ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 24. Гиперчувствительность - тип II
■ Реакции гиперчувствительности II типа опосредованы IgG- или IgM-антителами к антигенам клеточной поверхности и внеклеточного матрикса. Может происходить и образование антител к внутриклеточным компонентам, но они обычно непатогенны, хотя могут иметь диагностическое значение.
■ Трансфузионные реакции на эритроциты вызываются антителами к антигенам групп крови; образование таких антител может происходить независимо или в результате индукции предшествующим контактом с несовместимой тканью или кровью при трансплантации, переливании крови или беременности.
■ Антитела повреждают клетки и ткани вследствие активации комплемента, а также связывания и активации эффекторных клеток, несущих Fсγ-рецепторы.
■ Гемолитическая болезнь новорожденных развивается в тех случаях, когда материнские антитела к групповым антигенам крови плода проходят через плаценту и разрушают его эритроциты.
■ Повреждение тканей могут вызывать антитела к базальным мембранам, молекулам межклеточной адгезии или рецепторам. Характер патологии зависит от молекул- и тканей-мишеней.
Антитела IgG и IgM, связываясь с определенными клетками или тканями, вызывают развитие реакций гиперчувствительности II типа. Локализация возникающих повреждений ограничена тем самым клетками или тканями, экспонирующими соответствующие антигены. Патогенность, как правило, характерна для антител к антигенам клеточной поверхности, тогда как антитела к внутриклеточным антигенам обычно непатогенны. В отличие от этого в реакциях III типа участвуют антитела к растворимым антигенам сыворотки, вызывающие образование в крови комплексов антиген—антитело. Повреждения в данном случае связаны с неспецифическим отложением таких комплексов в тех или иных тканях и/или органах (см. гл. 25).
МЕХАНИЗМЫ ПОВРЕЖДЕНИЙ
При гиперчувствительности II типа повреждение клеток-мишеней обусловлено взаимодействием антител к антигенам клеточной поверхности или тканей с комплементом и различными эффекторными клетками (рис. 24.1).
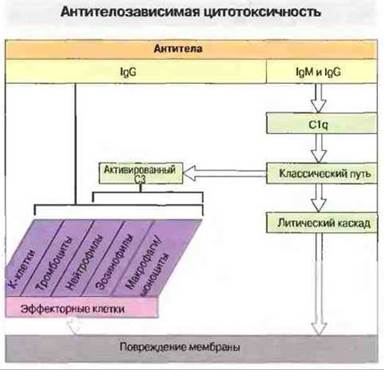
Рис. 24.1. Все эффекторные клетки (К-клетки, тромбоциты, нейтрофилы, эозинофилы и клетки мононуклеарно-фагоцитарного ряда) обладают рецепторами к Fc-фрагменту, посредством которых они взаимодействуют с антителами, фиксированными в тканях на клетках-мишенях. Активация С3 может непосредственно приводить к комплемент-опосредованному лизису клеток-мишеней или способствовать связыванию фагоцитов с их мишенями - при участии С3b, C3bi или C3d, которые одновременно активируют фагоциты.
Прикрепившиеся к поверхности клеток или тканей антитела могут связывать и активировать компонент С1 комплемента, вызывая следующие эффекты (см. гл. 4).
✵ Фрагменты комплемента (С3а и С5а), образующиеся при его активации, привлекают к данному участку макрофаги и полиморфноядерные клетки, а также стимулируют продукцию тучными клетками и базофилами молекул, привлекающих и активирующих другие эффекторные клетки (см. гл. 10).
✵ Активация комплемента по классическому пути и действие механизма усиления приводят к отложению С3b, С3bi и C3d на мембране клеток-мишеней.
✵ Активация комплемента по классическому пути на ее конечной стадии приводит к образованию лизирующего мембрану комплекса (С5b-9), который встраивается в мембрану клетки-мишени.
Эффекторные клетки — в данном случае макрофаги, нейтрофилы, эозинофилы и К-(киллерные) клетки — взаимодействуют посредством своих Fc-рецепторов с фиксированными на клетках антителами или через свои С3-рецепторы с мембраносвязанными С3b, С3bi и C3d (см. рис. 24.2 и гл. 5). Связывание антител с Fc-рецепторами фагоцитов стимулирует продукцию ими большего количества лейкотриенов и простагландинов, которые выполняют важную роль в процессе воспаления (см. гл. 5). Хемокины и хемотаксические молекулы, в том числе С5а, лейкотриен В4(LТВ4) и пептиды фибрина, также могут активировать поступающие в участок реакции клетки.
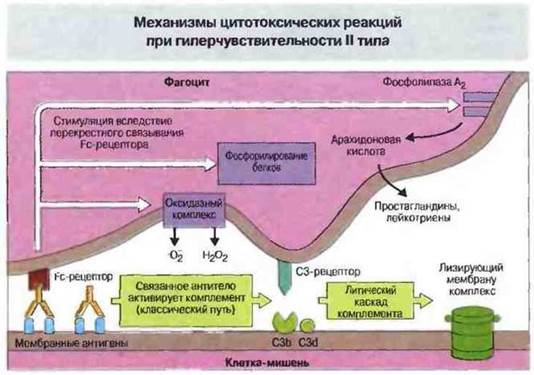
Рис. 24.2. Антитела, связываясь с мембранными антигенами клеток-мишеней, опсонизируют их, способствуя фагоцитозу. Перекрестное связывание антител с Fc-рецепторами на поверхности фагоцитов активирует мембранный оксидазный комплекс этих клеток, приводя к выделению ими кислородных радикалов; одновременно усиливается и фосфорилирование белков, т. е. происходит активация эффекторных клеток. Под действием фосфолипазы А2 из мембранных фосфолипидов высвобождается арахидоновая кислота. Иммунные комплексы индуцируют отложение фрагмента С3b комплемента, также способного взаимодействовать с рецепторами фагоцитов. Активация литического каскада приводит к сборке лизирующего мембрану комплекса (ЛМК) из компонентов комплемента С5-С9.
Прочно связанные с клетками-мишенями и полностью активированные эффекторные клетки могут вызывать значительные повреждения (см. гл. 10).
Антитела разных изотипов различаются по способности индуцировать такие реакции, что зависит от эффективности связывания ими C1q или взаимодействия с Fc-рецепторами эффекторных клеток. Фрагменты комплемента или IgG могул выступать в роли опсонинов, связываясь с тканями хозяина или с микроорганизмами; опсонизированные частицы поглощаются фагоцитами. Повышая активность лизосомных ферментов фагоцитов и продукцию ими высокоактивных метаболитов кислорода, опсонины увеличивают способность этих клеток не только разрушать патогенные микробы, но и вызывать иммунопатологические реакции гиперчувствительности II типа (рис. 24.3). Например, у больных ревматоидным артритом нейтрофилы из синовиальной жидкости при стимуляции выделяют больше супероксид-анионов по сравнению с нейтрофилами из крови. Это считают следствием активации нейтрофилов в пораженных суставах под действием таких медиаторов, как иммунные комплексы и фрагменты комплемента.
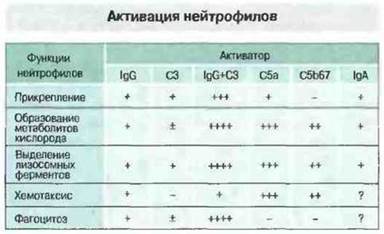
Рис. 24.3. Нейтрофилы активируются фиксированными на клетках антителами (IgG или IgA) и активированными компонентами комплемента. Каждый из этих активаторов стимулирует определенные функции нейтрофилов Обратите внимание, что активированный С3 (включая С3b, C3bi и C3d - в зависимости от зрелости участвующих в реакции клеток) и IgG усиливают эффекты друг друга. Действуя вместе, они формируют особенно сильный активационный сигнал для эффекторных клеток.
Эффекторные клетки вызывают характерные для гиперчувствительности II типа повреждения клеток собственного организма посредством тех же механизмов, какими они действуют на инфекционные агенты (рис. 24.4). Так, большинство патогенных микробов (если они не резистентны к воздействию фагоцитов) уничтожается внутри фаголизосом в результате совместного действия высокоактивных метаболитов кислорода и азота, радикалов, ионов, ферментов, изменения pH и влияния других факторов, обеспечивающих лизис. Если объект слишком крупный для фагоцитоза, эффекторные клетки выделяют содержимое своих гранул и лизосом в направлении сенсибилизированной мишени (экзоцитоз) (рис. 24.4). В определенных случаях, например при эозинофильной реакции на инвазию шистосом (см. гл. 18), выброс содержимого гранул обеспечивает защиту; когда же мишенью оказываются сенсибилизированные антителами клетки хозяина, эта реакция приводит к повреждению собственных тканей (рис. 24.5).
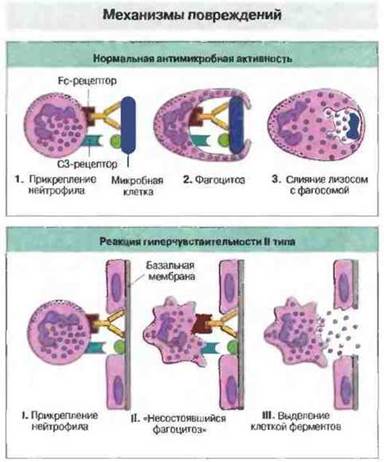
Рис. 24.4. Повреждающее действие нейтрофилов на собственные ткани - это отражение нормальной антибактериальной функции этих клеток. 1. Нейтрофилы взаимодействуют с микробами через свои Fc- и С3-рецепторы. 2. Затем микробную клетку поглощает фагоцит, внутри которого она разрушается по мере слияния лизосом и фагосомы с образованием фаголизосомы (3). При реакциях гиперчувствительности II типа отдельные нагруженные антителами клетки хозяина также могут подвергаться фагоцитозу, но при крупных размерах мишени, если это, например, базальная мембрана (I), нейтрофилы неспособны фагоцитировать ее (II) и выделяют содержимое лизосом наружу, повреждая близлежащие клетки (III).
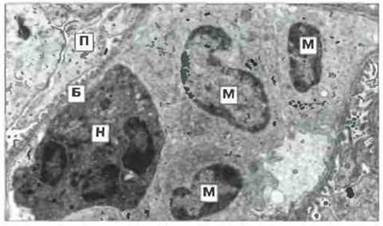
Рис. 24.5. Фагоциты, атакующие базальную мембрану. На электронной микрофотографии видны нейтрофил (Н) и три моноцита (М), связанные с базальной мембраной (Б) капилляра почки кролика, у которого имеются антитела к базальной мембране. (П - подоцит.) х 3500. (Фото любезно предоставлено проф. G.A. Andres.)
Антитела вызывают реакцию гиперчувствительности и путем перекрестного связывания К-клеток с тканями-мишенями. К-клетки присутствуют главным образом в популяции больших гранулярных лимфоцитов и связываются с антителами через свои высокоаффинные Fc-рецепторы. В условиях in vitro они оказывают цитотоксическое действие на клетки многих типов, осуществляя сто преимущественно посредством цитокинов и путем экзоцитоза гранул. Однако оценить вклад активности К-клеток в развитие реакций гиперчувствительности II типа довольно трудно, отчасти из-за того, что не существует двух клеток-мишеней, обладающих равной чувствительностью к данной эффекторной клетке. Чувствительность клеток-мишеней зависит от количества экпрессированного на их поверхности антигена и от их способности к репарации повреждений. Например, для лизиса одного эритроцита достаточно одного лизирующего мембрану комплекса, тогда как для лизиса большинства ядерных клеток требуется много таких комплексов.
Ниже рассмотрен ряд ситуаций, в которых, как принято считать, реакции гиперчувствительности II типа служат основной причиной деструкции клеток-мишеней, т. е. иммунопатологии.