ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 26. Гиперчувствительность - тип IV
КЛЕТОЧНЫЕ РЕАКЦИИ ПРИ ГИПЕРЧУВСТВИТЕЛЬНОСТИ IV ТИПА
Эксперименты на мышах, лишенных тех или иных генов методом генного нокаута, подтвердили, что для возникновения реакций замедленного типа в ответ на бактериальную внутриклеточную инфекцию необходимы Т-клетки, несущие αβ-, а не γδ-ТкР. Сенсибилизированные αβ-Т-клетки, стимулированные соответствующим антигеном, АПК, перед делением претерпевают бласттрансформацию (рис. 26.14). На этом основан тест для определения стимуляции лимфоцитов (см. гл. 29). Стимуляция лимфоцитов сопровождается синтезом ДНК, который можно выявить по включению радиоактивно меченного тимидина (нуклеотида, необходимого для синтеза ДНК). Тест состоит в том, что лимфоциты больного культивируют с вероятным антигеном, проверяя, вызывает ли он трансформацию клеток. Положительный результат данного теста указывает, однако, только на присутствие Т-клеток иммунологической памяти и не обязательно на защитный иммунитет.
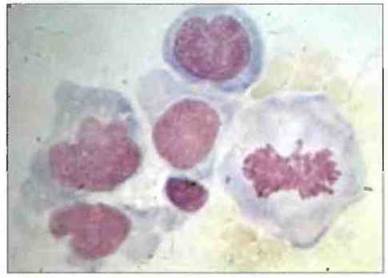
Рис. 26.14. Трансформированные лимфоциты. Активированные соответствующим антигеном Т-клетки, прежде чем делиться, претерпевают бласттрансформацию. Микрофотография демонстрирует клетки-бласты с увеличенными ядрами (один из лимфоцитов находится в метафазе клеточного деления).
После активации, вызванной АПК, Т-клетки выделяют многочисленные провоспалительные цитокины, которые привлекают и активируют макрофаги. К таким цитокинам относятся ИФγ, лимфотоксин, ИЛ-3 и ГМ-КСФ. Преобладание Tx1-клеточных цитокинов связано с активацией Т-клеток цитокином ИЛ-12, который выделяют макрофаги под действием бактериальных продуктов; этот цитокин подавляет выделение цитокинов Тх2-кпетками. Роль отдельных цитокинов можно проанализировать в опытах на нокаутных мышах, лишенных гена определенного цитокина. Например, мыши, лишенные гена ИФγ, не способны активировать макрофаги и бороться с туберкулезной инфекцией {рис. 26.15). При гранулематозных реакциях активированные макрофаги служат основным источником ФНО и гранулема развивается за счет аутоамплификации с превращением макрофагов в эпителиоидные клетки (рис. 26.16 и 26.17). Последние продолжают секретировать ФНО, стимулируя дальнейшее образование эпителиоидных клеток, которые, сливаясь, формируют гигантские клетки (рис. 26.18).
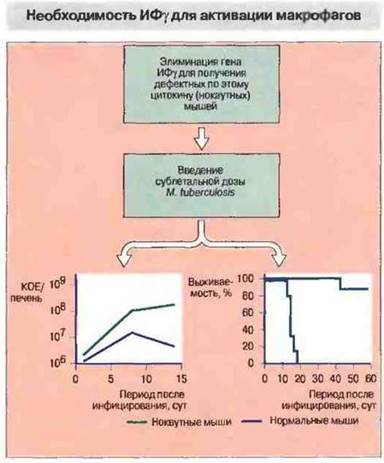
Рис. 26.15. У мышей, лишенных гена ИФγ, отсутствует активация макрофагов при внутриклеточной бактериальной инфекции. Макрофаги, хотя и накапливаются в очаге инфекции, не образуют типичных гранулем. Неконтролируемая инфекция (график слева) вызывает обширный некроз тканей и гибель животных (график справа).
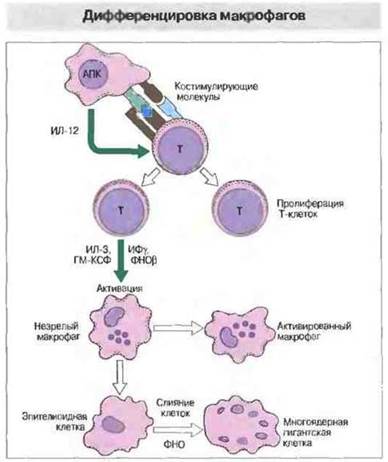
Рис. 26.16. Бактериальные продукты стимулируют секрецию макрофагами ИЛ-12. Активация Т-клеток этим цитокином приводит к выделению ИФγ и других цитокинов - ФНОβ, ИЛ-3 и ГМ-ГСФ, которые активируют макрофаги, помогая уничтожению внутриклеточных паразитов. Неспособность ликвидировать антигенный стимул обусловливает постоянное выделение цитокинов и дифференцировку макрофагов в эпителиоидные клетки, которые секретируют большое количество ФНОα. Некоторые эпителиоидные клетки сливаются, образуя многоядерные гигантские клетки.
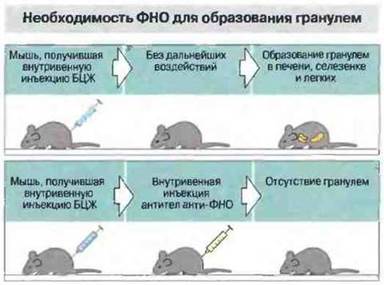
Рис. 26.17. ФНО необходим для образования эпителиоидно-клеточных гранулем. Если получившим БЦЖ мышам ввести антитела анти-ФНОα, гранулемы у них не образуются.
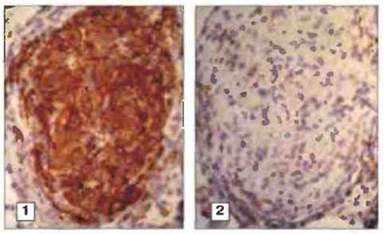
Рис. 26.18. Эпителиоидные клетки в гранулеме из легкого при саркоидозе. 1. Эпителиоидные клетки и гигантские клетки в центре окрашены специфическими антителами RFD-9. 2. Окружающие гранулему зрелые тканевые макрофаги, окрашенные специфическими антителами RFD-7. (Микрофотографии любезно предоставлены С. S. Munro.)