ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 8. Источники разнообразия антигенраспознающих структур
ВАРИАБЕЛЬНОСТЬ ИММУНОГЛОБУЛИНОВ
Молекулы иммуноглобулинов состоят из легких и тяжелых полипептидных цепей. Легкие цепи относятся к k или λ-типу. Теоретически любая легкая цепь может объединяться с любой тяжелой, поэтому количество возможных антигенсвязываюших центров разной специфичности равно произведению числа цепей одного типа и числа цепей другого. Например, если имеется 104различных легких цепей, способных объединиться с любой из 104 разных тяжелых цепей, то вполне возможно появление 108 молекул антител различной специфичности. У каждою типа цепей существует свой собственный механизм увеличения многообразия, поскольку они колируются генами, локализованными в разных хромосомах (рис. 8.3).
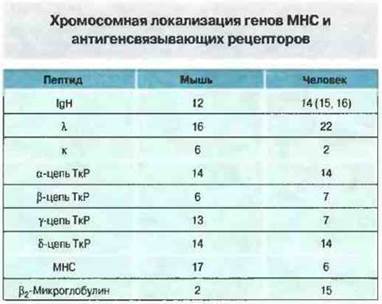
Рис. 8.3. Числа означают номера хромосом, в которых локализованы гены, кодирующие различные полипептиды иммуноглобулинов, ТкР и продуктов МНС человека и мыши. Заметьте, что все локусы располагаются совершенно раздельно, за исключением локуса δ-цепей ТкР, локализованного среди генов локуса α-цепей ТкР.
Полиморфизм иммуноглобулинов проявляется как вариабельность разных частей их молекул (рис. 8.4.). Рассмотрим вначале идиотипическую вариабельность, обусловленную разнообразием структуры антигенсвязываюших центров.
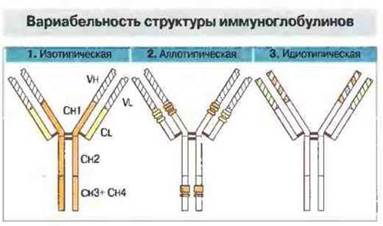
Рис. 8.4. Все иммуноглобулины построены из разного количества сходных четырехцепочечных единиц и могут иметь варианты трех типов:
1. Изотипические варианты, обусловленные экспрессией гаметных генов, присутствующих у всех особей данного вида и кодирующих тяжелые (μ, δ, γ, ε, α) и легкие (k и λ) цепи, а также «каркасные» аминокислотные остатки в составе их V-областей (подгруппы).
2. Аллотилические варианты, обусловленные внутривидовой аллельной изменчивостью.
3. Идиотипические варианты, представляющие разнообразие антигенсвязывающих центров (паратопов) и обусловленные, в частности, изменчивостью гипервариабельных участков V-областей.
By и Кэботом был проведен анализ аминокислотных последовательностей большого числа легких и тяжелых цепей различных по специфичности антител. Источником идентичных иммуноглобулинов служили миеломы (моноклональные В-клеточные опухоли). Результаты показали, что вариабельность легких цепей λ-типа обусловлена аминокислотными заменами в трех гипервариабельных участках, окруженных относительно неизменными «каркасными» остатками. Позднее, когда было установлено, что именно эти гипервариабельные участки контактируют с антигеном, они были названы участками, «определяющими комплементарность» (CDR, от англ. complementarity-determining regions).
Вначале были изучены CDR мыши — вида, особенность которого состоит в ограниченном разнообразии λ-цепей (их содержат менее 5% антител) из-за весьма малого числа Vλ-генов. Так, например, из 19 λХ-цепей, для которых была определена аминокислотная последовательность, 12 оказались идентичными, а 7 остальных отличались одна от другой и от последовательности, принятой за прототип, всего лишь несколькими остатками (рис. 8.5).
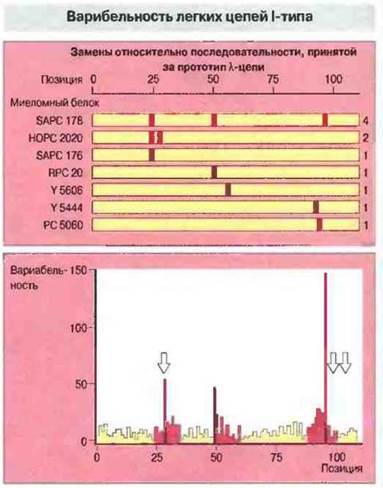
Рис. 8.5. Аминокислотная последовательность семи мышиных миеломных λ1-белков. В позициях, окрашенных желтым цветом, находятся те же аминокислотные остатки, что и в последовательности, принятой за прототип (МОРС 104Е). Красным цветом отмечены позиции, в которых имеются замены. Справа указано число специфических нуклеотидных замен в ДНК, которые требуются, чтобы получить наблюдаемые изменения в аминокислотной последовательности. В нижней части рисунка помещен график вариабельности аминокислотных остатков в разных позициях легких цепей (по Кэботу и By), рассчитанной как частное от деления числа различных остатков в данной позиции на частоту наиболее часто встречающегося остатка (ср. гл. 6). В некоторых цепях были обнаружены дополнительные остатки (в позициях, указанных стрелками), но здесь для лучшей сопоставимости они опущены.
Вариабельные остатки в составе тяжелых цепей подобным же образом сосредоточены в трех гипервариабельных участках (CDR), с обеих сторон окруженных почти неизменными «каркасными» участками (рис. 8.6), которые по сходству или идентичности можно разделить на группы (рис. 8.7).
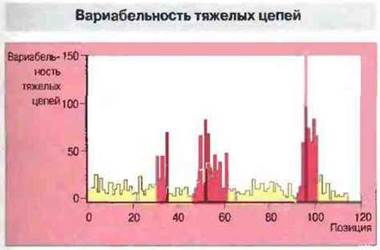
Рис. 8.6. График By и Кэбота, показывающий, что вариабельные аминокислотные остатки сосредоточены в трех участках V-области тяжелых цепей.
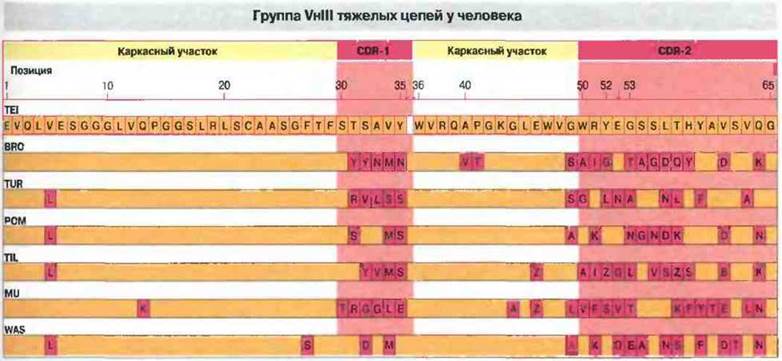
Рис. 8.7. Аминоконцевая последовательность (позиции 1-65) попавших в группу VhIII тяжелых цепей шести миеломных белков человека сопоставлена на диаграмме с последовательностью TEI, принятой за прототип. Остатки, идентичные прототипу, окрашены желтым цветом, а отличающиеся в тех же позициях - темно-красным. Большинство различий между тяжелыми цепями, относящимися к одной группе, сосредоточено в участках, определяющих комллементарность - CDR-1 и CDR-2. Участок CDR-3 на рисунке не показан.