ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 15. Эволюция иммунной системы
■ У беспозвоночных важную роль в ликвидации чужеродного материала выполняют фагоцитоз и инкапсулирование; у многих видов эти процессы осуществляются белыми клетками крови.
■ Установлена возможность распознавания чужеродных трансплантатов у животных, появившихся уже на ранних этапах эволюции, но специфичность реакции и клональная экспансия клеток у беспозвоночных недостаточно доказаны.
■ В процессах распознавания «свое/не-свое» участвуют лектины и профенолоксидаза.
■ У многих беспозвоночных в иммунорегуляции участвуют цитокин-подобные молекулы.
■ Иммуноглобулинами беспозвоночные не обладают, хотя у многих из них обнаружены IgG-подобные молекулы. У некоторых вторичнополостных наблюдаются индуцируемые гуморальные иммунные реакции широкого спектра.
■ У всех челюстноротых позвоночных имеются В-клетки и IgM. У пойкилотермных животных, несмотря на присутствие дополнительных тяжелых цепей, не относящихся к μ-цепям, аффинность антител остается низкой. Метод рекомбинантной ДНК позволяет обнаружить у позвоночных разных уровней филогенеза несколько типов организации генов Ig.
■ У хрящевых рыб идентифицирован главный комплекс гистосовместимости (МНС). Начиная с костных рыб, появляются Т-клетки; рецепторные молекулы этих клеток в настоящее времяизучаются.
■ К элементам системы врожденного иммунитета у позвоночных относятся нормальные клетки-киллеры (НК), фагоциты, белки комплемента и иммунорегуляторные цитокины.
Изучение разнообразных видов позвоночных позволяет составить представление об эволюционном развитии иммунной системы вплоть до формирования ее сложноорганизованных механизмов у млекопитающих. Однако филогенез системы приобретенного иммунитета позвоночных, особенно его молекулярных основ, остается неясным, несмотря на многочисленные исследования иммунитета у беспозвоночных. В то же время изучение беспозвоночных проливает свет на происхождение врожденного иммунитета (например, фагоцитоза), полностью сформированного уже у позвоночных. Поскольку' беспозвоночные чрезвычайно многочисленны и разнообразны — одиночные и колониальные формы, полостные и бесполостные, обладающие и не обладающие кровеносной системой, всего более 95 % всех видов животных на Земле — среди них можно найти много удобных объектов для экспериментальных исследований.
На рис. 15.1 упрощенно представлено эволюционное древо царства животных с разделением от вторичнополостных на две основные ветви, принципиально различающиеся стадиями эмбриогенеза. Одна ветвь, ведущая к первичноротым — моллюскам, кольчатым червям и членистоногим, - дивергировала на этом раннем этапе эволюции от пути, ведущего к появлению вторичноротых — иглокожих, оболочников и позвоночных. Исследования в области иммунитета у беспозвоночных проводятся в основном на членистоногих и моллюсках, в связи с тем что многие из них являются переносчиками возбудителей болезней или вредителями сельскохозяйственных культур. Поэтому филогенетически более близкие к позвоночным таксоны (например, оболочники и иглокожие) привлекают гораздо меньше внимания. Кроме того, поскольку все предки позвоночных — вымершие виды, выводы о происхождении иммунитета у позвоночных, сделанные на основании изучения беспозвоночных (например, оболочников) (рис. 15.2), могут быть лишь гипотетическими, основанными на предположении о близком родстве некоторых современных животных с предковыми позвоночными. На рис. 15.1 перечислены также обнаруженные у беспозвоночных реакции клеточного и гуморального иммунитета.
На рис. 15.3 представлены вероятные этапы эволюции клеток крови и иммунной системы у позвоночных. Хотя иммуноциты имеются и у беспозвоночных, только позвоночные обладают лимфоцитами с высокой специфичностью и среди них клетками иммунологической памяти. Какие же факторы внешней среды могли обусловить усложнение иммунной системы у позвоночных? Возможно, на уровне столь высокоорганизованных долгоживущих животных возросла угроза рака и вирусных инфекций и влияние этих факторов способствовало формированию высокочувствительной иммунной системы на основе эффекторных клеток крови, способных распознавать чужеродные пептиды в ассоциации с гликопротеинами главного комплекса гистосовместимости (МНС) на поверхности инфицированных или мутантных клеток.
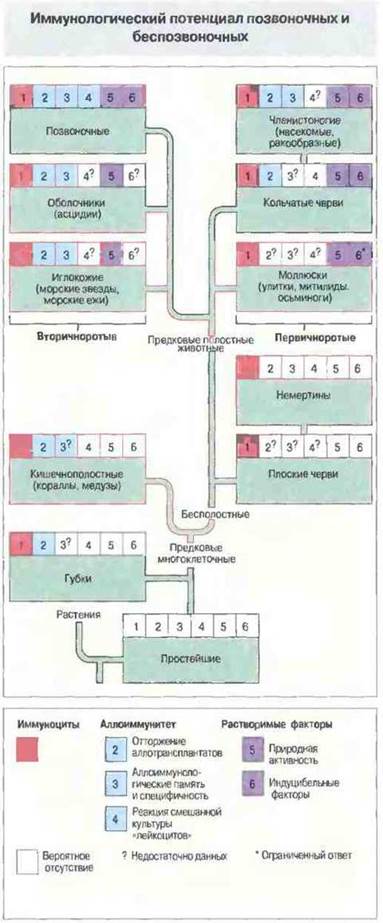
Рис. 15.1. Некоторые сведения о клеточных и гуморальных иммунных реакциях у различных беспозвоночных и позвоночных.

Рис. 15.2. Две конкурирующие за пространство колонии асцидий (оболочники) Clavelina lepadiformis (С) и Morchellium argus (М). В центре (указано стрелкой) виден сифон нижележащей асцидии. Диаметр отдельной особи Clavelina приблизительно 4 мм. (Фото любезно предоставлено д-ром Р. Dyrynda.)
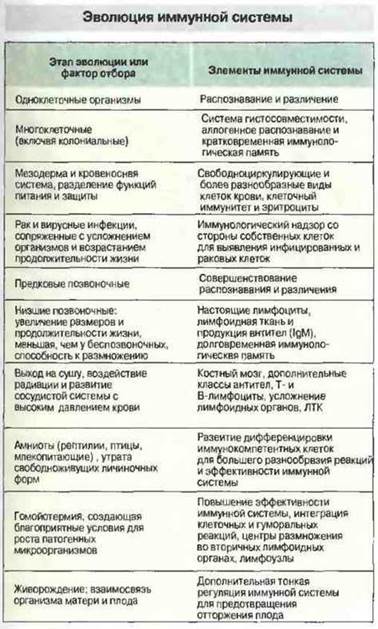
Рис. 15.3. Этапы эволюции, возможно важные в филогенезе клеток крови и иммунной системы. (С разрешения по Rowley A.F., Ratcliffe N.A., eds. Vertebrate Blood Cells. Cambridge: Cambridge University Press, 1988.)
ЛТК - лимфоидная ткань в слизистой кишечника.
ИММУНИТЕТ У БЕСПОЗВОНОЧНЫХ
Классификация клеток крови у беспозвоночных
У большинства беспозвоночных имеются белые кровяные клетки (лейкоциты), но красные клетки (эритрошпы), как правило, отсутствуют. Лейкоциты могут либо быть зафиксированными (неподвижными), либо свободно мигрировать по кровеносным сосудам, либо заселять заполненные жидкостью полости тела, называемые целомом (целомоциты) или гемоцеломом (гемоциты).
Первые клетки крови образовались, вероятно, из свободноживуших преяковых клеток, сходных с простейшими. У примитивных многоклеточных, например у губок, кишечнополостных и плоских червей, блуждающие фагоцитарные амебоциты не только выполняют функцию защиты, но и участвуют в процессах питания и экскреции. На уровне вторичнополостных (первично- и вторичноротых), тела которых крупнее и более сложно устроены, возникла необходимость в циркуляторной системе для переноса питательных веществ и отходов внутри организма. Амебоподобные клетки, больше не нужные для собирания пиши, вероятно, мигрировали из соединительной ткани в эту циркуляторную систему. Здесь из них образовалось множество типов клеток, часть которых приобрела специфическую роль в иммунореактивности (рис. 15.4).
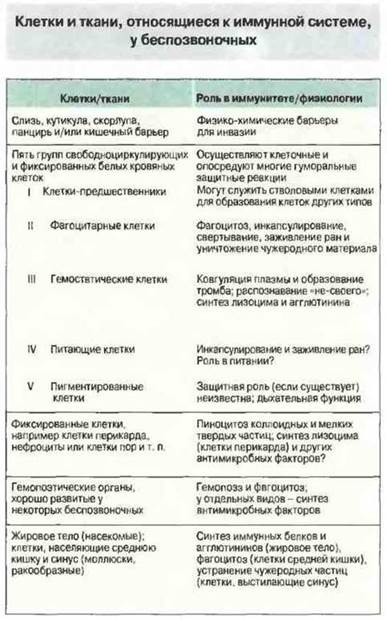
Рис. 15.4. Иммунная система у беспозвоночных. (С разрешения по Ratcliffe N.A. 1985. Immunol. Lett. 10, 253-270; Elsevier Science Publications.)
Из-за огромного разнообразия беспозвоночных их свободные лейкоциты невозможно классифицировать, в отличие от лейкоцитов у позвоночных, только по окрашиваемости и морфологии. Однако по функциональным критериям удается выделить пять основных групп таких клеток (см. рис. 15.4).
✵ Клетки-предшественники, которые наряду с имеющейся в том или ином количестве гемопоэтической тканью могут выполнять роль стволовых клеток. По строению они напоминают лимфоциты позвоночных (рис. 15.5), но данных о действительной гомологии очень мало.
✵ Фагоцитарные клетки (рис. 15.6), вероятно, представляют собой единственный тип клеток, присутствующий у всех представителей царства животных. Они соответствуют гранулоцитам или макрофагам млекопитающих, но несут иные поверхностные маркеры.
✵ Гемостатические (гранулярные) клетки принимают участие в коагуляции и заживлении ран и важны как эффекторные для распознавания «не-своего».
✵ Питающие клетки присутствуют лишь у некоторых видов.
✵ Пигментные клетки имеются у многих видов, но только у некоторых они содержат дыхательный пигмент, напоминая тем самым эритроциты позвоночных.
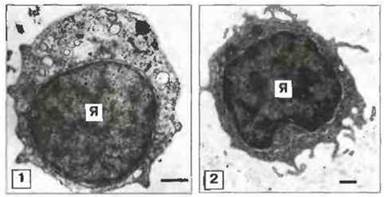
Рис. 15.5. Электронные микрофотографии лимфоцит-подобной клетки асцидии Сionа intestinalis (1) и лимфоцита рыбы Blennius pholis (морская собачка) (2). Обратите внимание на сходство строения этих клеток: обе содержат крупное ядро (Я) и тонкий слой недифференцированной цитоплазмы. Шкала 0,5 мкм. (Фото любезно предоставлены д-ром A.F. Rowley; с разрешения из Endeavour (New Series) 13, 72-77, © Maxwell Pergamon Macmillan pic, 1989.)
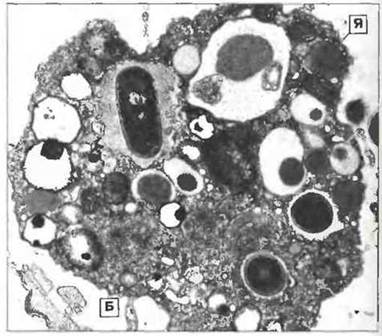
Рис. 15.6. Электронная микрофотография фагоцитарной клетки оболочника Сionа intestinalis. Внутри клетки видны поглощенные бактерии (Б). (Я - ядро.) Шкала 0,5 мкм. (Фото любезно предоставлено д-ром A.F. Rowley.)
Беспозвоночные лишены лимфоцитов и антител, но тем не менее обладают эффективными механизмами защиты
Иммунные системы беспозвоночных, очевидно, не включают в качестве компонентов ни иммуноглобулинов, ни взаимодействующих субпопуляций лимфоцитов, ни лимфоидных органов. Тем не менее само существование огромного числа и разнообразия беспозвоночных свидетельствует о наличии у них эффективных систем защиты собственного организма.
Как и у позвоночных, первой линией обороны у них служат эффективные физико-химические барьеры (рис. 15.4). Слизь, покрывающая тело у многих кишечнополостных, кольчатых червей, моллюсков и некоторых оболочников, задерживает и уничтожает потенциально патогенные микроорганизмы (рис. 15.7). Твердый наружный скелет, такой как панцирь или скорлупа, у некоторых кишечнополостных и моллюсков, иглокожих и насекомых также препятствует инвазии в организм чужеродного материала.
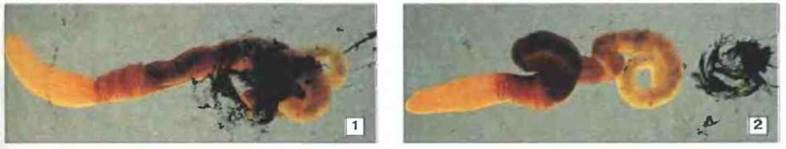
Рис. 15.7. Слизистый слой, окружающий тело желудевого червя Saccoglossus ruber, обволакивает и удаляет чужеродные частицы. Червя помещали на 2-3 мин в суспензию угля в морской воде, а затем переносили в чистую морскую воду. Через 12 мин в слизистом слое, окутывающем его тело, все еще присутствует большое количество угля (1). Через 15 мин уголь полностью удален и лежит в отдельном шарике слизи (2). С попавшими в слизь микроорганизмами дело, по-видимому, обстоит точно так же. Желудевые черви принадлежат к группе «высших беспозвоночных», родственных оболочникам. Шкала 5 мм (Фото любезно предоставлены д-ром DA. Millar.)
При нарушении целостности этих барьеров против проникшего в организм чужеродного тела действует целый ряд взаимосвязанных клеточных и гуморальных защитных механизмов, к которым относятся следующие.
✵ Свертывание/коагуляция крови и заживление ран.
✵ Фагоцитоз.
✵ Инкапсулирование.
✵ Действие врожденных и индуцибельных антимикробных факторов.
Эти механизмы предполагают распознавание «не-своего» и присутствие рецепторных молекул в крови и на поверхности клеток крови.
Раны быстро закрываются благодаря коагуляции жидкостей организма, вызываемой гемостатическими клетками и компонентами плазмы
Раны, образующиеся у беспозвоночных при травме или инвазии паразитов, быстро заживают, что предотвращает смертельную потерю жидкостей организма. Раны закрываются вследствие выпячивания жирового тела или кишки, сокращения мышц, коагуляции плазмы, миграции клеток крови к месту поражения и их агрегации с образованием тромба и/или за счет отложения меланина. Миграцию лейкоцитов к ране стимулируют, вероятно, цитокин подобные факторы (см. ниже).
Тромбирование раны с коагуляцией плазмы наблюдается главным образом у членистоногих, хотя получены данные об аналогичном процессе у кольчатых червей и иглокожих. В коагуляции участвуют гемостатические клетки, которые агрегируют в месте повреждения и выделяют свое содержимое, вызывающее свертывание плазмы и образование прочной гемоцитарной «пробки». У многих видов в этом процессе участвуют и компоненты плазмы. Как и у млекопитающих, сама поврежденная ткань, микробные компоненты, а также изменения концентрации Са2+ или pH индуцируют сложный каскад ферментативных реакций в месте повреждения. Это напоминает альтернативный путь активации комплемента. Система настолько чувствительна, что у мечехвостов, например, она активируется эндотоксином Escherichia coli в дозе всего 4 нг/мл. Процесс коагуляции чрезвычайно важен, поскольку обеспечивает высокочувствительное распознавание «чужого» благодаря дегрануляции гемостатических клеток. В коагуляции может принимать участие фермент профенолоксидаза (ПФО): превращаясь под действием каскада сериновых протеаз в фенолоксндазу (ФО), она способствует образованию медиаторов более поздних стадий иммунного ответа (рис. 15.8). Недавно установлено, что каскад ПФО существует и у других беспозвоночных, таких как круглые черви и оболочники.
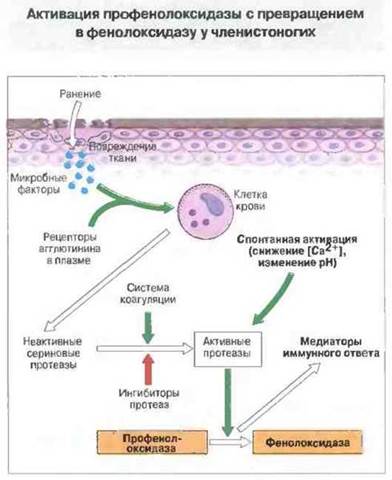
Рис. 15.8. Вероятная схема активации профенолоксидазы (ПФО) с образованием фенолоксидазы (ФО) у членистоногих Процесс стимулируется местным повреждением ткани, микроорганизмами и изменениями концентрации Са2+ и pH и может привести к коагуляции плазмы и появлению факторов, опосредующих более поздние иммунные реакции (В основном по результатам исследований, проведенных д-рами К. Soderhall, М. Ashida и N. Radcliffe.)
Внедрившихся микробов поглощают фагоцитарные клетки, более крупные агенты окружаются многоклеточными капсулами
Фагоцитарные клетки присутствуют у всех беспозвоночных и вместе с врожденными гуморальными факторами (см. ниже) образуют первую линию защиты от микроорганизмов (см. рис. 15.6). Как и у позвоночных, здесь наблюдаются все фазы их действия: хемотаксис, прикрепление, поглощение и уничтожение. Однако распознавание мишени опосредуют не Fc-рецепторы, и лишь у одного вида на поверхности фагоцитов обнаружены С3b-подобные рецепторы. Фагоцитоз, как и у позвоночных, может происходить без участия опсонизирующих факторов. Однако у моллюсков, членистоногих и оболочников в присутствии лектинов плазмы и компонентов профенолоксидазного каскада он усиливается.
Если внедрившиеся патогенные микробы слишком крупны или многочисленны, вокруг них образуются многоклеточные агрегаты, называемые узелками или капсулами и напоминающие гранулемы у млекопитающих (рис. 15.9). Секвестрированные таким образом микроорганизмы предположительно погибают под действием лизосомных ферментов и лизоцима, содержащихся в лейкоцитах, а также под действием пероксидазы и активных форм кислорода (обнаруженных у некоторых круглых червей, моллюсков и членистоногих).
И фагоцитоз, и инкапсулирование зависят от кооперации гемостатических и фагоцитарных клеток (см. ниже).
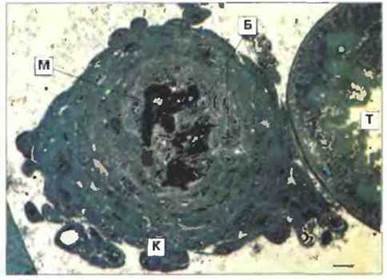
Рис. 15.9. Инкапсулирование бактерий кровяными клетками гусеницы. На последней стадии развития личинки бабочки Pieris brassicae в нее вводили бактерии (убитые нагреванием клетки Bacillus cereus). Через 24 ч образовавшуюся вокруг бактерий капсулу удаляли и делали ее срезы. Обратите внимание на темную пигментированную центральную область, состоящую из меланина (М), многослойный покров из клеток крови (К), палочковидные бактерии (Б) и прикрепление капсулы к мальпигиевой трубочке (Т). Шкала 10 мкм.
Жидкости организма содержат у беспозвоночных ряд врожденных и индуцибельных гуморальных защитных факторов
Врожденные защитные факторы. У беспозвоночных не обнаружено иммуноглобулинов, но жидкости организма содержат ряд защитных гуморальных факторов - агглютинины, лизоцим и другие лизины, иные антимикробные соединения, лизосомные ферменты и обездвиживающие факторы. Имеются также данные, указывающие на присутствие компонентов комплемент-подобной системы. Например, у морских ежей на фагоцитах могут присутствовать С3b-подобные рецепторы и обнаружена гуморальная литическая система, сходная с системой комплемента. Кроме того, установлено, что кровь гусениц реагирует с одним из факторов яда кобры (С3b кобры) и при этом появляется С3-конвертазная активность. Обладающий ею фермент расщепляет молекулы С3 быка с образованием молекул, подобных С3b. Профенолоксидазный каскад у членистоногих сравним с альтернативным путем активации системы комплемента, поскольку оба они стимулируются непосредственно компонентами микробных клеток и включают ряд последовательно активируемых протеаз (рис. 15.8). Для того чтобы подтвердить наличие у беспозвоночных альтернативных путей активации комплемента, необходимы детальные исследования на молекулярном уровне.
Индуцибельные гуморальные защитные факторы. Известно, что у беспозвоночных содержание агглютининов и гемолизинов иногда возрастает, однако подробно индуцибельные антимикробные факторы исследованы только у насекомых. Предполагается присутствие этих факторов и у некоторых других беспозвоночных, но для выявления их у большего числа организмов и для описания свойств необходимо подбирать подходящие иммуногены и/или проводить эксперименты по иммунизации. У таких насекомых, как мухи, бабочки и пчелы, через несколько часов после введения антигена может появляться до 15 антибактериальных белков (рис. 15.10). Многие из этих пептидов очищены и секвенированы; они обладают активностью широкого спектра, но сохраняют ее лишь в течение нескольких суток, чем резко отличаются от иммуноглобулинов позвоночных. Недавно аналогичные антибактериальные белки были обнаружены и у некоторых позвоночных (см. стр. 293). По-видимому, они представляют собой древние, но все еще важные факторы иммунитета. Один из таких факторов, цекропин, называемый также Р4 или гемолином (рис. 15.10), гомологичен (38 %) некоторым доменам иммуноглобулинов. Возможно, гемолин — это примитивная форма иммуноглобулина, но не исключено и его независимое развитие у беспозвоночных. У таракана американского обнаружен другой вид индуцибельного белка, гораздо более сходный с иммуноглобулинами позвоночных. Он обладает молекулярной массой 700 кДа, высокоспецифичен и сохраняет активность в течение недель, а не суток. Для сопоставления его с иммуноглобулинами позвоночных требуются исследования на молекулярном уровне.
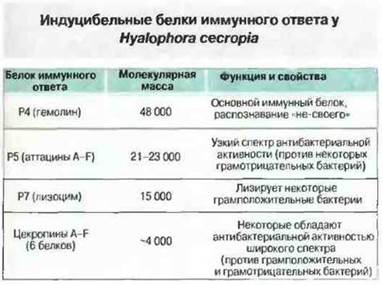
Рис. 15.10. Индуцибельные иммунные белки у моли Hyalophora cecropia, выделенные из гемолимфы через 10 ч после иммунизации бактериями (Enterobacter cloacae). (По Н. Boman, D. Hultmark и др.)
В распознавании «не-своего» и в межклеточной кооперации участвуют разнообразные факторы
Беспозвоночные обладают способностью распознавания, часто с весьма высокой специфичностью, разнообразною чужеродного материала. Это распознавание осуществляют присутствующие в жидких средах организма молекулы, к которым относятся агглютинины, компоненты профснолоксидазного каскада, а также цекропин насекомых (гемолин).
Агглютинины. Эти белки, выделенные из гемолимфы моллюсков и насекомых и из крови оболочников, усиливают распознавание тест-частиц in vitro и их выведение из циркуляции in vivo. Агглютинины присутствуют также на поверхности клеток крови и служат молекулами-мостиками, соединяющими лейкоциты с чужеродными частицами, как и в иммунной системе млекопитающих.
Система профенолоксидазы (ПФО). У членистоногих и эта система служит, вероятно, источником распознающих факторов. При превращении ПФО в фюнолоксилазу (ФО) (см. рис. 15.8) из гемостатических клеток высвобождаются факторы распознавания, которые усиливают фагоцитоз и инкапсулирование чужеродных агентов. Неданно было установлено, что очищенный агглютинин из крови таракана, обозначенный BDLI, активирует каскад ПФО; этот факт позволяет объединить представления о функции двух систем распознавания. Белок BDLI обладает также высоким структурным и функциональным сходством с маннозоевнзываюшмми лектинами (МСЛ) позвоночных. (МСЛ представляют собой необходимые компоненты неспецифического иммунитета позвоночных, поскольку они способны связываться с поверхностью проникших в организм микробов и активировать системы комплемента.) Подобно МСЛ, белок BDLI активирует комплемент и содержит коллаген- и углевод-распознающие домены.
Гемолин. Кроме агглютининов и ПФО, в гемолимфе насекомых присутствует и иммунный белок, называемый гемолином. В нем различают четыре иммуноглобулин-подобных домена, которые связываются с поверхностью бактерий и могут участвовать в распознавании чужеродных молекул.
Процесс распознавания «не-своего» с последующим фагоцитозом предполагает кооперацию между гемостатическими и фагоцитарными клетками (рис. 15.11). Таким образом, хотя беспозвоночные лишены взаимодействующих субпопуляций антигенпрезентирующих клеток и лимфоцитов, клеточный иммунитет и у них осуществляется путем кооперации разнообразных мммуноцитов.
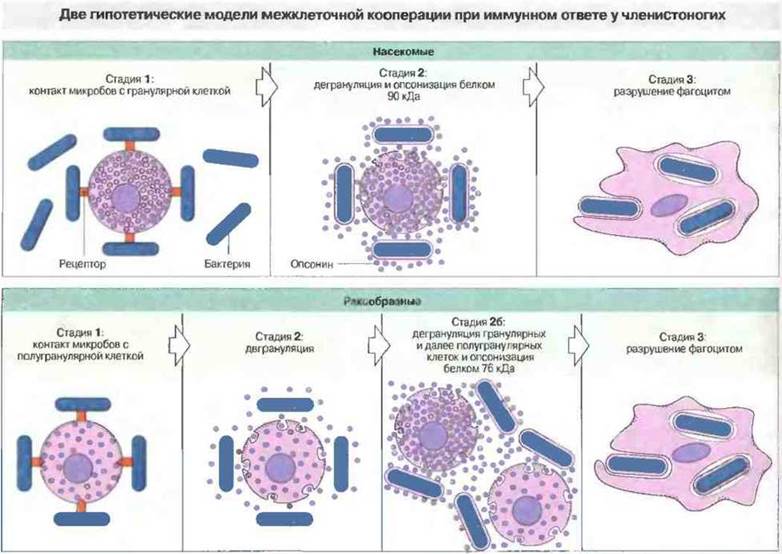
Рис. 15.11. Схемы, основанные на результатах изучения реакций очищенных популяций клеток крови на тест-частицы. У насекомых распознавание «не-своего» (стадии 1 и 2) осуществляются гранулярными (гемостатическими) клетками, а уничтожение (стадия 3) - фагоцитами. Согласно модели, у ракообразных на стадии 26 происходит резкое усиление реакции за счет взаимодействия полугранулярных и гранулярных клеток. Белки 90 и 76 «Да, обнаруженные у насекомых и ракообразных соответственно, представляют собой опсонизирующие (распознающие) молекулы, образующиеся при активации профенолоксидазного каскада. (По Radcliffe N. In: Warr G.W., Cohen N., eds. Phylogenesis of Immune Functions. Oxford: CRC Press, 1991: 62. Данные по ракообразным - из работы проф. К. Soderhall и др.)
У видов - переносчиков возбудителей болезней клеточные и гуморальные защитные механизмы определяют устойчивость к паразитарным инвазиям
Все больше данных указывает на то, что способность некоторых беспозвоночных (например, комаров, мух цеце, мошек, москитов, клешей и легочной улитки) передавать болезни (малярию, сонную болезнь, кожный лейшманиоз, онхоцеркоз, болезнь Чагаса и шистосомоз) существенно зависит от их иммунореактивности. Так, установлено, что иммунный ответ в форме инкапсулирования у москитов и легочной улитки обусловливает эффективную изоляцию и, возможно, гибель инкапсулированных паразитов. Кроме того, в недавно проведенных исследованиях у москитов, мух цеце, мошек и клещей обнаружены кишечные агглютинины — возможно, главные факторы, влияющие на размножение и выживание в этих насекомых простейших-паразитов, поглощаемых с кровью. У некоторых мух цеце (Glossina), в частности, ингибирование агглютинина средней кишки D-глюкозамином значительно ускоряет их инфицирование Trypanosoma bruceiи T.rhodesiense. Важную роль может играть и профенолокендаза; в слюнных железах самок мухи цеце Glossina morsitans morsitans, уровень ПФО у которых гораздо выше, чем у самцов, присутствует намного меньше зрелых форм Т. b. rhodesiense. Установлено также, что индуцибельные антибактериальные пептиды, присутствующие в крови мошек Simulium, убивают личинок паразитических нематод. Многое еще предстоит изучить в этой новой и увлекательной области исследований, в том числе способы преодоления паразитами защитных иммунных реакций в организме переносчиков.
Защитные реакции у беспозвоночных регулирует набор цитокинов, частью сходных с интерлейкинами позвоночных
Обнаруженные у беспозвоночных цитокин-подобные молекулы, возможно, взаимосвязанно регулируют защитные реакции, подобно тому как это происходит у позвоночных. Присутствие родственных цитокинам молекул уже у простейших указывает на то, что они имеются у всех представителей животного царства. Например, феромон простейших, Еr-1, структурно и функционально сходен с интерлейкином 2 (ИЛ-2). Кроме того, у кольчатых червей, иглокожих и оболочников недавно были выделены соединения, обладающие ИЛ-1α-, ИЛ-1β- и ФHО (фактор некроза опухолей)-подобной активностью. Первые две активности определяли с помощью тест-системы для позвоночных (по пролиферации мышиных тимоцитов), причем эти активности ингибировала поликлональная антисыворотка к ИЛ-1 позвоночных. Цитокин ИЛ-1 беспозвоночных стимулирует агрегацию, фагоцитарную активность и пролиферацию клеток «крови» этих животных. ФНО-подобную активность у беспозвоночных выявляли по цитотоксическому действию на линию клеток L929; этот метод также обычно используется для определения ФHО позвоночных.
У беспозвоночных найдены и другие соединения, обладающие цитокин-подобной активностью. Это, например, фактор, снижающий число плазмонитов (лейкоцитарного типа), активатор лейкоцитов (гемокипин) и различные стимуляторы инкапсулирования и фагоцитоза у насекомых. Фактор, продуцируемый лейкоцитами иглокожих (фактор морской звезды), оказывает митогенное действие на лимфоциты млекопитающих и индуцирует скопление белых кровяных клеток у морских звезд. Кроме того, у оболочников обнаружен провоспалительный цитокин, влияющий на продукцию антител, фагоцитоз и клеточно-опосредованную цитотоксичность у позвоночных, а также на фагоцитарную активность лейкоцитов у креветок. Предстоит дальнейшее изучение этих молекул. Наконец, следует упомянуть и о том, что в иммунореактивности юз его к крови беспозвоночных, как установлено, участвуют эйкозаноиды и опиатные пептиды.
Многие беспозвоночные обладают способностью к отторжению алло- и ксеногенных трансплантатов
Для иммунитета позвоночных характерна усиленная и высокоспецифичeская реакция (память) на повторное воздействие антигена. Ее осуществление зависит от лимфоцитов и главного комплекса гистоcовмeстимости (МНС). Степень специфичности и запоминание антигена у беспозвоночных были изучены в опытах по трансплантации и имплантации с определением цитотоксичности. Из-за жесткости наружного скелета или мягкости наружных покровов беспозвоночным трудно производить пересадку тканей. Нелегко и решить, произошло ли отторжение. Несмотря па эти трудности, удалось установить, что у большинства беспозвоночных происходит разрушение ксенотрансплантатов, а губки, кишечнополостные, аннелиды, насекомые, иглокожие и оболочники обладают и способностью распознавания аллогенного материала (рис. 15.11, 15.12 и 15.13). Кажущееся отсутствие отторжения аллогенной ткани у моллюсков связано, вероятно, с техническими трудностями пересадки. Однако не у всех групп беспозвоночных, способных отторгать аллотрансплантаты, эти реакции характеризуются специфичностью и памятью; обычно специфичность их крайне невысока, а память недолговременна (см. рис. 15.1). Значительные расхождения результатов трансплантации могут быть связаны с тем, что некоторые исследователи не учитывали зависимость процесса отторжения от температуры.
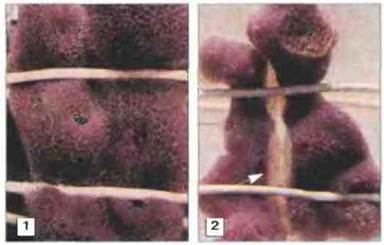
Рис. 15.12. Клеточный иммунитет у губок: аллогенная несовместимость и изогенная совместимость. У двух интактных тел губок (Callyspongia spp.) одной и той же колонии и двух от разных колоний создавали общую систему циркуляции, скрепляя их друг с другом виниловыми проводками. 1. Сосуществование изогенных парабионтов (из одной колонии) сохранялось неопределенно долго, х 0,5 2. Несовместимость аллогенных парабионтов (из разных колоний) через 7-9 сут (24 -27 С) проявлялась цитотоксическими реакциями и некрозом (указан стрелкой), х 0,25. (Фото любезно предоставлены д-ром W. Hildemann.)
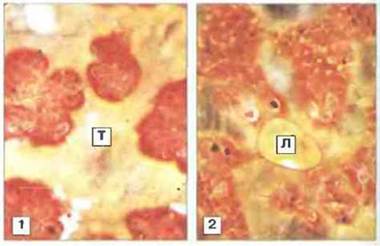
Рис. 15.13. Трансплантационный иммунитет у иглокожих (Dermasterias). 1. Несмотря на технические трудности, этот аутотрансплантат (Т) сохранился в прекрасном состоянии через 300 сут после подсадки. 2. Аллотрансплантат (Л), отторгшийся на 286 сут (14-15 °С), выглядит бледным и сморщенным. В отторжении принимают участие лимфоцит-подобные клетки и более крупные фагоцитарные клетки, х 4 (Фото любезно предоставлены д-ром W. Hildemann.)
Наличие аллогенного распознавания у колониальных беспозвоночных, таких как губки, кишечнополостные и оболочники, неудивительно, так как целостности колоний постоянно угрожает разрастание соседних колоний (рис. 15.2). Эксперименты с личинками колониальных оболочников показали, что аллогенное распознавание и оплодотворение контролирует один генный локус с множественными аллелями. Таким образом, имеется определенное сходство между системой, существующей у оболочников, и генами гистосовместимости у млекопитающих.
Следует еще раз подчеркнуть, что, несмотря на лишь ограниченные специфичность и память, характерные для распознавания алло- и ксеногенных трансплантатов у беспозвоночных, их иммунная система функционирует аффективно и успешно. Так или иначе, внедрение патогенных микроорганизмов и паразитов вызывает у беспозвоночных быстрый иммунный ответ, что и обеспечивает громадное разнообразие и изобилие этих животных.
У беспозвоночных имеются молекулы - предшественники МНС и молекулы с иммуноглобулин-подобными доменами. Аллогенное распознавание, обнаруженное у многих беспозвоночных, указывает на возможное присутствие у них молекул — предшественников главного комплекса гистосовместимости (МНС). Поскольку иммуноглобулинами эти животные нe обладают, можно предполагать, что система МНС сформировалась раньше и независимо от системы иммуноглобулинов у позвоночных. Иными словами, у примитивных позвоночных при сохранении системы МНС отдельно и независимо развивалась система иммуноглобулинов, обеспечивающая более тонкое распознавание посредством циркулирующих антител и специфичных рецепторов клеточной поверхности. В ходе дальнейшей эволюции позвоночных происходила, по-видимому, возрастающая интеграция МНС и системы иммуноглобулинов, что обеспечило высокий уровень регуляции, необходимый для взаимодействия антигенпрезентирующих клеток (АПК) и лимфоцитов. Это представление, однако, остается гипотетическим; нет ни структурных, ни функциональных доказательств экспрессии клетками беспозвоночных гликопротсинов МНС или димерных рецепторов для аллоантигенов. Кроме того, у беспозвоночных может отсутствовать реакция смешанной культуры лейкоцитов (см. рис. 15.1), которая у позвоночных служит функциональным маркером МНС. Поэтому возникло другое предположение: некоторые исследователи считают, что молекулы МНС позвоночных произошли от белков теплового шока (см. ниже).
В то же время обнаружены β2-микроглобулин-подобные молекулы у земляных червей, ракообразных и насекомых, что подтверждает возможность существования антигенов - предшественников МНС у беспозвоночных. β2-Микроглобулин позвоночных колируется геном, не сцепленным с МНС, но ассоциирован с молекулами МНС класса I и принадлежит к суперcемейству иммуноглобулинов. Таким образом, молекулы МНС могут быть потомками одной и той же, со держащей один домен и сходной с β2-иммуноглобулином молекулы, многочисленные разновидности-производные которой возникли в результате перестроек и дупликаций генов и давления отбора.
Наконец, существует целая группа обнаруженных у беспозвоночных молекул, таких как Thy-I (присутствует в мозге кальмара), амальгам, фасциклин II, нейроглиан и гемолин (все у насекомых), также принадлежащих к суперсeмейству иммуноглобулинов; предполагается, что эти молекулы появились в процессе эволюции для опосредования межклеточных взаимодействий и потенциально могут обеспечивать распознавание иммунной системой «не-своего». Этот уровень эволюции, по-видимому, достигнут у насекомых, обладающих гемолином (см. выше).