ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 8. Источники разнообразия антигенраспознающих структур
ГЕНЫ АНТИГЕНРАСПОЗНАЮЩЕГО Т-КЛЕТОЧНОГО РЕЦЕПТОРА
Структуры ТкР, связывающие антиген и молекулы МНС, кодируются четырьмя различными группами генов. Гены α- и β-цепей экспрессируются большинством периферических Т-клеток, тогда как гены γ- и δ-цепей — одной из субпопуляций тимоцитов и одной минорной популяцией периферических Т-клеток. Две пары цепей (α/β или γ/δ) в ассоциации с цепями γ, δ, ε и ζ полипептидного комплекса CD3 образуют полный ТкР (см. гл. 7).
Общий порядок расположения генов ТкР в хромосоме удивительно напоминает расположение генов тяжелых цепей иммуноглобулинов. Интересно, что δ-гены ТкР со всеми своими наборами D-, J- и С-сегментов располагаются среди α-генов (рис. 8.25).
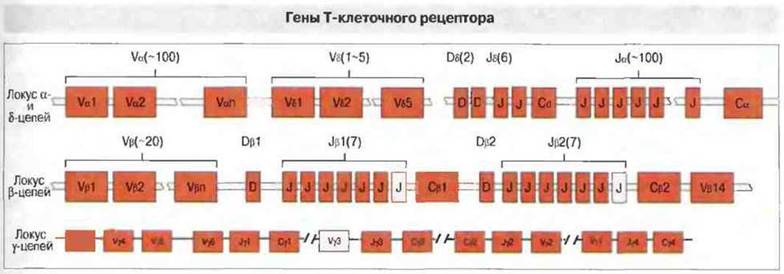
Рис. 8.25. По механизму рекомбинаций гены ТкР сходны с генами тяжелых цепей иммуноглобулинов. На рисунке представлено расположение генов в локусах α-, δ-, β- и γ-цепей Т-клеточного рецептора мыши. Гены δ-цепей размещаются среди генов α-цепей. В локусе β-цепей имеются два кластера тандемных дупликаций J-сегментов. Последние сегменты каждого Jβ-кластера и Vγ3-сегмент - это псевдогены.
Разнообразие генов ТкР создается рекомбинациями V-D-J с небольшими различиями для каждого локуса
Генные сегменты, кодирующие α-цепи, расположены на первый взгляд просто. Сложность состоит только в том, что между V- и J-локусами α-цепей находятся V-, D- и J-локусы δ-цепей. Подобно V-областям к-цепей иммуноглобулинов, V-области α-цепей ТкР кодируются геном, возникающим в результате рекомбинаций Vα- и Jα-сегментов. Степень вариабельности α-цепей существенно увеличена за счет необычайно большого числа Jα-сегментов.
Локус β включает два комплекта D-, J- и С-сегментов. Большинство Vβ-сегментов сгруппировано вместе, но один (Vβ14) удален на 3'-конец локуса. Тандемные дупликации генных Dβ-, Jβ- и Cβ-сегментов, по-видимому, возникли в довольно ранний период эволюции млекопитающих, так как они имеются как у мыши, так и у человека. Широкое разнообразие β-цепей возникает в процессе рекомбинаций, поскольку к всевозможным перестройкам V-D-J добавляются варианты сочетаний V-J и V-D-DJ. Использование всех трех рамок считывания D-сегментов дополнительно увеличивает разнообразие.
Порядок расположения генных сегментов в локусе γ-цепей у мыши и человека весьма различен. Мышиный локус удивительно напоминает локус легких цепей иммуноглобулинов он содержит четыре Сγ-гена (включая один псевдоген), каждый из которых может сочетаться с одним J-сегментом и присоединять от одного до четырех Vγ-ceгментов. В локусе γ-цепей нет ни одного D-сегмента. У человека имеется восемь Vγ-сегментов, за которыми по направлению вверх (5') следуют три Jγ-сегмента, Cγ1-ген, два добавочных Jγ-сегмента и Сγ2-ген. Существенный вклад в разнообразие γ-цепей вносят погрешности соединений V- и J-сегментов между собой и вставки добавочных нуклеотидов в местах такого соединения.
Локус δ-цепей (он был открыт при изучении α-локуса) организован довольно просто и содержит только пять Vδ-, два Dδ- и шесть Jδ-cегментов, однако расчетное число вариантов δ-цепей, которые могут появляться в результате неточности соединения, вставок добавочных нуклеотидов и использования всех трех рамок считывания D-сегментов, достигает 1014.
Гены ТкР имеют, по-видимому, одинаковый с В-клеточными Ig-генами механизм рекомбинаций, поскольку фланкирующие сигнальные последовательности рекомбинации (гептамер — 12- или 23-нуклеотидный спейсер — наномер) тех и других сходны. И в В-, и и Т-клетках действуют подобные или идентичные ферменты рекомбинации; генные Dβ- и Jβ-сегменты ТкР, экспериментально трансфицированные в В-клетки, могут и в них рекомбинировать адекватным образом. Существенный вклад в разнообразие ТкР вносит заметно развитая «N-региональная» вариабельность.
Такой важный источник разнообразия иммуноглобулинов, как соматический мутагенез, не оставляет последствий в генах ТкР. Вероятно, это обусловлено тем, что Т-клетки должны распознавать продукты МНС и поддерживать толерантность к собственным антигенам организма.