ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 9. Распознавание антигена
ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА
Презентации антигенов Т-клеткам предшествует процессинг
Циркулирующие антитела и реакции клеточного иммунитета, как правило, специфичны в отношении разных детерминант одного и того же антигена. Например, у мыши В-клетки распознают аминоконцевые эпитопы глюкагона, тогда как Т-клетки — его карбоксиконцевые детерминанты (рис. 9.18). Это происходит благодаря тому, что презентируются не интактные молекулы антигена, а их фрагменты (продукты расщепления, или процессинга — переработки) в ассоциации с продуктами МНС на клеточной поверхности. Клетки, процессирующие антиген для презентации — это либо специализированные ангигенпрезентируюшие клетки (АПК), способные стимулировать пролиферацию Т-клеток, либо инфицированные вирусами клетки организма, которые затем становятся мишенями для Тц.
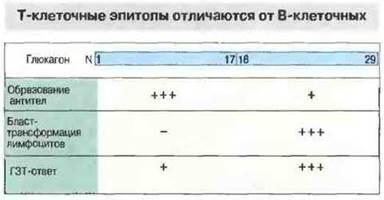
Рис. 9.18. Различные формы иммунного ответа при иммунизации двумя пептидами, из которых состоит молекула глюкагона. Антитела (В-клеточный ответ) направлены преимущественно к эпитопам N-концевого пептида, тогда как С-концевой пептид (позиции 18-29) стимулирует Т-клеточный ответ, в частности бласттрансформацию лимфоцитов in vitro и гиперчувствительность замедленного типа (ГЗТ) in vivo.
Процессинг антигена заключается в его расщеплении на пептидные фрагменты. Подавляющее большинство эпитопов, распознаваемых Т-клетками, представляет собой фрагменты пептидной цепи, часто недоступные для иммунного распознавания в составе молекул интактного белка. Только малая часть пептидных фрагментов белкового антигена способна связаться с соответствующей молекулой МНС. Более того, разные молекулы МНС связывают различные наборы пептидов. Например, Тх-клетки мышей разных гаплотипов (т. е. носителей различных молекул МНС) распознают разные пет иды вирусного антигена (рис. 9.19). Эти различия распознавания обусловлены главным образом способностью данного пептида связываться с определенной молекулой МНС класса II.
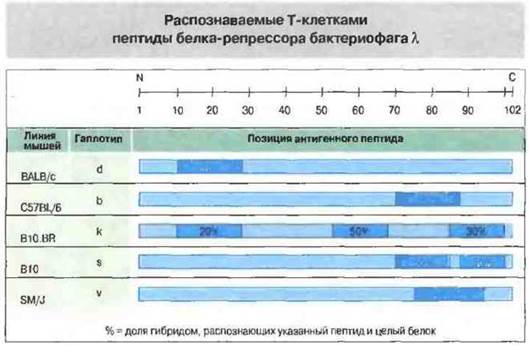
Рис. 9.19. Различия между пептидными фрагментами одного и того же белкового антигена, распознаваемыми Т-клетками мышей разных гаплотипов (т. е. носителями разных молекул МНС класса II). Животных иммунизировали белком-репрессором бактериофага X, получали Т-клеточные гибридомы и определяли их способность распознавать набор перекрывающихся пептидных фрагментов этого белка. Позиции антигенных пептидов отмечены темно-голубым цветом. Один из пептидов всегда иммунодоминантен, хотя для некоторых линий мышей антигенными были несколько пептидов. (По Roy S. etal., 1989).
Перед связыванием с молекулами МНС белковые антигены расщепляются на пептиды. Процессинг антигенов, в результате которого образуются пептиды, способные связаться с молекулами МНС, происходит во внутриклеточных органеллах антигенпрезентируюших клеток (рис. 9.20). Для целей исследования процесс расщепления антигена можно исключить, использовав в качестве антигенов синтетические пептиды. Применение именно таких легкосинтезируемых пептидов с известной аминокислотной последовательностью позволило идентифицировать эпитопы, распознаваемые Т-клетками разной специфичности (рис. 9.21). Аминокислотные замены в различных позициях дали возможность выяснить сравнительное значение того или иного аминокислотного остатка в составе определенных эпитопов. Кроме тог о, с помощью пептидов известной структуры была доказана способность молекул МНС классов I и II к прямому связыванию фрагментов антигена. Путем сравнения эффектов аминокислотных замен, т. с. их влияния на МНС-связывание и Т-клеточную реактивность, удалось установить, какие именно аминокислотные остатки контактируют с молекулой МНС и с Т-клеточным рецептором. Например, фрагмент, состоящий из остатков 52—61 полипептидной цепи лизоцима яичного белка, распознают Н-2-IАк-рестриктированныс Т-клетки. Этот пептид связывается с молекулами IAk, и, как установлено, в его взаимодействии с продуктами МНС класса II принимают участие три аминокислотных остатка, в то время как три других остатка осуществляют контакт с ТкР.
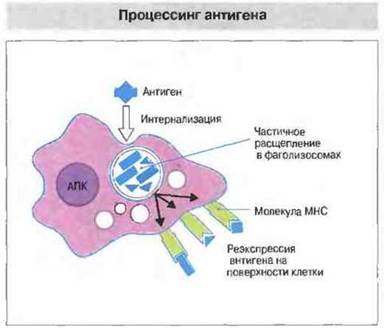
Рис. 9.20. Экзогенные антигены захватываются антигенпрезептирующими клетками, а затем расщепляются их протеолитическими фрагментами в специализированных внутриклеточных компартментах. Антигенные пептиды образуют комплекс с молекулами МНС класса II в везикулах, которые, направляясь к поверхности клетки, движутся навстречу эндоцитарным везикулам.
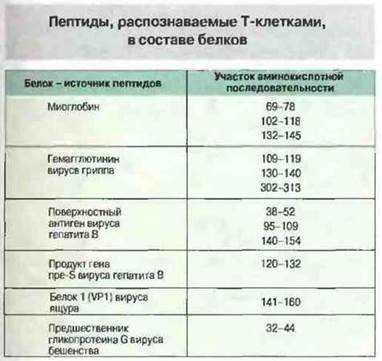
Рис. 9.21. В таблице представлены распознаваемые Т-клетками пептиды ряда белков. Чтобы активировать Т-клетки, можно использовать синтетические пептиды, повторяющие полную аминокислотную последовательность соответствующего белка, но часто для активации достаточно воздействия лишь одного или нескольких отдельных фрагментов его цепи.