ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 8. Источники разнообразия антигенраспознающих структур
■ Благодаря огромному разнообразию антител, синтезируемых В-клетками, и антигенраспознающих рецепторов, экспрессируемых Т-клетками, иммунная система способна распознавать множество различных антигенов и отвечать на них.
■ Молекула иммуноглобулина состоит из тяжелых и легких цепей; легкие цепи могут относиться к k- или λ-типу. Общее количество возможных вариантов антигенсвязывающих центров рассчитывается как произведение чисел различных тяжелых и легких цепей.
■ Легкие цепи иммуноглобулинов кодируются генными сегментами V и J; в кодировании тяжелых цепей также участвуют сегменты V и J, но дополнительное разнообразие вносят сегменты D.
■ Рекомбинации ограниченного числа генных сегментов V, D и J создают бесконечное число вариабельных доменов разной специфичности.
■ После антигенной стимуляции в генах легких и тяжелых цепей иммуноглобулинов происходят точечные соматические мутации. Гены Т-клеточных рецепторов при этом не подвергаются изменениям.
■ В кодировании ТкР участвуют четыре группы генов: гены α и β экспрессирует большинство периферических Т-клеток, а гены γ и δ - одна из субпопуляций Т-клеток тимуса и небольшая часть периферических Т-клеток.
■ Разнообразие ТкР, подобно разнообразию антител, создается в результате рекомбинаций между генными сегментами V, D и J, происходящих в каждом из локусов α-, β-, γ- или δ-цепей с небольшими различиями в механизмах.
■ Рекомбинацию генных сегментов V, D и J, кодирующих иммуноглобулины и Т-клеточные рецепторы, регулируют (по крайней мере отчасти) два активирующих ее гена (RAG-1 и RAG-2).
■ Кроме простых перестановок генных сегментов V, D и J в создании разнообразия иммуноглобулинов и ТкР имеют значение вставки добавочных нуклеотидов («N-региональная» вариабельность), изменение позиций стыковки генных сегментов и рамок считывания сегментов D.
■ Переключение изотипа иммуноглобулинов обусловлено рекомбинацией VDJ-генов с различными С-генами и дифференциальным сплайсингом РНК.
Способность иммунной системы распознавать антигены целиком зависит от антител, синтезируемых В-клетками, и антигенсвязывающих рецепторов, экспрессируемых Т-клетками. Обе эти клеточные популяции способны распознать множество разнообразных антигенов, но разными путями. Хотя антитела отличаются от Т-клеточных рецепторов (ТкР), разнообразие антигенной специфичности тех и других формируют весьма сходные механизмы, которые и будут рассмотрены в данной главе.
Благодаря своему поразительному разнообразию по специфичности центров связывания антигена антитела обеспечивают распознавание миллионов различных антигенов, встречающихся в окружающей среде. Кроме того, у антител каждого класса имеется характерная эффекторная область молекулы: например, IgE может связываться е Fc-рецепторами тучных клеток, тогда как IgG способен присоединяться к фагоцитам. Подсчитано, что структурных вариантов антител в организме образуется гораздо больше, чем всех прочих белков вместе взятых. Число синтезируемых организмом вариантов антител фактически превышает количество генов в нашем геноме. Как может возникать разнообразие такого масштаба? Первоначальные представления о процессах образования антител с годами существенно изменились, но вес же вызывает удивление, как удалось Эрлиху в начале столетия своей гипотезой боковых цепей вплотную приблизиться к современным взглядам (рис. 8.1). Его идея о селекции антигеном клеток, образующих антитела, почти совпадает с современной клонально-селекционной теорией, исключая размещение нескольких рецепторов разной специфичности на одной и той же клетке.
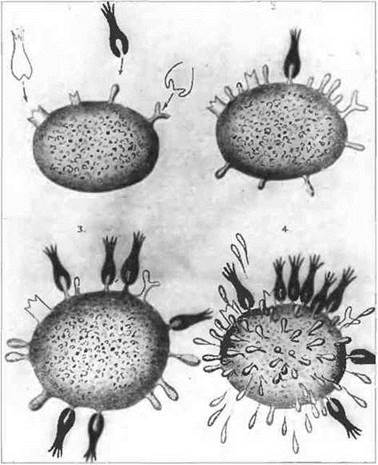
Рис. 8.1. Предложенная Эрлихом теория боковых цепей. Эрлих предположил, что соединение антигена с уже имеющимся рецептором на поверхности В-клетки (теперь известно, что это мембраносвязанный иммуноглобулин) заставляет ее синтезировать и секретировать повышенное количество таких рецепторов. Хотя, как показано на рисунке, Эрлих считал, что одна клетка способна производить антитела, связывающие более чем один тип антигена, тем не менее он предвосхитил и клонально-селекционную теорию иммунитета, и фундаментальное представление о существовании рецепторов к антигену еще до контакта с ним иммунной системы.
ТЕОРИИ ОБРАЗОВАНИЯ АНТИТЕЛ
В послеэрлиховский период представления об образовании антител утратили первоначальную простоту. Необходимость их пересмотра возникла в связи с тем, что химики научились синтезировать новые, отсутствующие в природе органические соединения и, как показал Ландштейнер, иммунная система оказалась способной отвечать образованием специфических антител на каждое из них. Сама возможность появления в результате естественного отбора в клетках иммунной системы тех генов, которые необходимы для образования антител ко всем этим новым, синтетическим веществам, казалась невероятной. В итоге появилась инструктивная гипотеза образования антител, согласно которой антиген, воздействуя на гибкую молекулу иммуноглобулина («инструктируя»), формирует в ней комплементарный себе центр связывания. Стремительный прогресс молекулярной биологии в 50 60-х гг. сделал инструктивную гипотезу образования антител неприемлемой, так как стало очевидным, что механизма «инструктирующего» действия антигена просто не существует. На новом витке развития научной мысли предпочтение вновь завоевали селекционные идеи. Почти одновременно Ерне и Бернетом была выдвинута клонально-селекционная теория, утверждавшая, что каждый лимфоцит образует иммуноглобулины только одной специфичности и что антиген выбирает и стимулирует клетки, несущие специфичные именно к нему антитела.
Однако еще оставался без ответа вопрос об источниках разнообразия антител. Теоретическое допущение о существовании своего особого гена для антител каждой из множества специфичностей немедленно открыло другую проблему. Половина аминокислотной последовательности любой легкой и четверть любой тяжелой цепи иммуноглобулинов всегда вариабельна, а остальная часть константна. Каким образом в случае предполагаемого множества генов антител возможно сохранение неизменной последовательности в константных областях иммуноглобулиновых цепей? На этот вопрос отвез или Драйер и Беннетт, предположив, что вариабельные и константные области кодируются отдельными генами, причем существует множество генов для вариабельных (V) и один или весьма ограниченное число генов для константных (С) областей. Теперь оставалось только объяснит ь источник многообразия вариабельных областей! Основой для этого стала идея соматического мутагенеза, согласно которой из относительно небольшого числа гаметных генов (гены зародышевой линии) в течение жизни индивида возникает множество модифицированных, т. е. подвергшихся мутациям генов. Кроме того, было высказано предположение, что полный V-ген может появляться в результате рекомбинации ряда генных сегментов. При разрезании и соединении фрагментов ДНК между ними могут встраиваться добавочные нуклеотиды, создавая дополнительную вариабельность, названную N-peгиональной, поскольку новая нуклетидная последовательность отличается от гаметной. Вместо мутаций источником разнообразия вариабельных областей могла бы служить, как предполагалось, генная конверсия с участием набора псевдогенов. В итоге было определено пять возможных источников разнообразия антигенраспознающих структур:
✵ множественность гаметных генов V-областей:
✵ соматический мутагенез;
✵ соматические рекомбинации между сегментами, образующими полный V-ген;
✵ генные конверсии;
✵ вставки добавочных нуклеотидов.
Сегодня известно, что у млекопитающих для создания разнообразия антител могут действовать все эти пят ь механизмов (рис. 8.2). Примечательно, что акулы располагают значительным числом кодирующих антитела генов и «не испытывают необходимости» в соматических рекомбинациях, тогда как у курицы число гаметных генов антител ограниченно и для этого вида характерен высокий уровень генной конверсии (см. гл. 15).
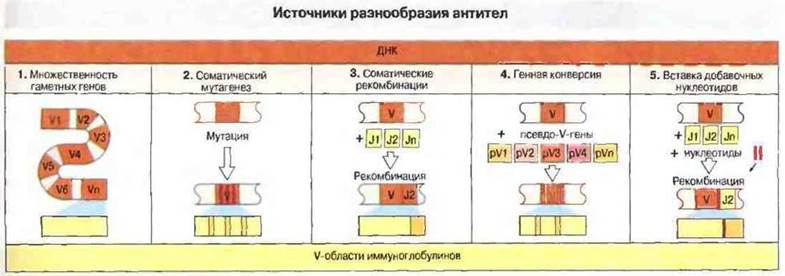
Рис. 8.2. Пять возможных источников структурного разнообразия V-областей Н- и L-цепей иммуноглобулинов.
1. Многочисленность гаметных генов. Имеется большое число отдельных неперестроенных генов (V1-Vn), каждый из которых кодирует V-домен отдельной специфичности.
2. Соматический мутагенез. В онтогенезе В-клеток в результате мутаций гаметного V-гена в разных В-клеточных клонах возникают различные V-гены.
3. Соматическая рекомбинация. В онтогенезе В-клеток происходит рекомбинация ряда генных сегментов (J1-Jn), соединяющихся с основной частью V-гена. В результате синтезируется белок, отдельные элементы которого кодируются разными генными сегментами.
4. Генная конверсия. Отрезки ДНК, принадлежащие ряду псевдо-V-генов, могут копироваться в функциональном V-гене, меняя его исходную нуклеотидную последовательность.
5. Вставка добавочных нуклеотидов. При рекомбинации перед соединением вырезанных V- и J-сегментов ДНК возможно встраивание между ними добавочных нуклеотидов, кодирующих дополнительные аминокислотные остатки V-областей.
Все эти пять механизмов служат источниками разнообразия антител у млекопитающих.