ИММУНОЛОГИЯ - Ройт А. - Мир 2000
Глава 18. Иммунитет к паразитарным инвазиям
■ Паразитами заражена значительная часть населения земного шара. Как правило, они специфичны в отношении хозяина и в большинстве случаев вызывают хроническую инвазию. Многие паразиты распространяются беспозвоночными-переносчиками и имеют сложный жизненный цикл, причем на разных стадиях развития синтезируют различные антигены.
■ Устойчивость организма-хозяина обеспечивают различные защитные реакции. Способностью уничтожать как простейших, так и гельминтов обладают эффекторные клетки - макрофаги, нейтрофилы, эозинофилы и тромбоциты. Они секретируют цитотоксические соединения, например высокоактивные метаболиты кислорода и оксид азота, и функционируют более эффективно при активации цитокинами.
■ Основная роль в развитии иммунитета принадлежит Т-клеткам. Антитела, как сами по себе, так и в сочетании с комплементом, эффективно действуют против внеклеточных паразитов. Они усиливают фагоцитарный и цитотоксический потенциал эффекторных клеток и могут предотвращать внедрение паразитов в новые клетки организма-хозяина.
■ Паразиты различными путями избегают действия иммунного ответа хозяина, иногда используя ответные реакции хозяина для своего собственного развития и в большинстве случаев противодействуя им.
■ При заражении паразитическими червями повышаются численность эозинофилов и уровень циркулирующих антител IgE. Для развития иммунного ответа важны Тх2- и Тх1-клетки. Тх2-клетки необходимы для изгнания кишечных гельминтов. Продукты тучных клеток взаимодействуют с эозинофилами.
■ Т-клетки, необходимые для осуществления защитных реакций, могут относиться к популяции как CD4+, так и CD8+. Тх1-клетки обеспечивают защиту против внутриклеточных простейших путем секреции ИФγ, активирующего макрофаги.
■ Паразитарные инвазии сопровождаются образованием больших количеств неспецифических антител, спленомегалией и гепатомегалией. Иммунопатологические эффекты в основном могут быть опосредованы Т-клетками.
Как правило, паразитарные инвазии вызывают активацию ряда иммунологических защитных механизмов, гуморальных и клеточных, причем эффективность ответных реакций зависит от природы паразита и стадии инвазии. В данной главе рассмотрены основные аспекты иммунитета к паразитарным инвазиям и особенно подробно к заболеваниям, вызываемым некоторыми из наиболее важных паразитов человека (рис. 18.1).
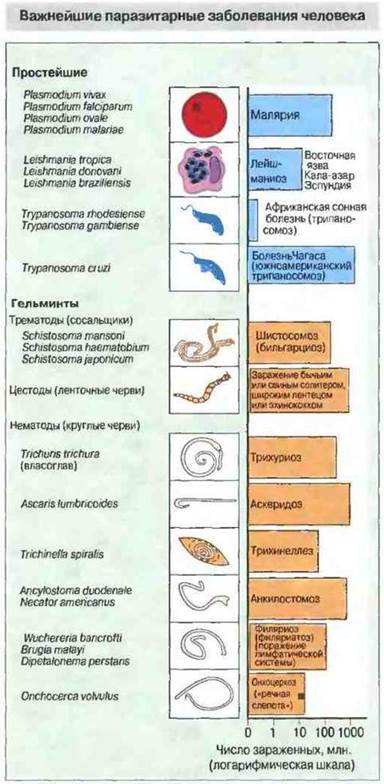
Рис. 18.1. Статистика зараженности паразитами по данным Всемирной организации здравоохранения (1993 г.).
Паразитические простейшие способны обитать в кишечнике (например, амебы), в крови (например, африканские трипаносомы), внутри эритроцитов (например, Plasmodium spp.), в макрофагах (например, Leishmania spp. и Toxoplasma gondii), в том числе локализованных в печени и селезенке (например, Leishmania spp.), или в мышцах (например, Trypanosoma cruzi). К паразитическим червям — гельминтам, которые заражают человека, относятся трематоды, или сосальщики (например, шистосомы), цестоды, или ленточные черви (например, широкий лентец, цепни, эхинококк), и нематоды, или круглые черви (например, Trichinella spiralis, анкилостомы, острицы, аскариды и нитчатки, или филярии). Цестоды и половозрелые анкилостомы заселяют кишечник, половозрелые шистосомы обитают в кровеносных сосудах, а некоторые филярии в лимфатической системе (рис. 18.2). Очевидно, что при таком разнообразном распространении возможны и самые разнообразные патологические реакции.
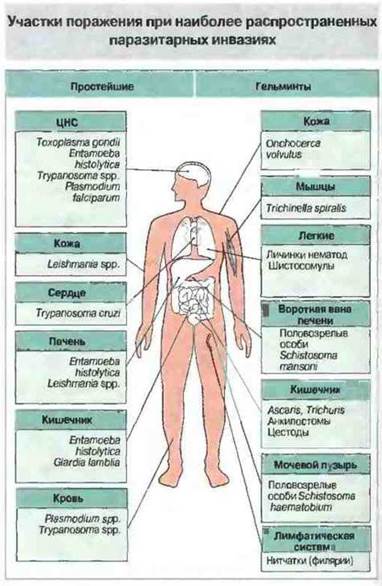
Рис. 18.2. Участки поражения при наиболее распространенных паразитарных инвазиях.
Многие гельминты имеют сложный жизненный цикл с миграцией через различные участки организма-хозяина, причем в различных тканях они проходят определенные стадии своего развития. На последнем этапе развития гельминт достигает области, где окончательно созревает и проводит оставшуюся часть жизненного цикла (рис. 18.3). Анкилостомы и личинки шистосом заражают хозяина, непосредственно проникая через кожу; ленточные черви и круглые черви попадают в организм человека через рот; распространение филярий зависит от промежуточного хозяина-насекомого или от переносчика. Большинство паразитических простейших распространяется насекомыми-переносчиками, а Toxoplasma, Giardia и амебы попадают в организм человека через рот. Возбудителей малярии переносят комары, трипаносом — мухи цеце, клопы и др. (в частности Т. cruzi - клопы-хищнецы подсемейства Triatominae), Leishmania — москиты. Разнообразие путей заражения создает дополнительные проблемы при разработке способов иммунизации.
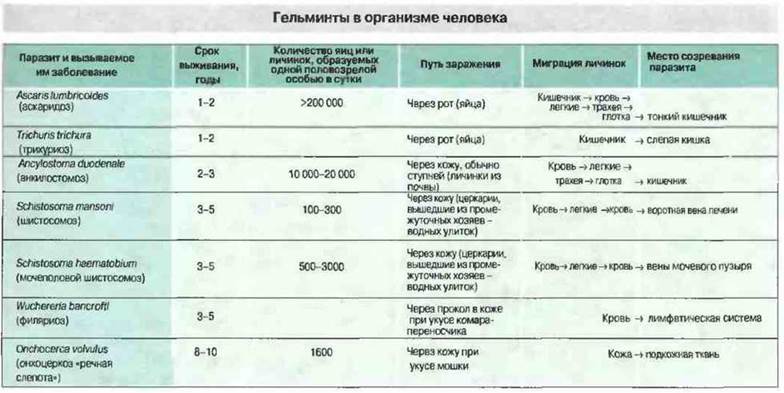
Рис. 18.3. Сроки выживания, размножение и пути миграции гельминтов в организме человека.
ОСНОВНЫЕ ОСОБЕННОСТИ ПАРАЗИТАРНЫХ ИНВАЗИЙ
Паразитами заражена большая часть населения земного шара
Паразитарные болезни представляют собой серьезную медицинскую проблему, особенно в тропических странах (рис. 18.1). Малярия, например, служит причиной смерти 1—2 млн. человек ежегодно. Кишечными гельминтами заражена треть населения земного шара; тяжесть заболевания зависит от степени инвазии, однако у детей даже инвазии средней тяжести могут вызывать задержку роста и замедление умственного развития. Паразитарные болезни характеризуются также анемией и истощением.
Паразитарные инвазии имеют ряд общих особенностей. Паразитические простейшие и черви значительно крупнее бактерий и вирусов (рис. 18.4) и соответственно обладают большим количеством и разнообразием антигенов. Для некоторых видов характерна смена поверхностных антигенов — антигенная изменчивость. Паразиты, имеющие сложный жизненный цикл, могут экспрессировать некоторые антигены только на определенной стадии развития, вызывая соответствующий специфический иммунный ответ. Так, у малярийных паразитов белковая оболочка спорозоита (инфекционная, или расселительная, стадия, в виде которой плазмодий передается комарами и инфицирует организм-хозяин) вызывает образование специфичных антител, не реагирующих с паразитом на эритроцитарной стадии его цикла развития. Паразитический червь Т. spiralis на разных стадиях жизненного цикла также экспрессирует различные антигены.
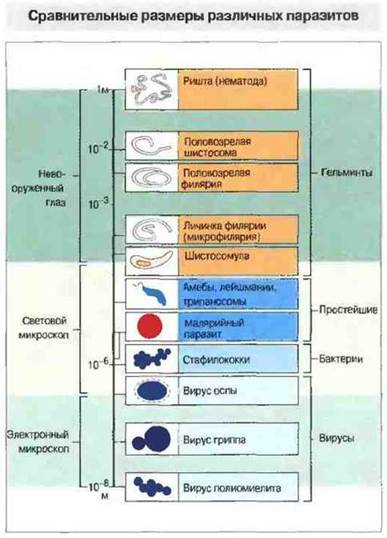
Рис. 18.4. Сравнительные размеры различных паразитических организмов.
Простейшие, достаточно мелкие по размерам, чтобы паразитировать внутри клеток человека, обладают особыми свойствами проникновения в клетки и ткани организма-хозяина. Мерозоиты малярийного плазмодия (стадия цикла развития, на которой плазмодий способен проникать в эритроциты) связываются на поверхности клеток крови с определенными рецепторами и используют для проникновения внутрь клетки специальную органеллу — роптрий. Паразитические лейшмании для внедрения в макрофаги используют рецептор комплемента, чтобы помочь клеткам хозяина захватить их. Эти паразиты могут также проникать внутрь клетки, взаимодействуя с маннозо-фукозным рецептором на поверхности макрофагов.
Большинство паразитов специфично в отношении хозяина
За миллионы лет эволюции паразитические организмы хорошо адаптировались к своим хозяевам и обнаруживают ярко выраженную специфичность к ним. Например, возбудители малярии птиц, грызунов или человека способны размножаться только в организме данного хозяина. Из этого правила имеются некоторые исключения: например, паразитическое простейшее Т. gondiiспособно не только заражать все содержащие ядро клетки млекопитающих и размножатся в них, но также инфицировать незрелые эритроциты млекопитающих, культивируемые клетки насекомых и ядерные эритроциты птиц и рыб. Сходным образом, свиной солитер способен заражать человека.
Устойчивость хозяина к паразитарной инвазии может быть генетически обусловленной
Резистентность индивидуальных хозяев к инвазии варьирует и может контролироваться различными генами иммунного ответа. Ряд линий мышей (и некоторые люди), имеющие определенные гены МНС, не обладают способностью к активной продукции антител против одного из пептидов оболочки спорозоитов малярийного паразита, поскольку их Т-клетки не сенсибилизируются. В то же время наличие некоторых антигенов HLA, широко распространенных у жителей Западной Африки, но редко встречаемых у европеоидов, по-видимому, коррелирует с иммунитетом к тяжелым формам малярии.
Гены не-МНС также могут иметь важное значение.
✵ Чувствительность мышей к заражению Leishmania donovani и некоторыми другими паразитами определяется одним доминантным геном, регулирующим активацию макрофагов (см. гл. 1).
✵ Мерозоиты возбудителя малярии Plasmodium vivax используют для проникновения в эритроциты специфический рецептор их поверхности - антиген группы крови Даффи. У некоторых африканских народностей этот антиген утрачен — возможно, под давлением отбора — и они высокоустойчивы к заражению этим паразитом.
Для большинства гельминтозов характерно присутствие значительного количества паразитов у относительно небольшого числа лиц, однако следует отметить, что это необязательно обусловлено генетическими различиями в устойчивости. Результаты поведенческих исследований указывают на то, что даже в небольших сообществах отдельные индивиды сильно различаются по риску заражения, так как у них весьма различны возможности контакта с паразитами.
Многие паразитарные инвазии — хронические заболевания
Паразиту не выгодна гибель организма-хозяина, по крайней мере до тех пор, пока не произойдет его передача другому хозяину. При хронической инвазии тип иммунного ответа с течением времени меняется и зависит от наличия циркулирующих антител, персистентной антигенной стимуляции и образования иммунных комплексов (рис. 18.5). Для хронических инвазий типичны иммуносупрессия и иммунопатологические реакции.
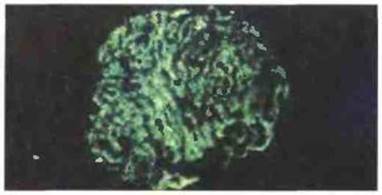
Рис. 18.5. Отложение иммунных комплексов при нефротическом синдроме, характерном для четырехдневной малярии. Микрофотография (флуоресцентная электронная микроскопия низкого напряжения) почечного клубочка (материал биопсии от нигерийского ребенка). При малярии, вызванной Plasmodium malariae, в результате отложения иммунных комплексов может развиться гломерулонефрит. Срез окрашен ФИТЦ, конъюгированным с IgG к антигенам человека; можно видеть гранулярное отложение иммуноглобулинов в капиллярных петлях почечного клубочка. (Фото любезно предоставлено д-ром V. Houba.)
Устойчивость организма-хозяина зависит от разнообразных эффекторных механизмов иммунитета
Развитие иммунитета - это сложный процесс, основанный на взаимодействии многих типов клеток в течение определенного периода времени. Часто эффекты осуществляются локально, и различные клетки, секретирующие отдельные медиаторы, могут находиться в участках иммунологического отторжения. Кроме того, иммунные реакции, препятствующие размножению паразита в организме, могут отличаться от реакций, обеспечивающих развитие окончательной устойчивости. При некоторых гельминтозах формируется «сопутствующий иммунитет»; первичная инвазия не ликвидируется, но стабилизируется, и хозяин приобретает резистентность к заражению новыми гельминтами того же вида.
Гуморальный ответ как таковой необходим для уничтожения внеклеточных паразитов, например локализующихся в крови (рис. 18.6), жидкостях организма или кишечнике. Однако то, какой тип ответа в итоге будет главным, обеспечивающим защиту, зависит от природы паразита. Например, антитела сами по себе или в сочетании с комплементом могут повреждать некоторых внеклеточных паразитов, однако их действие усиливается, когда они функционируют совместно с эффекторными клетками. Как отмечено выше, при одной и той же инвазии в борьбе с паразитом на разных стадиях его жизненного никла участвуют различные эффекторные механизмы. Так, при малярии антитела к внеклеточным формам паразита блокируют их способность проникать в новые клетки, в то время как клеточноопосредованный ответ предотвращает развитие шизонтов в гепатоцитах. Протективный иммунитет к малярии не коррелирует непосредственным образом с уровнем антител и может быть индуцирован даже в их отсутствие. Это установлено на мышах, иммунизированных специально полученным штаммом Salmonellatyphimurium; в геном бактерий этого штамма методом генетической инженерии внедрен ген, колирующий поверхностный антиген спорозоитов малярийного плазмодия. При последующем заражении таких мышей спорозоитами у животных, несмотря на отсутствие специфических антител, развивался иммунитет к возбудителю малярии.
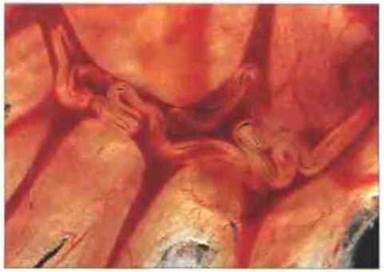
Рис. 18.6. Пары половозрелых шистосом в кровеносных сосудах брыжейки. Шистосомы высокоустойчивы к интенсивному воздействию эффекторных механизмов иммунитета. Половозрелые особи могут персистировать в организме в среднем 3-5 лет. (Фото любезно предоставлено д-ром A. Agnew.)