Молекулярная биология: Структура и функции белков - Степанов В.М. 2005
Выделение белов
Аффинная (биоспецифическая) хроматография белков
Как уже отмечалось, способы разделения белков, основанные на различиях в их физико-химических свойствах, не могут быть высокоспецифичными. С этой точки зрения гораздо перспективнее препаративные методы, которые базируются на функциональных различиях белков. Для множества белков, включая ферменты, их ингибиторы, транспортные белки, иммуноглобулины, регуляторные белки, токсины, рецепторы, главнейшая функциональная черта состоит в способности избирательно связывать те или иные лиганды — субстраты, коферменты, аллостерические эффекторы, антигены и т.п.
Такое связывание по существу своему весьма специфично, и это свойство резко выделяет тот или иной белок из множества других. На использовании функционально обусловленной способности белков обратимо связываться с соответствующими лигандами и основан метод аффинной, или биоспецифической, хроматографии.
Для синтеза аффинного сорбента отвечающий специфичности данного белка лиганд (субстрат или его аналог при хроматографии фермента) присоединяют к инертной матрице так, чтобы возможно меньше затронуть те элементы его структуры, которые непосредственно участвуют во взаимодействии с белком. Иногда лиганд прямо вводят в реакцию с функциональными группами на поверхности матрицы, однако чаще их соединяют через промежуточное звено («вставку», «ножку» и т.п.), чтобы отдалить лиганд от матрицы и уменьшить пространственные препятствия для его сближения с белком.
К каждому из элементов структуры аффинного сорбента предъявляются определенные требования. Матрица должна быть инертной и не создавать стерических препятствий для белка. Чаще всего в качестве матрицы используют макропористые гели, образованные поперечно-сшитыми гидрофильными полимерами, например производное агарозы — сефарозу, синтетические полимеры или неорганические носители, макропористые производные кремнезема — силикагель или стекло Присоединение лигандов к сефарозе (непосредственно или через вставку) обычно проводят, активируя ее бромцианом, Бромциан (сильный яд!) в щелочной среде реагирует с гидроксильными группами сефарозы, образуя весьма активный эфир изоциановой кислоты:
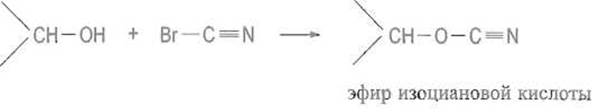
Последний взаимодействует затем с аминогруппами лиганда L или «вставки» с образованием производного изомочевины, которое является сильным основанием и в обычном диапазоне pH несет положительный заряд:
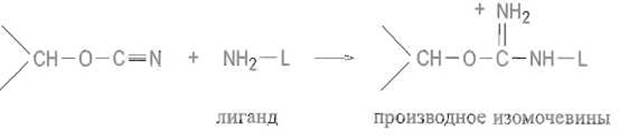
Таким образом, присоединение лиганда к сефарозе, активированной бромцианом, одновременно создает на матрице эквивалентное содержанию лиганда число катионных групп. Следовательно, такой сорбент помимо биоспецифического связывания белка лигандом может проявлять свойства анионита, что необходимо учитывать при его использовании.
Известны и другие способы присоединения лигандов к матрице, однако в каждом случае следует учитывать изменения в характере носителя, сопровождающие реакцию с лигандом, а также большую или меньшую неустойчивость образовавшейся связи, которая иногда может вызывать постепенную «утечку» лиганда. Например, изомочевина может превращаться в производное гуанидина в присутствии значительных концентраций первичных аминов или аммиака, что влечет за собой отщепление лиганда.
Сложнее требования, предъявляемые к лиганду. Он прежде всего должен достаточно сильно взаимодействовать с белком. Так, принято считать, что для полу гения сорбентов, предназначенных для выделения ферментов, в качестве лигандов могут быть использованы такие ингибиторы или аналоги субстрата, которые имеют константу ингибирования (субстратную константу), т.е. константу диссоциации фермент — лиганд, не хуже 10-4 М. При этом учитывается, что присоединение лиганда к матрице ухудшает (т.е. повышает) константу ингибирования по крайней мере на один порядок.
При подборе лигандов для аффинной хроматографии ферментов приходится изменять структуру субстрата, чтобы предотвратить возможность его превращения ферментом в продукт, например, расщепления пептидного лиганда, если речь идет о хроматографии протеиназ. Очевидно, что сорбент, содержащий в качестве лиганда истинный субстрат, будет трансформирован при первом же контакте с ферментом и окажется непригодным.
При пропускании раствора, содержащего сложную смесь белков, других биополимеров, низкомолекулярных соединений, солей, пигментов и т.п., через биоспецифический сорбент, построенный по описанной выше схеме, лиганд образует комплекс только с тем белком, который имеет участок связывания, комплементарный структуре лиганда. В результате именно этот белок удерживается аффинной колонкой, тогда как все остальные компоненты смеси проходят через нее не задерживаясь.
После промывания колонки для удаления неспецифически удерживаемых примесей белок элюируют, изменяя состав протекающего через колонку раствора так, чтобы ослабить взаимодействие белка с лигандом. Для этого прибегают к изменению pH, добавляют к элюенту неорганические соли, органические растворители. Все эти факторы, воздействуя на структуру зоны связывания лиганда и подавляя отдельные виды белок-лигандньгх взаимодействий, например ионные связи и гидрофобные контакты, обеспечивают десорбцию. В отдельных случаях прибегают к аффинной элюции раствором лиганда или его аналога. Этот прием, основанный на конкуренции за связывание белка между присоединенным к матрице и растворенным лигандами, весьма специфичен и эффективен, но дорог.
В качестве примера рассмотрим применение циклопептидного антибиотика — грамицидина S (см. гл. 2) — в качестве лиганда при аффинной хроматографии протеолитических ферментов. Он содержит ряд гидрофобных аминокислот, соответствующих специфичности многих протеиназ, и два остатка орнитина, δ-аминогруппы которых позволяют легко присоединить циклопептид к активированной бромцианом сефарозе или другим матрицам:
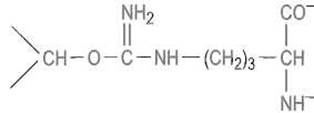
остаток орнитина, аминогруппа которого присоединена к активированной бромцианом сефарозе
Грамицидин S связывает протеиназы различных классов, однако он устойчив к их действию и не подвергается гидролизу, по-видимому, из-за присутствия в структуре остатков D-фенилаланина, пролита, а также из-за особенностей конформации цикпопептида. Следовательно, грамицидин S является природным аналогом субстратов протеиназ и хорошо подходит для роли лиганда широкой специфичности. Так, при пропускании через колонку с грамицидин - S-сефарозой сложной смеси веществ, содержащихся в культуральной жидкости одного из штаммов бактерии Bacillus subtilis, сорбент связывает только металлопротеиназу, которую удается полностью освободить от примесей и получить практически чистый белок.
Еще эффективнее сорбенты, содержащие такие лиганды, которые взаимодействуют не только с зоной связывания, но и с каталитическим центром фермента. Например, производные бензилянтарной кислоты оказались хорошими лигандами для выделения карбоксипептидаз — ферментов, отщепляющих аминокислоты от карбоксильного конца пептидной цепи. Сходство лиганда с субстратами карбоксипептидаз на первый взгляд невелико, однако оказывается, что группа НООС—СН2 способна выступать в качестве своеобразного аналога пептидной связи:

В результате ß-карбоксильная группа бензилянтарной кислоты взаимодействует с каталитическим центром фермента, а боковая бензильная группа и а-карбоксил — с компонентами зоны связывания субстрата. Образующийся комплекс лиганда, обычно присоединяемого к матрице через аминогруппу, введенную в пара-положение бензольного кольца, связывает фермент весьма прочно.
Широко используют своеобразный вариант аффинной хроматографии, основанный на применении в качестве лигандов синтетических красителей антрахинонового ряда, например цибакрона голубого:
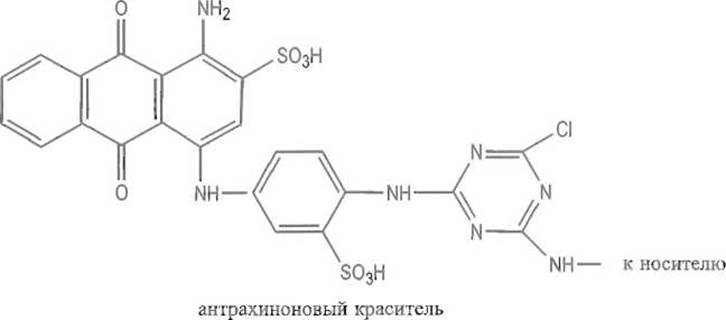
Плоская структура, образованная тремя конденсированными ароматическими кольцами замещенного антрахинона, к которому в положении 1 присоединены замещенный анилин и триазиновый цикл, способна избирательно взаимодействовать с целым рядом белков. Лиганд связывается, по-видимому, во впадинах, которые нередки на поверхности белков, в особенности в активных центрах ферментов. Высказывались предположения, что антрахиноны вместе с присоединенными ядрами анилина и триазина образуют структуру, стерически близкую природным лигандам, например таким коферментам, как NAD, и имитируют свойственный последним способ связывания с белками. Ввиду этого сорбенты, содержащие цибакрон голубой и некоторые другие антрахиноновые красители, особенно рекомендуются для аффинной хроматографии белков, содержащих NAD или иные соединения нуклеотидной природы.
Нет необходимости, чтобы нуклеотид или сходное по строению соединение было похоже на субстрат — оно может имитировать эффектор, регулирующий активность фермента. Так, например, хорошие результаты дает хроматография на антрахиноновых красителях аспартаткарбамилазы, где нуклеотиды выступают как аллостерические эффекторы.
Впрочем, круг белков, связываемых этими сорбентами, значительно шире и включает, например, сывороточный альбумин. Последний легко присоединяет различные лиганды, в том числе некоторые лекарственные препараты и триптофан, который можно (с большими оговорками) считать напоминающим конденсированные кольца антрахинона. Структура лиганда — замещенного антрахинона — не безразлична, поэтому, используя разные красители, удается получить серию сорбентов, не одинаково связывающих различные белки.