирается P-кольцо, субъединицы которого (Figl) экспортируются генеральной Sec-системой, а затем подвергаются дополнительному процессингу с помощью периплазматического шаперона FlgA и тиол:дисульфид-оксидоредуктазы Dsb-типа (см. раздел 8.4.2).
6. На шестом этапе собирается L-кольцо, субъединицы которого также экспортируются генеральной Sec-системой.
7. На седьмом этапе образуется незрелый крюк — из субъединиц FlgE и кэпа FlgD, которые экспортируются секреторной системой II, действующей и на последующих этапах.
8. На восьмом этапе образуется зрелый крюк, длину которого с точностью до 10% контролирует цитоплазматический белок FliK. В его отсутствии бактерии образуют гипертрофированные крюки, или «поликрюки» (англ polyhook).
9. На девятом этапе кэп отторгается, после чего на дистальном конце крюка из субъединиц FlgKL формируется соединительная зона.
10. На десятом этапе к крюку присоединяется кэп нити, состоящий из пентамера α βyδε субъединиц FHD.
11. На одиннадцатом этапе из субъединиц FHC собирается незрелая нить.
12. На последнем, двенадцатом этапе образуется зрелая нить, состоящая из 2 • 104 флагеллино- вых субъединиц. Длина ее строго не контролируется.
Сборка жгутика происходит за несколько секунд со скоростью 2 • 104 кДа • с-1, и лимитируется только темпом трансляции. Неясно, каким образом флагеллин проталкивается по каналу вращающейся нити — возможно, в этом участвуют центробежные силы.
Экспортный аппарат жгутика. Состоит из субъединиц FlhAB, FliOPQR, FliHIJ (рис. 62), выступает в цитоплазму, проходит через центральное отверстие ротора и частично погружен в MS-кольцо.
В состав экспортного аппарата входят интегральные, амфипатические и периферические субъединицы. Связь с MS-кольцом обеспечивает димер амфипатических субъединиц (75 кДа), каждая из которых состоит из восьми трансмембранных а- спиралей и периферических доменов, находящихся на цитоплазматической поверхности СМ. Димер амфипатических субъединиц FlhB (42 кДа), каждая из которых содержит четыре трансмембранных a-спирали, выполняет роль сигнальной пептидазы, а также контролирует прохождение субстратных белков через секреторную пору. Функция малых интегральных субъединиц FliOPQR (10-30 кДа), состоящих из 1-6 трансмембранных a-спиралей, не выяснена. В число периферических субъединиц входит АТФаза FHI и ее негативный регулятор FHH, а также молекулярный шаперон FliJ.
Как мы уже отмечали, субъединицы экспортного аппарата жгутика (в первую очередь, Flil) взаимно гомологичны субъединицам секреторной системы III, которая экспортирует факторы вирулентности Salmonella, Shilgella и Yersinia. По-видимому, жгутиковый секреторный аппарат был эволюционным предшественником секреторной системы III этих патогенов млекопитающих, а не наоборот (поскольку он имеется у более архаичных бактерий).
Интересно, что С-конец субъединицы FHI гомологичен каталитической β-субъединице F0F1-АТФазы.
Плавание с помощью экзоплазматических жгутиков. Если бактерия, имеющая жгутики, находится в жидкой изотропной среде, плавание не носит целесообразного характера. Клетка плывет в случайном направлении и на доли секунды «выключает» жгутиковый мотор, а затем вращение возобновляется в прежнем или в противоположном направлении.
Если среда анизотропна, движение приобретает адаптивный смысл. Оно контролируется сенсорными системами, влияющими на продолжительность заплывов и, соответственно, на частоту остановок.
Располагаются жгутики как по одному, так и пучками, при этом они могут находиться на полюсах, на боку или по всей поверхности клетки. Чаще всего клетка образует жгутики только одного типа, и они имеют постоянное расположение. Однако бывают исключения, например, Vibrio parahaemolyticus при плавании образует зачехленный полярный жгутик, работающий за счет энергии Smf. При переходе к роению вместо него появляются латеральные незачехленные жгутики, вращающиеся за счет Pmf (см. ниже).
У клетки с одним жгутиком, или монотриха (греч. monos — единственный и trichoma — волосы) при проплыве жгутик вращается по часовой стрелке или против часовой стрелки. При этом он соответственно толкает клетку вперед или тянет ее за собой (примером служит Pseudomonas aeruginosa). Иногда жгутик вращается только по часовой стрелке (примером служит Rhodobacter sphaeroides, у которой нить жгутика имеет редкую для бактерий форму правозакрученной спирали). В обоих случаях при выключении мотора клетка перестает двигаться самостоятельно и за счет броунова движения непредсказуемо изменяет направление своего движения.
У клетки со многими жгутиками, или политриха (греч. poly — множественный и trichoma — волосы) при заплыве жгутики объединяются в жгут, вращающийся против часовой стрелки. При вращении по часовой стрелке жгут расплетается. В результате независимого вращения отдельных жгутиков клетка начинает «кувыркаться» и непредсказуемо изменяет свою ориентацию.
Движение прокариотов в анизотропной среде всегда носит поисковый характер. Если сенсорные системы оценивают направление движения как физиологически более выгодное, то продолжительность заплыва увеличиваются. В частности, в случае хемотаксиса более выгодно оказаться в среде с более высокой концентрацией привлекающих веществ, или аттрактантов (лат. attracto — прикасаться). Поэтому клетка «растягивает удвольствие», продвигаясь вверх по градиенту аттрактанта, и реже кувыркается. Точно так же более выгодно оказаться в среде с меньшей концентрацией отпугивающих веществ, или репеллентов (лат. re-pello — отталкивать). Поэтому клетка «уходит от неприятности» и, продвигаясь вниз по градиенту репеллента, реже кувыркается. В свою очередь, клетка чаще кувыркается, удаляясь от области с более высокой концентрацией аттрактанта или приближаясь к области с более высокой концентрацией репеллента. «Заплывы» обязательно перемежаются с реориентацией путем кувыркания. В целом движение носит вероятностный характер и состоит из целесообразных заплывов, остановок и нецелесообразных заплывов.
Вероятность вращения жгутика против часовой стрелки повышается с помощью регулятора ответа (англ. response regulator) — цитоплазматического белка CheY, который входит в состав двухкомпонентной «фосфорелейной» сигнальной системы хемотаксиса (см. III том учебника). Ее вторым компонентом является цитоплазматическая сенсорная киназа CheA, которая получает внешний сигнал от хеморецепторов, расположенных в СМ (см. раздел 8.4.2). В фосфорилированном
состоянии белок CheY связывается с переключателем мотора FHM, входящим в состав базального тела.
В итоге, плывя зигзагами, клетка постепенно смещается в благоприятную для себя сторону. Такая стратегия движения называется «клинокинезом» (лат. clinatus — склоненный).
Роение. Плавание с помощью жгутиков можно рассматривать как типичный пример индивидуальной, или «авантюристической» подвижности.
В свою очередь, роение, или «плавание по твердой поверхности» — это типичный пример коллективной подвижности. В отличие от плавания роение происходит в двух координатах, и бактерии движутся только вперед или назад.
В природной среде роение жгутиконосных бактерий распространено очень широко. Оно является важным, но не единственным способом колонизации увлажненных твердых субстратов, поскольку с этой целью можно использовать и коллективное скольжение (см. раздел 8.6.3.3).
В лабораторных условиях наблюдать роение на быстро подсыхающем плотном 1-2%-ном агаре довольно трудно. Роящиеся клетки, или «швермеры» (нем. schwärmen — роиться) перемещаются только по мягкому агару или в присутствии поверхностно-активных веществ.
Наиболее подвижны клетки на периферии колонии, однако в целом они движутся со скоростью 2-10 мкм • с-1, что значительно меньше скорости плавания.
Роение связано с клеточной дифференциацией. Прежде всего, для него требуется больше жгутиков, поскольку швермерам приходится преодолевать силы поверхностного натяжения и продвигаться в слизи, которую сами же они и образуют. Кроме того, бактерия изменяет характер распределения жгутиков по поверхности своего тела, и/или вместо жгутиков одного типа образуются жгутики альтернативного типа. Примером первого варианта дифференциации является Proteus mirabilis, у которой роение было впервые описано еще в 1885 г. Эта бактерия плавает с помощью 6-10 равномерно распределенных по ее клеточной поверхности, или «перитрихальных» (греч. péri — вокруг и trichoma — волосы) жгутиков. На увлажненной твердой поверхности клетка сильно удлиняется и гиперфлагеллируется, хотя строение жгутиков остается прежним. Примером второго варианта дифференциации является Vibrio parahaemolyticus. Эта бактерия плавает при помощи единственного зачехленного жгутика типа Fla (от англ. flagella). На увлажненном твердом субстрате она образует перитрихальные незачехленные жгутики типа Laf (от англ. lateral flagella).
Важную роль в роении играет внутрипопуляционная коммуникация, или «чувство кворума» (англ. quorum sensing).
Чувство кворума — это способ химической коммуникации между бактериями. Оно связано с биосинтезом растворимых сигнальных молекул — автоиндукторов, или феромонов (греч. pherein — переносить и hormon — возбуждать). В качестве феромонов грамотрицательные бактерии используют главным образом лактоны N-ацилгомосерина (англ. acylhomoserine lactone, AHL), а грамположительные бактерии — специфические пептиды, подвергающиеся посттрансляционной модификации.
С помощью поверхностных рецепторов бактерия оценивает наружную концентрацию феромонов, которые она же и выделяет, что позволяет косвенно судить о локальной плотности собственной популяции. После того, как бактерии получают сигнал о наличии кворума, они приступают к адаптивным действиям. Обычно это требует больших метаболических затрат, которые поодиночке нецелесообразны. В определенной степени это напоминает структурно-функциональную стратегию многоклеточного организма.
Примерами коллективных действий, основанных на чувстве кворума, являются биолюминесценция, биосинтез антибиотиков, приобретение состояния компетентности для генетической трансформации, дифференциация эндоспор, конъюгативный перенос плазмид, образование и секреция факторов патогенности, а также роение с целью коллективной секреции экзоферментов или для образования плодовых тел.
При роении биосинтез жгутикового аппарата контролируется тем же регуляторным каскадом, что и при плавании. Поскольку центральная роль принадлежит мастер-оперону flhDC,экспрессия его генов под действием позитивных регуляторов возрастает на порядок, что и определяет дифференциацию швермеров у таких роящихся бактерий, как Р. mirabilis.
Направление движения при роении контролируется фосфорелейной системой хемотаксиса, хотя сама она в дифференциации швермеров не участвует.
Плавание с помощью периплазматических жгутиков. Данный тип подвижности уникален для спирохет (филы BXVII Spirochaetes). Большинство спирохет имеет форму спирализованного цилиндра, что не вполне соответствует названию типового рода (лат. spira — спираль и греч. chaite — грива; «бактерия со спиральным гребнем»). В редких случаях спирохеты действительно образуют спиральный гребень и даже принимают форму уплощенной синусоиды.
Хотя спирохеты обладают грамотрицательным морфотипом, базальное тело жгутика заякорено в СМ, кольца C/MS/P/L отсутствуют, а спиральная нить не покидает пределов периплазматического компартмента. Жгутики расположены субтерминально, поодиночке или пучками, на одном или двух концах «протоплазматического цилиндра» (англ. protoplasmiccell cylinder). Они направлены к противоположному полюсу клетки и либо доходят до него, либо заканчиваются до или после экватора. В том случае, когда одиночные встречные жгутики или пучки до 100 жгутиков перекрываются, они образуют «аксиальную нить» (англ axial filament). Поскольку спирохеты имеют гибкую оболочку, протоплазматический цилиндр вместе с муреиновым саккулусом и ОМ повторяют жесткую суперспиральную форму жгутиков, которые, таким образом, играют роль экзоскелета (см. раздел 8.3.9.1).
С помощью периплазматических жгутиков спирохеты лучше плавают в более вязкой среде, где их скорость достигает 20 мкм • с-1: «Штопор ввинчивается в пробку, а не в воду». Поскольку многие спирохеты являются симбионтами или патогенами животных, особенность их плавания способствует инвазии и диссеминации. Кроме того, поскольку периплазматические жгутики не выходят на наружную поверхность клетки, спирохеты защищены от антижгутиковых антигенов.
Периплазматические жгутики спирохет отличаются от экзоплазматических жгутиков других бактерий. В частности, они имеют более толстую нить (~25 нм), которая состоит из флагеллинов (FlaB1-3), распределенных в разных участках нити, и белка чехла (FlaA). Как у других бактерий, жгутик спирохет имеет крюк и базальное тело. Структура их еще не выяснена.
По аналогии с другими бактериями, флагеллин спирохет экспортируется с помощью секреторной системы III. Препротеин чехла к суперсемейству флагеллинов не относится. Он содержит сигнальный пептид и транспортируется при помощи генеральной Sec-системы. Предполагается, что за счет чехла нить стабилизируется в левозакрученной конфигурации.
Подвижность спирохет имеет адаптивное значение. Она контролируется системой хемотаксиса и, видимо, вискозитаксиса (когда внешним сигналом служит градиент вязкости). Однако сам механизм плавания спирохет более сложен, чем плавание с помощью экзоплазматических жгутиков. У них существуют четыре основных варианта подвижности:
— заплыв «вперед», жгутики вращаются в противоположных направлениях (против часовой стрелки на переднем и по часовой стрелке на заднем конце клетки);
— заплыв «назад», жгутики вращаются в противоположных направлениях (по часовой стрелке на переднем и против часовой стрелки на заднем конце клетки);
— остановка «первого типа», жгутики вращаются в одном направлении (против часовой стрелки на переднем и заднем концах клетки);
— остановка «второго типа», жгутики вращаются в одном направлении (по часовой стрелке на переднем и заднем концах клетки).
Таким образом, направление движения изменяется, если оба пучка жгутиков (или два антиподных жгутика) одновременно изменяют направление вращения. Если оно изменяется только на одном конце клетки, происходит остановка.
Две «остановочные» формы морфологически не различаются, хотя иногда клетка изменяет свою форму и перегибается посередине. У представителей рода Leptospira при вращении жгутиков против часовой стрелки концы клетки имеют спиралевидную форму, а при вращении по часовой стрелке загибаются крючком.
Такие же четыре варианта подвижности существуют у представителей рода Spirillum (класс «Betaproteobacteria») и других спирилл, однако эти бактерии образуют антиподные пучки экзоплазматических жгутиков, а их жесткий саккулус при движении не деформируется.
8.6.2.3. Жгутики архей
Подобно бактериям, большинство архей образует жгутики. Они имеются у представителей всех основных цитофизиологических групп домена Фксрфуф — экстремальных галофилов, метаногенов, термоплазм и серазависимых гипертермофилов. Широкое распространение жгутиков у архей-экстремофилов свидетельствует об исключительно высокой стабильности этих структур.
Жгутик архей — аналог бактериального жгутика. Хотя внешне жгутики архей напоминают бактериальные и так же вращаются, они не гомологичны им и отличаются по способу сборки и устройству мотора. Нить жгутика архей представляет собой не левозакрученную, а правозакрученную спираль. У перитрихальных архей жгут, собранный из отдельных жгутиков, вращается по часовой стрелке и толкает клетку вперед. При изменении направления вращения он не расплетается и тянет клетку за собой. Плавают архей относительно медленно — их скорость составляет <10% бактериальной (2-3 мкм • с-1), роение у них не описано.
В геномах архей не обнаружены гомологи генов, кодирующих не только флагел- лины бактериального типа, но и субъединицы оси, субъединицы крюка со вспомогательными белками, а также субъединицы колец, статорных белков и переключателей направления вращения. В то же время ряд вспомогательных «жгутиковых» генов архей (flаС-J) у бактерий не встречается.
В отличие от субъединиц бактериального жгутика, которые экспортируются двумя секреторными системами, транспорт субъединиц археотного жгутика происходит только с помощью генеральной Sес-системы. Способ крепления жгутика в теле археотной клетки неизвестен, базальное тело не выявлено, а строение и механизм вращения мотора не установлены (см. ниже).
Нить жгутика и археотные флагеллины. Нить жгутика архей тоньше, чем у бактерий (10-15 нм против ~20 нм). Под электронным микроскопом она выглядит как квази-кристаллическая структура из спирально упакованных субъединиц и продольно уложенных протофиламентов. Центральный канал в нити археотного жгутика не выявлен. Возможно, он вообще отсутствует.
В отличие от бактерий, обычно имеющих единственный флагеллин, жгутики архей в большинстве случаев построены из 2-5 флагеллинов молекулярной массой 20-100 кДа. Они расположены в разных частях нити (исключением служит единственный флагеллин Т. volcanium). Флагеллины архей входят в суперсемейство взаимно гомологичных белков, не проявляющих гомологии с членами суперсемейства бактериальных флагеллинов. Кроме того, они синтезируются в виде препротеина с сигнальным пептидом, который удаляется при секреции. Наконец, флагеллины архей обычно гликозилированы, что у бактерий встречается довольно редко.
Крюк. В отличие от бактериального крюка, размер которого составляет ~50 нм, у архей гибкое сочленение жгутика сильно варьирует по длине (70-300 нм).
«Базальное тело». Архей лишены ОМ, и ригидные слои клеточной стенки у них не гомологичны бактериальным. Структуры, напоминающие кольца, равно как и другие, заякоривающие жгутик в клеточной оболочке, у них не обнаружены. Поскольку в большинстве случаев ригидные слои у архей тоньше и механически «слабее» бактериальных или вообще отсутствуют (примером служат представители р. Thermoplasma), роль базального тела возлагают на себя интрацитоплазматические структуры — полярные мембраноподобные органеллы (см. раздел 8.3.11.3).
Регуляция биосинтеза и сценарий сборки археотного жгутика. Так же, как у бактерий, жгутики архей имеют адаптивное значение, и их биогенез контролируется внешней средой. Например, у Methanospirillum hungatei жгутики образуются только в зоне широкого температурного оптимума (30-37°С), но не экспрессируются при 25 или 40°С. У другого метаногена, M. jannashii, биосинтез флагеллинов FlaB2, FlaB3, FlaD и FlaE стимулируется при снижении парциального давления водорода. Архей имеют «бактериальный» набор генов регуляторного каскада хемотаксиса (cheAB, cheY и cheRW), и их движение осуществляется по принципу клинокинеза.
Флагеллины архей, по-видимому, экспортируются с помощью генеральной Sec- системы (SecDEFY). Префлагеллин подвергается посттрансляционному процессингу под действием сигнальной пептидазы. Гликозилирование археотного флагеллина осуществляется не в цитоплазме, а на наружной поверхности СМ.
Предполагается, что сборка нити археотного жгутика происходит в ее основании, что напоминает механизм сборки нити бактериальной фимбрии, но приципиально отличается от механизма сборки нити бактериального жгутика.
Сходство жгутиков архей с фимбриями IV типа. Установлено, что флагеллин гомологичен пилину — субъединице фимбрии IV типа. В обоих случаях препротеины служат субстратами для сигнальной пептидазы. Один из жгутиковых белков архей (Flal) гомологичен консервативной субъединице фимбрий IV типа Neisseria gonorrhoeae и Р. aeruginosa —АТФазе PilT. Согласно общепринятой модели, фимбрии IV типа втягиваются внутрь клетки, на чем основано движение рывком, или «твитчинг» (см. ниже). Предполагается, что фимбрии IV типа способны вращаться. Таким образом, структурное сходство жгутиков архей с фимбриями IV типа может означать, что они обладают одинаковыми кинематическими свойствами.
8.6.3. Фимбрии
Наряду со жгутиками, примером специализированного компартмента со смешанной локализацией служат фимбрии IV типа.
В настоящем разделе мы рассмотрим не только этот, но и другие типы фимбрий, а также затронем более широкую тему, касающуюся адгезинов.
В отличие от жгутиков, которые в первую очередь выполняют роль локомоторных органелл и в основном используются для плавания в жидкой среде, фимбрии обеспечивают контактные взаимодействия трех типов:
— между гомологичными или гетерологичными бактериями;
— между бактериями и другими живыми организмами;
— между бактериями и (не)органическими субстратами.
Такие контакты определяются различными нековалентными взаимодействиями (ионными, водородными, гидрофобными и силами ван дер Ваальса). В общем случае они называются адгезией и осуществляются с помощью адгезинов.
Адгезины. Для поверхностной адгезии, т. е. для прикрепления к органическим или неорганическим телам прокариоты могут использовать разнообразные адгезины.
В самой общей форме адгезины прокариотов можно определить, как факторы иммобилизации клеток. Они позволяют установить относительно долговременные анизотропные взаимодействия, которые приносят биологическую пользу отдельной клетке, популяции, а также локальному экоценозу.
С помощью адгезинов прокариоты, использующие неорганические субстраты в качестве донора или акцептора электронов, прикрепляются к твердым минеральным частицам.
Взаимная адгезия позволяет прокариотам, особенно бактериям образовывать микроколонии, пленки на поверхности жидкой среды, а также биопленки на твердых субстратах.
Так как субстраты и бактерии чаще всего одноименно (отрицательно) заряжены, выгодно осуществлять дистанционную адгезию и размещать адгезины в дистальной части протяженной придаточной структуры — жгутика, фимбрии или шипа.
Адгезины играют особо важную роль у симбиотических бактерий (например, у цианобактерий родов Anabaena и Nostoc, вступающих в эктосимбиоз с высшими растениями).
Для патогенных бактерий адгезины служат одним из факторов, позволяющих им колонизировать макроорганизм. При помощи адгезинов патогенные бактерии распознают поверхностные рецепторы на хозяйских клетках. Специфика этого взаимодействия определяет таргетинг бактерии по отношению к определенному хозяину, а также тому или иному типу хозяйских тканей. Кроме того, адгезины позволяют бактерии зафиксироваться на поверхности ткани в начальной фазе инфекции, что очень важно в проточной среде (например, благодаря адгезинам энтеропатогенные бактерии не удаляются из кишечника в результате перистальтики).
Среди адгезинов, находящихся в «эпицентре» контактного взаимодействия бактерий с эукариотными клетками, выделяют сборную группу лектинов (лат. lectio — собирание в группу). Лектины содержат эпитопы, распознающие олигосахаридные цепи гликопротеинов, которые расположены на поверхности животной или растительной клетки.
Функции адгезинов могут выполнять:
— индивидуальные поверхностные белки;
— поверхностные белковые кластеры;
— придаточные структуры.
Адгезины первой и второй групп называются «нефимбриевыми» адгезинами (англ. non- fimbrial adhesins, NFA).
Примерами мономолекулярных адгезинов являются липопротеины стрептококков (см. раздел 8.5.1.3) или спирохет (см. 8.4.1.2).
Примерами адгезинов, образующих поверхностные белковые кластеры, являются инвазии и интимин. Первый из них образует Y. paratuberculosis; второй имеется у энтеропатогенной бактерии Е. coli штамм ЕРС. В обоих случаях на поверхность клетки выходит пучок столбчатых доменов адгезина, дистальные концы которых выполняют роль лектина.
Важнейшими адгезинами третьей группы являются филаментозные придаточные структуры — фимбрии.
8.6.3.1. Фимбрии бактерий
Бактериальные фимбрии (лат. fimbria — нитка), или «пилусы» (лат. pilus — волос) представляют собой длинные и тонкие нитевидные придаточные структуры, состоящие из белковых субъединиц.
Способностью образовывать фимбрии обладают многие патогенные и непатогенные грамотрицательные бактерии. Значительно реже фимбрии встречаются у грамположительных бактерий. У архей аналогичные структуры образуются только в исключительных случаях.
Фимбрии состоят из субъединиц (протомеров) белка пилина и представляют собой прямые и жесткие, реже, гибкие полые цилиндры длиной до 2 мкм и диаметром 3-10 нм. Фимбрии различаются по размеру, морфологии и локализации, а также по числу копий на клетку (1-500).
Классификация фимбрий. Современная классификация фимбрий непоследовательна и запутана, и поэтому мы приводим ее в упрощенном виде. Ее исходный вариант был разработан в начале 1970-х годов применительно к энтеропатогенным и уропатогенным представителям сем. Enterobacteriaceae, в частности Е. coli. Фимбрии подразделяют на четыре типа, прежде всего в соответствии с их антигенными свойствами. Помимо этого, учитываются морфология и ультраструктура фимбрий, а также молекулярный состав их субъединиц.
К I типу относятся «маннозо-чувствительные» фимбрии (вызываемая ими агглютинация эритроцитов курицы или морской свинки ингибируется маннозой или гликозидами со сходной структурой).
Ко II типу относятся «маннозо-нечувствительные» фимбрии, а также фимбрии, не вызывающие гемагглютинацию.
К III типу относятся фимбрии, которые агглютинируют эритроциты человека, обработанные танниновой кислотой.
К IV типу относятся так называемые «половые» фимбрии, участвующие в бактериальной конъюгации.
Однако такая классификация не универсальна хотя бы потому, что гемагглютинацию вызывают не все варианты фимбрий.
Более конструктивна классификация фимбрий на два типа — главный и альтернативный. Будучи результатом конвергентной эволюции, эти фимбрии имеют разную локализацию и разные механизмы биогенеза (рис. 63).
Рис. 63. Строение бактериальных фимбрий. А — фимбрия главного типа; Б — фимбрия альтернативного типа; СМ — цитоплазматическая мембрана; ОМ — наружная мембрана; М — муреиновый саккулус.

Фимбрии главного типа. Эти фимбрии относятся к типам I — III по традиционной классификации. Они имеют жесткую структуру, образуются в большом числе и расположены перитрихально. У них нет базального тела, и поэтому нить непосредственно прикрепляется к наружной поверхности клетки (рис. 63, А). Субъединицы нити транслоцируются через СМ при помощи генеральной Sec-системы. Поскольку транслоцированный пилин находится в развернутой конформации, периплазматические молекулярные шапероны, в частности FimC, предохраняют его от преждевременной агрегации. Комплекс пилин/шаперон презентируется «белку-привратнику» (англ. usher), который входит в состав ОМ. Привратник FimDобразует проводящий канал и пропускает через ОМ отдельные субъединицы фимбрии. Однако он слишком узок для мультимерной нити, которая полимеризуется только в экзоплазматическом компартменте. Возможно, что белок-привратник не только образует пору,
но и выполняет активную роль в транслокации пилина, действуя в качестве флиппазы (см. раздел 8.4.1.1). Кроме того, привратник служит докинг-площадкой для полимеризации нити в основании и обеспечивает ее прикрепление к ОМ.
Фимбрии главного типа выполняют только адгезионные функции, и поэтому их иногда называют фимбриями «общего назначения», или «адгезионными» фимбриями.
Фимбрии альтернативного типа. Эти фимбрии относятся к IV типу по традиционной классификации. Они могут иметь жесткую или гибкую структуру, образуются в количестве 1-2 на клетку и расположены на одном из ее полюсов. Для них характерно присутствие базального тела, субъединицы которого гомологичны субъединицам базального тела жгутика (рис. 63, Б). Основание нити упирается в СМ, и она проходит через секретиновую пору, расположенную в ОМ (см. раздел 8.6.1). Субъединицы пилина транслоцируются с помощью секреторной системы III через канал, проходящий внутри базального тела. Затем нить полимеризуется путем встраивания новых субъединиц в ее основание.
Фимбрии альтернативного типа обладают не только адгезионными свойствами. Они способны быстро «сокращаться» благодаря разборке основания на отдельные
субъединицы, которые образуют особый пул в СМ. Однако дистальный конец нити по-прежнему прикреплен к организму-партнеру или к субстрату. Поэтому клетка подтягивается к ним, и между ними устанавливается тесный контакт. Благодаря такому свойству фимбрии альтернативного типа участвуют в твитчинге, или подвижности «рывком», а также в образовании партнерских пар при конъюгации (см. ниже).
8.6.3.2. Фимбрии главного типа
В состав этих фимбрий входят главные субъединицы, минорные субъединицы, регуляторные белки, периплазматический молекулярный шаперон и привратник. Они кодируются хромосомными или плазмидными оперонами.
Периплазматический молекулярный шаперон молекулярной массой ~25 кДа взаимодействует с субъединицами фимбрии в соотношении 1:1. Он содержит два бочкообразных домена, каждый из семи антипараллельных ß-цепей. Белок-привратник (82 кДа) состоит из 24 антипараллельных интегральных ß-цепей и двух протяженных цитоплазматических доменов — N-концевого и С-концевого.
Фимбрии главного типа исключительно устойчивы — на них практически не действуют такие реагенты, как додецилсульфат или мочевина, хотя при обработке насыщенным гуанидином или горячей кислотой они диссоциируют на отдельные субъединицы.
В популяции энтеробактерий биогенез фимбрий главного типа периодически то прекращается, то возобновляется. Такое явление называется «фазовой» изменчивостью. Переключение из состояния «фимбрия образуется» (англ. phase on) в состояние «фимбрия не образуется» (англ. phase off) происходит в результате хромосомной инверсии или на уровне инициации транскрипции. Частота переключений регулируется условиями окружающей среды (значением температуры, присутствием определенного источника углерода и т. д.).
Фазовая изменчивость в случае фимбрий носит адаптивный характер. Они полезны для адгезии к эпителиальным клеткам, однако их биогенез является «дорогостоящим» процессом. Кроме того, прикрепившаяся бактерия более уязвима к действию иммунных систем макроорганизма. Поскольку бактерия, с одной стороны, не может прогнозировать успешность инвазии, но, с другой стороны, должна к этому подготовиться, она «на всякий случай» диверсифицирует свою популяцию, одновременно образуя клетки с фимбриями и без них.
Наиболее известными представителями фимбрий главного типа являются Fim- и Рар-фимбрии.
Fim-фимбрии. Такие фимбрии («маннозо-чувствительный» I тип по традиционной классификации) образуются примерно у 80% штаммов бактерий из сем. Enterobacteriaceae, в том числе у энтеротоксигенных штаммов Е. coli.
Нить Fim-фимбрии представляет собой полый цилиндр длиной до 2-10 мкм и диаметром 5-10 нм. Ширина внутреннего отверстия составляет 1,5 нм. Стенки цилиндра образованы 103протомерами пилина FimA (17 кДа), которые укладываются в плотную правозакрученную спираль, на один оборот которой приходятся ~3,2 протомера.
Гены, кодирующие Fim-фимбрию, объединены в хромосомный fim-оперон. В число продуктов его экспрессии, помимо пилина FimA и лектина FimH входят белки FimC(периплазматический шаперон), FimD (привратник), FimFG (регуляторы роста нити в длину) и FimEB (регуляторы фазовой изменчивости).
Субъединицы лектина FimH (29 кДа) содержат пилиновый домен, с помощью которого они встраиваются в нить фимбрии, и N-концевой лектиновый домен, содержащий «D-маннозный» эпитоп. Между ними расположен короткий интердоменный линкер. Лектин FimH обеспечивает презентацию фимбрии гликопротеинам хозяйской клетки, которые содержат N-замещенные олигоманнозные цепи.
Рар-фимбрии. Эти фимбрии («маннозо-нечувствительный» II тип по традиционной классификации) образуют клетки уропатогенных штаммов энтеробактерий.
Гены, кодирующие Рар-фимбрию (от англ. pyelonephritis), объединены в хромосомный рар-оперон. В число продуктов его экспрессии, наряду с пилином РарА (20 кДа), лектином PapG, периплазматическим шапероном PapD и привратником РарС, входят регуляторы фазовой изменчивости PapIB. Субъединица РарН, расположенная между привратником и основанием нити, регулирует рост нити в длину (аналогичную роль играет субъединица FimF в составе Fim-фимбрии).
Лектин PapG обеспечивает презентацию фимбрии гликолипидным рецепторам эритроцитов человека, содержащим «не маннозный» эпитоп из двух остатков галактозы, соединенных связью α1 —> 4. Лектин PapG расположен не в дистальной части нити диаметром 7 нм, как у Fim-фимбрии, а на конце ее филаментозного придатка диаметром 2 нм, состоящего из цепочки субъединиц белка РарЕ.
Как уже указывалось, у грамположительных бактерий фимбрии встречаются реже, чем у грамотрицательных бактерий.
В качестве примеров можно назвать гемагглютинирующие маннозо-нечувствительные фимбрии Acinomyces naeslundii и Corynebacterium diphtheriae (фила BXIV Actinobacteria). В обоих случаях пилин имеет молекулярную массу ~60 кДа, что втрое превышает молекулярную масу пилина грамотрицательных бактерий.
8.6.3.3. Фимбрии альтернативного типа
Как мы уже отмечали, фимбрии альтернативного типа принципиально отличаются от фимбрий главного типа по морфологии, числу копий, локализации, механизму биогенеза и выполняемым функциям.
Как любая фимбрия, они обладают адгезионными свойствами. Однако, в отличие от фимбрий главного типа, их функции и шире, и специфичнее «простой» адгезии. В частности, они могут быть связаны:
— с клеточной подвижностью;
— с образованием биопленок;
— с секрецией факторов вирулентности и токсинов;
— с транспортом ДНК входе «горизонтального» переноса генов (при конъюгации или при генетической трансформации) или с восприятием от бактериофага инфекционной ДНК.
Фимбрии IV типа и подвижность с их помощью. При помощи фимбрий IV типа осуществляется особый тип подвижности. Он известен под двумя названиями - «движение благодаря втягиванию фимбрий» (англ. retractile mobility) и «рывок», или «твитчинг» (англ. twitching).
Твитчинг был открыт в начале 1970-х годов у «оппортунистического» патогена Acinetobacter calcoaceticus — представителя нормальной микробиоты кожи человека, который проявляет патогенные свойства при ослаблении иммунитета и/или на фоне интенсивного лечения антибиотиками. Позднее этот тип подвижности обнаружили у «госпитального» патогена (англ. nosocomial pathogen) Pseudomonas aeruginosa, а также у патогенных штаммов Е. coli, Neisseria gonorrhoeae и многих других грамотрицательных бактерий. Тогда же твитчинг определили, как скачкообразное движение отдельной клетки по увлажненной поверхности твердого субстрата со скоростью в несколько десятых долей микрометра в секунду.
Вскоре выяснилось, что в результате суммирования твитчинга у индивидуальных клеток бактериальные колонии могут распространяться в радиальном направлении со скоростью до 0,3 мкм • с-1 (примером служит Myxococcus xanthus). Это приводит к образованию визуально наблюдаемых зон дисперсии, которые имеют форму концентрических кругов или протуберанцев. В обоих случаях бактерии располагаются параллельно друг другу. Перед смещением соседние клетки сначала выстраиваются «в затылок» друг другу, а затем резко принимают продольное положение. Твитчинг по своей природе социален, хотя при контакте с твердой поверхностью субстрата его могут осуществлять и отдельные клетки.
Таким образом, твитчинг является одним из механизмов коллективного скольжения. Благодаря ему происходит распространение колоний по поверхности богатой среды и образование биопленок. С помощью твитчинга М. xanthus образует на бедной среде плодовые тела.
Твитчинг вызывается втягиванием фимбрий IV типа, которые имеют диаметр 6 нм, достигают в длину 5 мкм и расположены поодиночке либо группами. У палочковидных бактерий они находятся на одном или обоих полюсах клетки.
Биогенез фимбрий IV типа у М. xanthus и Р. aeruginosa контролируется соответственно системами хемотаксиса frz и chp. За этот метаболический процесс, а также за сам твитчинг отвечают не менее 35 генов. В частности, ген pilA кодирует субъединицы фимбрии, а ген pilD их процессинг. Субъединица РПВ и АТФаза PilT, расположенные на внутренней поверхности СМ, обеспечивают сборку фимбрии и контролируют ее длину, а белок PilQ выполняет роль секретина. Как уже отмечалось, аппарат сборки фимбрии IV типа содержит элементы, гомологичные элементам секреторной системы III. Выяснено, что минорные белки, или «псевдопилины» (англ. pseudopilin) участвуют в сборке нити и функционировании фимбрии. Их конкретные функции пока еще не установлены.
Втягивание фимбрии IV типа происходит со скоростью ~1 мкм • с-1, и создается физическое усилие ~10-10 Н. Этот процесс осуществляется спонтанно, без затраты энергии. Нижний конец фимбрии как бы «расплавляется», и субъединицы пилина вливаются в пул, расположенный в СМ. Из-за уменьшения длины фимбрии клетка резко подтягивается к субстрату. Это и есть собственно твитчинг.
Фимбриям IV типа принадлежит важная роль в образовании биопленок (см. раздел 7.3.5).
Нгр-фимбрии. Фимбрии IV типа участвуют не только в подвижности, но и в секреции факторов вирулентности и токсинов.
Примером служат Нгр-фимбрии (от англ. hypersensitive response and pathogenicity). Кодирующий их оперон является одним из представителей высококонсервативных hrс-оперонов (от англ. hypersensitive response conserved). Продукты экспрессии hrс-оперонов вызывают стресс у инфицированного макроорганизма, а также оказывают патологическое воздействие на его клеточные структуры и физиологические процессы. Такие опероны входят в состав «островков патогенности» у зоопатогенных или фитопатогенных бактерий.
В инфраструктурном плане геном бактерий состоит из двух неравных частей. Основу генома составляет ядро, или кор (англ. core) в виде ансамбля генов, детерминирующих важнейшие структурно-функциональные свойства клетки; эти гены сходны по содержанию ГЦ-пар, набору используемых кодонов и слабо мутируют. Меньшая часть генома (~15%) —это «геномные островки» (англ. genome island), или участки генома, отличающиеся от кора по содержанию ГЦ-пар, а также набору используемых кодонов.
Геномные островки происходят от интегрированных в геном мобильных генетических элементов (плазмид, транспозонов и бактериофагов). Они содержат гены для специализированного метаболизма, устойчивости к антибиотикам и гены для адаптации к абиотическим факторам среды, а также гены для симбиоза и вирулентости. В последнем случае говорят об «островках патогенности» (англ. pathogenicity island).
Размер геномных островков 10-200 т. п. н. Они устроены по общему принципу и включают в себя прямые повторы, мобильные элементы, структурные и регуляторные гены. Концы геномного островка, как правило, фланкированы прямыми повторами (англ. direct repeat), приобретенными в результате интеграции в хромосому мобильного генетического элемента. Основу геномного островка составляют структурные гены. Как правило, речь идет о полном наборе генов, необходимом для реализации какого-либо признака, что делает возможным и целесообразным его «горизонтальный» перенос.
Примерами /irc-оперонов служат уор-оперой чумной бактерии Yersinia pestis, а также /ггр-оперон фитопатогенов Agrobacterium tumefaciens, Erwinia amylowora, Pseudomonas syringae, Ralstonia solanacearum и Xanthomonas campestris.
Нгр-фимбрии достигают в длину 2 мкм, что позволяет бактерии пробуравить клеточную стенку хозяина и доставить в его цитоплазму разные факторы патогенности. Среди них можно назвать оказывающие регуляторный эффект HrpZ-бел- ки, или «гарпины» (англ, harpin, от аббревиатуры Hrр) и AvrB-белки (от англ. Agrobacterium-virulence), а также токсины, которые ингибируют хозяйские ферменты, например, глутаминсинтетазу.
Пилин Hгр А имеет небольшой размер (6-11 кДа). Его структура очень вариабельна; взаимная гомология этого белка у разных бактерий не превышает 30%. Биогенез Нгр-фимбрии тесно связан с работой секреторной системы III - базальная часть насаживается на иглу, а затем наращивается за счет протомеров, выходящих через дистальный конец нити. Такой способ сборки характерен для бактериальных жгутиков, но для фимбрий он уникален.
Половые фимбрии. Еще одной, очень важной функцией фимбрий IV типа является контактная передача ДНК от одной бактерии к другой.
Этот процесс называется конъюгацией, и в нем участвуют «половые» фимбрии (англ. sex pilus). В отличие от Нгр-фимбрий, они не используются для секреции белка, а участвуют только в конъюгации или крайне редко становятся каналом для выхода во внешнюю среду трансформирующей ДНК (примером служит N. gonorrhoeae).
Среди половых фимбрий наиболее известны F-фимбрии Е. coli, которые закодированы в конъюгативных F-плазмидах. Клетки, несущие F-фимбрии (F+), называются донорными, или «мужскими», а клетки без F-фимбрий (F-) акцепторными, или «женскими». Донор-акцепторная пара участвует в процессе конъюгации — наиболее распространенном у бактерий способе горизонтального переноса генетической информации.
Конъюгация представляет собой контактный процесс, который индуцируется физико-химическими факторами внешней среды или определяется межклеточным сигналингом на определенной фазе развития популяции. Конъюгация способствует генетической рекомбинации и естественному отбору, в частности она позволяет распространить в популяции патогенных бактерий гены
устойчивости к антибиотикам. Конъюгация возможна у близкородственных или эволюционно дистанцированных бактерий и иногда происходит даже на интердоменном уровне (см. ниже).
В состав аппарата для конъюгации у грамотрицательных бактерий входят:
— половая фимбрия, обеспечивающая взаимную адгезию партнеров;
— гомолог секреторной системы III, образующий конъюгативную пору, или проводящий канал в клеточной оболочке донора, по которому транспортируется одноцепочечная Т-ДНК (от англ. transfer — перенос).
Конъюгация у грамположительных бактерий еще слабо изучена. Она происходит без участия половых фимбрий.
Роль F-фимбрии при конъюгации заключается в том, чтобы распознать рецептор на женской клетке и прикрепиться к ней, а затем сократиться. Благодаря этому устанавливается тесный контакт между оболочками клеток-партнеров (англ. conjugational junction).
Информация о контакте (англ, mating signal) поступает на «релаксосому» — комплекс F-плаз- миды с белком IHF (от англ, integration host factor), релаксазой TrwA и геликазой TrwC. Одноцепочечная Т-ДНК с помощью гетерогексамера белка TrwB передается периферической АТФазе NBS (от англ. nucleotide binding site). Последняя играет роль помпы и проталкивает Т-ДНК через пору в клеточной оболочке, или «транслокон».
Существуют два варианта конъюгативных фимбрий. Первый из них представлен длинными и гибкими F-фимбриями, а второй — короткими и жесткими Р-фимбриями.
F-фимбрия имеет длину 2-20 мкм, ее диаметр составляет ~8 нм, а ширина внутреннего канала — 2 нм. Стенки F-фимбрии образованы плотно уложенным F-пили- ном, по пять субъединиц на один оборот спирали. Нить F-фимбрии служит рецептором для нитчатых «мужских» бактериофагов, которые адсорбируются по всей ее длине (фаги f2, MS2, R17 и Q(3) или только на ее конце (фаги f1, fd и М13).
За образование F-пилина отвечают три гена. Структурный ген traA кодирует «препилин» молекулярной массой ~13 кДа. Продукт гена traQ участвует в процессинге препилина с помощью сигнальной пептидазы LepB, которая расположена на внутренней поверхности СМ и отрезает от него макропептид, состоящий из 51 аминокислоты. Продукт гена traX путем N-ацетилирования образует из оставшегося фрагмента препилина зрелый пилин (7 кДа).
Хотя F-фимбрия состоит только из одного типа субъединиц, в их секреции и сборке задействованы не менее 17 генов, кодирующих структурные и регуляторные белки (рис. 63, Б).
Субъединица ТгаК образует секретиновую пору в ОМ, через которую фимбрия «прорастает» на поверхность клетки. С помощью субъединиц TraV секретиновое кольцо прочнее фиксируется в ОМ. Субъединицы ТгаВ прикрепляют секретиновое кольцо к СМ и совместно со вспомогательным белком TraD образуют в оболочке сквозную пору диаметром 22Â. Субъединицы TraEL участвуют в сборке фимбрии с помощью «сопрягающей» АТФазы ТгаС и регулируют рост фимбрии в длину, а субъединицы TraF выполняют роль периплазматических шаперонов.
Вспомогательные субъединцы TraGN стабилизируют взаимодействие половых партнеров, а субъединицы TraHUW и ТгbС принимают участие в образовании поры и «сокращении» фимбрии.
После адгезии F-фимбрии к поверхностым рецепторам женской клетки (роль которых, в частности, играют порины) или после адсорбции «мужского» бактериофага на F-фимбрии она «сокращается» путем разборки на проксимальном конце, причем это происходит спонтанно, без затраты энергии. В результате этого женская клетка или адсорбированный бактериофаг доставляются к поверхности мужской клетки, и между ними векторно переносится ДНК.
Т-фимбрии. Особым примером конъюгативных фимбрий служит Т-фимбрия.
В данном случае ДНК передается не от бактерии к бактерии, а от бактерии к растительной клетке. В результате такого «интердоменного» переноса Т-ДНК интегрируется в эукариотный геном.
Т-фимбрия кодируется mr-опероном, входящим в состав Ti-плазмиды (от англ. tumor inducing) фитопатогенной бактерии Agrobacterium tumefaciens, которая вызывает неопластический рост стеблей — «коронарные галлы» (англ, crown gall disease). Результатом интеграции Т-ДНК в растительную хромосому является не только аномальный рост хозяина, но и индуцированный синтез специфических азотсодержащих субстратов, которые поддерживают рост паразитических бактерий в ткани коронарного галла — опинов (англ. opin). Название «опин» происходит от октопина — карбоиминного соединения, состоящего из пирувата и аргинина, которое было впервые обнаружено у осьминога из рода Octopus.
В состав Ti-плазмиды входят генные локусы для биосинтеза и катаболизма опинов, генные локусы для образования фитогормонов (ауксинов и цитокининов), а также генный локус для конъюгативного переноса virD и генный локус для вирулентности virB. Локус vir В состоит из 11 генов, которые кодируют особый вариант конъюгативной системы — транс локон и Т-фимбрию (по аналогии с аппаратом межбактериальной» конъюгации).
Конъюгативная Т-фимбрия относится к группе коротких и жестких Р-фимбрий. Ее субъединицы сходны с таковыми у F-фимбрии. Биогенез этих фимбрий протекает сходным образом (см. рис. 63). В качестве секретина используется белок Vir7 (аналог субъединицы ТгаК), сквозную пору образуют субъединицы Vir9, VirlO и Vir4 (аналоги субъединиц TraBDV), а роль «сопрягающей» АТФазы играет белок Viril (аналог субъединицы ТгаС). Сборку Т-фимбрии регулируют субъединицы Vir 6 и Vir8 (аналог субъединицы TraEL).
Молекулярное строение Т-фимбрии, а также механизм секреции Т-ДНК обладают интересными особенностями. Прежде всего при процессинге пропилина ТгbС в цитоплазме бактерии происходит довольно редкое явление — циклизация белка с помощью пептидазы TraF, которая образует ковалентную связь между N- и С-концами одной и той же полипептидной цепи. В свою очередь, релаксаза VirD2 образует ковалентную связь с 5'-концом одной из цепей ДНК. Белок VirE2, который секретируется независимо от гибрида Т-ДНК с «пилотной» релаксазой, связывает Т-ДНК в цитоплазме хозяина и обеспечивает ее таргетинг в ядро.
8.6.3.4. Фимбрии архей
Убедительных доказательств того, что у архей есть фимбрии, не получено, хотя у некоторых видов родов Aeropyrum, Hyperthermus, Sulfophobococcus, Sulfurococcus и Thermodiscus(фила AI Crenarchaeota), а также у некоторых видов родов Methanobacterium, Methanobrevibacter и Methanothermobacter (фила АII Euryarchaeota) описаны похожие на фимбрии филаментозные придаточные структуры диаметром ~10 нм и длиной до 15 мкм.
Уникальным признаком экстремально термофильных архей рода Pyrodictium occultum (фила AI Crenarchaeota) служит образование «каннул» (лат. cannula — тростинка).
Эти трубчатые структуры располагаются по отдельности или в пучках и образуют трехмерный сетчатый матрикс, внутри которого покоятся клетки дискоидной
формы. Каннула представляет собой полый цилиндр длиной до 150 мкм и диаметром 25 нм. Стенка имеет толщину 2-3 нм и образована спирально уложенными гликопротеиновыми субъединицами, которые не диссоциируют даже при жесткой обработке додецилсульфатом. Хотя генетические детерминанты, состав и биогенез каннул не изучены, бросается в глаза их морфологическое, а, может быть, и функциональное сходство с фимбриями.
Таким образом, микрокомпартменты типа фимбрий встречаются у представителей обоих прокариотных доменов.