ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 8. Источники разнообразия антигенраспознающих структур
РЕГУЛЯЦИЯ СИНТЕЗА ИММУНОГЛОБУЛИНОВ
Очевидна сложность внутриклеточной регуляции синтеза иммуноглобулинов. На первой его стадии генный сегмент D сближается с сегментом J и затем к ним присоединяется сегмент V. У пре-В-клеток полностью собранные μ-цепи обнаружены в цитоплазме. Эти вновь синтезированные тяжелые цепи сохраняют связь с белками BiP/GRP78 эндоплазматического ретикулума, пока не соединятся с легкими цепями. Пре-В-клетки не образуют легких цепей, но синтезируют небольшие количества сходных с ними псевдоцепей (λ5 и VpreB у мыши, 14,1 и VpreB у человека), которые ассоциируют с μ-цепями. В результате на клеточную поверхность транспортируется небольшое количество IgM-подобных молекул, функция которых еще неясна. Известно, однако, что у мышей, неспособных синтезировать легкие псевдоцепи, существенно снижено количество В-клеток. Присутствующие в цитоплазме пре-В-клеток μ-цепи стимулируют рекомбинацию сегментов V-J в локусе к-цепей, а затем (если необходимо) и в локусе λ-цепей. Предполагается, что стимуляция рекомбинаций в указанных локусах то материнской, то отцовской хромосомы повторяется периодически, пока либо не появится функциональный рекомбинированный ген V-области легкой цепи, либо не истощится генетический материал, либо не будет устранена сама В- клетка. Если не учитывать возможные соматические мутации, рекомбинантные V-гены легких цепей в дальнейшем существенно не меняются. Однако В-клетке еще предстоит переключение Сн-генов для синтеза иммуноглобулинов разных изотипов и переход после активации на продукцию его секретируемой формы. Факты и гипотезы, касающиеся биосинтеза иммуноглобулинов, представлены на рис. 8.23.
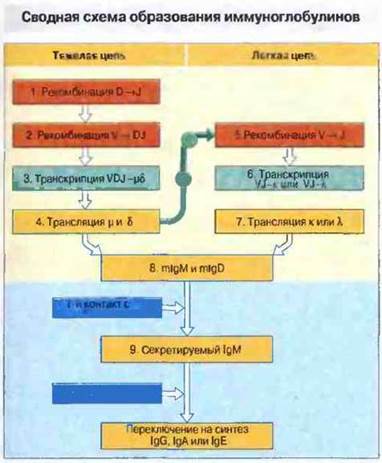
Рис. 8.23. Стадии образования иммуноглобулинов в В-клетках. В пре-В-клетках периодически инициируются попытки рекомбинации D- и J-сегментов, а затем V-сегмента для получения функционального V-D-J-гена тяжелой цепи, который транскрибируется вместе с Сμ- и Cδ-генами и транслируется в μ- и δ-цепи мембраносвязанных Ig. В свою очередь μ-цепи индуцируют рекомбинации в локусах легких цепей. Клетка пытается создать функциональные цепи, используя локусы то материнской, то отцовской хромосомы, пока не достигнет успеха или не кончатся неперестроенные наборы генных сегментов. Невозможность синтеза функционального иммуноглобулина приводит к устранению данного клона В-клеток. Зрелые В-клетки экспрессируют мембраносвязанные IgM и IgD. После первичной антигенной стимуляции эти клетки способны синтезировать секретируемый IgM; после иммунизации Т-зависимым антигеном В-клетки могут с помощью Т-клеток переключаться на синтез IgG, IgA или IgE. На этой стадии часто возникают соматические мутации V-гeнов.
Цепи иммуноглобулинов транслируются в рибосомах и переносятся через мембрану эндоплазматического ретикулума
Прежде чем начнется синтез антител, первичный транскрипт РНК должен освободиться от нитронов путем сплайсинга. В начале и в копне каждого нитрона находятся особые нуклеотидные последовательности РНК, названные донорным и акцепторным контактами. Предполагается, что они взаимодействуют друг с другом и с ядерными рибонуклеопротеинами и в результате происходит вырезание нитронов с последующей сшивкой разрыва для восстановления непрерывности цепи мРНК. Конечно, очень важно, чтобы этот процесс осуществлялся точно, без сдвига рамки считывания.
Матричная (информационная) РНК иммуноглобулиновых цепей транслируется на рибосомах, и новосинтезированные полипептиды направляются в эндоплазматический ретикулум, где легкие и тяжелые цепи образуют молекулу антитела (рис. 8.24). Мембраносвязанная и секреторная формы иммуноглобулина процессируются по-разному для правильной внутриклеточной сортировки, механизм которой пока полностью не выяснен, и достижения места назначения.
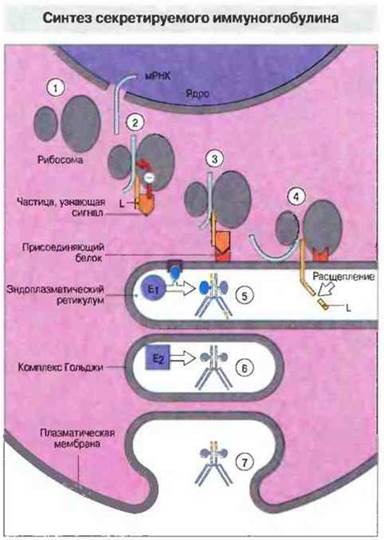
Рис. 8.24. Матричная РНК для тяжелой цепи секретируемой формы Ig покидает ядро и поступает в цитоплазму, где связывается с рибосомой (1). Транслированная на рибосоме лидерная (сигнальная) последовательность (L) синтезируемого полипептида связывается с частицей, узнающей сигнал (SRP, от англ. signal recognition particle), которая блокирует дальнейшую трансляцию (2), Комплекс SRP-рибосома мигрирует к эндоплазматическому ретикулуму (ЭР), на мембране которого SRP взаимодействует с присоединяющим белком (3). Трансляция при этом может продолжаться, и синтезируемая цепь проникает сквозь мембрану внутрь ЭР (4). Лидерная последовательность в просвете ЭР отщепляется, и готовая цепь объединяется с другими Н- и L-цепями в субъединицу молекулы иммуноглобулина (5). Группа ферментов (Е1) присоединяет к иммуноглобулину углеводы (показаны синим) и одновременно от ЭР отделяется транспортный пузырек (6). В комплексе Гольджи другая группа ферментов (Е2) модифицирует углеводные единицы иммуноглобулина, и затем полностью готовая молекула секретируется из клетки путем обратного пиноцитоза (7).