МИКРОБИОЛОГИЯ БИОЛОГИЯ ПРОКАРИОТОВ ТОМ III - А. В. ПИНЕВИЧ - 2009
ГЛАВА 16. ЦИТОГЕНЕТИКА
16.8. Эпигенетическое наследование
Современный термин «эпигенетическое» наследование (англ. epigenetic inheritance; от греч. epi— дополнять что-либо; в данном случае — наследование, не имеющее прямого отношения к генам) обозначает такие наследственные изменения характера экспрессии генов, которые непосредственно не затрагивают их первичную структуру. Иными словами, помимо нуклеотидной последовательности ДНК существует дополнительная генетическая информация, которая получает фенотипическое проявление в определенных условиях окружающей среды.
Эпигенетические системы представляют собой самосохраняющиеся альтернативные состояния белка или ДНК, которые позитивно регулируются по принципу обратной связи.
Примерами эпигенетических феноменов служит наследственное самовоспроизведение структуры белка (прионы), дифференциальная экспрессия эукариотного генома посредством метилирования или ацетилирования ДНК-связывающих белков, а также контроль прокариотных промоторов путем изменения паттерна метилирования ДНК под воздействием регуляторных белков.
Механизм эпигенетического наследования у бактерий исследован, главным образом, на примере контроля экспрессии генов с помощью метилирования специфических последовательностей ДНК.
Напомним, что у бактерий метилирование ДНК обеспечивает также:
— контроль инициации репликации (см. раздел 16.4.1.1);
— расхождение сестринских хромосом (см. раздел 16.4.2.1);
— mismatch-репарацию (см. раздел 16.5.1.3);
— транспозицию мобильных генетических элементов (см. раздел 16.3.1.3);
— контроль упаковки фаговой ДНК (см. раздел 16.5.2);
— антирестрикционное метилирование в ходе конъюгативного переноса плазмид (см. раздел 16.6.1.3).
Внутренним сигналом для реализации этих разнообразных феноменов служит «полу метилированное» состояние только что реплицированной ДНК (см. раздел 16.4.1.1). В свою очередь, это состояние регулирует специфические ДНК-белковые взаимодействия. Важно подчеркнуть, что полуметилированное состояние существует временно и как таковое не наследуется.
Большинство эпигенетических систем состоит из ДНК-метилазы Dam и ДНК- связывающих белков, которые конкурируют с метилазой за специфические сайты- мишени в ДНК. Иначе говоря, при связывании с ДНК белки защищают сайты метилирования от метилазы Dam. Метилирование, в свою очередь, мешает связыванию этих белков. В итоге, сайт-мишень существует в альтернативных состояниях — метилированном и неметилированном.
Фазовая изменчивость. Наиболее известный пример эпигенетического наследования связан с фазовой изменчивостью (англ. phase variation).
В частности, фазовую изменчивость проявляют уропатогенные штаммы Е. coli, способные образовывать фимбрии II типа. В данном случае их называют «фазовыми» Рар-фимбриями (сокр. англ. pielonephritis-associated pili). Экспрессия генов при этом носит альтернативный характер — Рар-фимбрии либо образуются (англ. phase on), либо не образуются (англ. phase off).
Фимбрии, или пилусы (см. I том учебника), образующиеся у возбудителей пиелонефрита, играют важную роль в прикреплении уропатогенных бактерий к эпителиальной выстилке верхнего отдела мочеточников, что, в конечном счете, способствует колонизации почек. В генетически однородной популяции бактерий одновременно присутствуют клетки с Рар-фимбриями и без них.
Истинный смысл такой фазовой изменчивости неизвестен. Возможно, это позволяет бактериям: (1) уклониться от иммунной защиты хозяина; (2) путем попеременной адгезии-открепления подняться вверх по мочеточнику; (3) преодолеть контактное ингибирование их роста и размножения.
Фазовая изменчивость Рар-фимбрий определяется ДНК-метилазой Dam (см. раздел 16.4.1.1), которая метилирует остатки аденина в двух сайтах GATC. Один из них, GATCPRXзанимает проксимальное положение, а другой, GATCDIS — дистальное положение по отношению к общему промотору генов, кодирующих пилины РарАВ (рис. 220).
Рис. 220. Механизм перехода между состояниями «phase on» и «phase off». А — регуляторный участок оперона рар; Б — состояние «phase off»; В — состояние «phase on».Цифрами отмечены сайты для связывания белка Lrp, пунктирная стрелка указывает направление транскрипции.
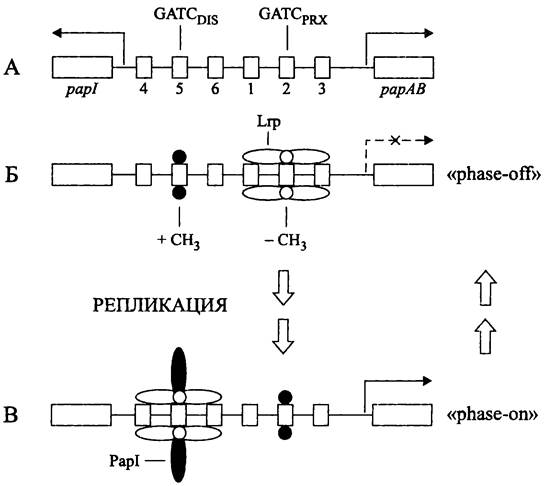
В промежутке между генами papl и рарАВ расположены сайты 1-6 для связывания глобального регуляторного белка Lrp (см. раздел 18.1.2). Внутри 5-го и 2-го сайтов, соответственно, находятся GATCDIS и GATCPRX. Благодаря этому метилирование контролирует эффективность связывания белка Lrp, а также связывание впомогательного регулятора — белка Papi (промотор его гена находится левее GATCDIS).
Когда клетки находятся в состоянии «phase он», белки Lrp/PapI взаимодействуют с GATCDIS. защищая его от метилазы Dam. При этом GATCPRXостается свободным и метилируется. В состоянии «phase off» GATCPRX защищен только белком Lrp и не метилируется, a GATCDIS метилируется (рис. 220). Противоположные паттерны метилирования создаются в результате конкуренции между белками Dam и Lrp за сайты связывания 1-3 и 4-6. Блокада транскрипции генов рарАВ в состоянии «phase off» объясняется тем, что белок Lrp перекрывает промотор, изолируя его от РНК-полимеразы.
Переход «phase off» —> «phase on» происходит посредством полного метилирования GATCPRX и полного деметилирования GATCDIS - Предполагается, что для этого требуются два раунда репликации ДНК, с промежуточным образованием полуметилированных сайтов, когда белки Lrp/PapI имеют более высокое сродство к полуметилированному GATCDIS, чем к полностью метилированному.
Есть идея, что при обратном переходе «phase on» —> «phase off», который также требует репликации ДНК, белки Lrp/PapI покидают сайты 4-6, что дает возможность метилазе Dam конкурировать с белком Lrp за связь с GATCdis (рис. 220). Так или иначе, фазовая изменчивость Рар- фимбрий имеет очень сложный, лишь отчасти расшифрованный механизм.
Существует целый ряд «Pap-подобных» систем, выступающих в роли позитивных регуляторов фазовой изменчивости. В числе генов и оперонов, которые регу-
лируются при помощи данного механизма, следует отметить ген белка Ag43 наружной мембраны Е. coli; гены репарационной системы VSP энтеробактерий (сокр. англ. very shortrepair); гены tra-оперона, обеспечивающего конъюгативный перенос плазмиды Salmonella entérica серовар Typhimurium, и т. д.
Помимо белка Lrp, к числу ДНК-связывающих белков, блокирующих Dam-мети- лирование, относятся другие глобальные регуляторы транскрипции, например, белок OxyR (см. раздел 18.1).
Следует подчеркнуть, что в основе фазовой изменчивости могут лежать и неэпигенетические механизмы. В этом случае она обусловлена не преобразованием конфигурации ДНК или модификацией взаимодействия между матрицей и РНК-полимеразой, а изменением нуклеотидной последовательности в результате сайтспецифичной рекомбинации или мутации.
Субпопуляции симбиотических бактерий, особенно комменсалов или патогенов, вынужденных оперативно адаптироваться к изменяющимся условиям окружающей среды, обладают повышенной частотой спонтанного мутирования (см. раздел 16.6.1.1). Негативная сторона данного явления заключается в том, что штаммы-мутаторы и штаммы-гипермутаторы неизбежно накапливают вредные мутации.
Чтобы избежать этого, симбиотические бактерии выработали повышенную способность к изменениям генома только в участках, рассчитанных на непредвиденные обстоятельства (англ. contingency loci). Такие участки отвечают, главным образом, за структуру или секрецию детерминант, обеспечивающих контакт бактерии с ее хозяином (липополисахаридов, жгутиков, фимбрий и др.).
Воздействие на экспрессию генов в этих участках осуществляется разными способами, основанными на обратимых превращениях нуклеотидной последовательности или конфигурации ДНК. В обоих случаях это приводит к фазовой изменчивости — соответственно, эпигенетической и неэпигенетической, причем эта изменчивость может быть альтернативной или мультивариантной.
Неэпигенетические изменения чаще всего связаны с увеличением или уменьшением числа микросаттеллиттов (англ. microsatellite), или коротких повторяющихся последовательностей, а также длины этих последовательностей, что приводит к сдвигу рамки считывания. Кроме того, путем инверсии или сайтспецифичной рекомбинации локально изменяются кодирующе участки ДНК.
Фазовая изменчивость и эпигенетического, и неэпигенетического типа носит вероятностный характер. В результате нее создаются случайные комбинации фенотипов, конкурентная пригодность которых тестируется быстро меняющимися условиями окружающей среды.
* * *
На этом завершается обзор цитогенетики прокариотов.
Знание генетических структур, процессов и программ позволит в дальнейшем познакомиться с реализацией разнообразных вегетативных и дифференцированных паттернов развития у прокариотов. Гибкость адаптации дает возможность бактериям и археям существовать в разнообразных природных нишах, а также в лабораторной обстановке или на биотехнологическом производстве.