Зоологія хордових - підручник - Й. В. Царик - 2013
Розділ 5. НАДКЛАС ЧОТИРИНОГІ (НАЗЕМНІ ХРЕБЕТНІ) TETRAPODA, seu QUADRIPEDA.
АНАМНІЇ (ANAMNIA) й АМНІОТИ (AMNIOTA)
5.4. КЛАС ССАВЦІ, або ЗВІРІ, MAMMALIA, seu THERIA
5.4.1. Особливості організації ссавців
Зовнішній вигляд. Дуже різноманітні за розміром і зовнішнім виглядом. Найменший зі сучасних ссавців - багатозуб карликовий Suncus etruscus (з мідицеподібних) масою 1,2—1,7 г і довжиною тіла 3,8-4,5 см. Найбільший із наземних ссавців - африканський слон висотою 3,5 м і масою до 4-5 т, а серед водних - синій кит, окремі особини якого сягають довжини 33 м і маси 150 т (як 30-35 слонів). Тіло ссавців розділене на голову, шию, тулуб, парні кінцівки і хвіст.
У тюленевих, сирен і, особливо, китоподібних є обтічне тіло, кінцівки перетворюються на ласти; рибоподібного вигляду сиренам і китам надає шкірястий хвостовий плавець. Навколоводні ссавці (качконіс, хохуля, бобер, нутрія, видра) мають щільний волосяний покрив, що протидіє намоканню, вкорочену шию, розвинуті плавальні перетинки між пальцями, більше або менше сплющений хвіст. У багатьох коне- і оленеподібних аналогічний вигляд: високі ноги, компактний тулуб, довга рухлива шия, що допомагає переміщатися на далекі відстані й уникати однакових ворогів - великих псоподібних. Схожі між собою за формою тіла зайцеподібні та мишоподібні. Пристосування до швидкого пересування на задніх кінцівках у відкритих біотопах привело до вироблення подібної форми тіла - слабкі передні й міцні задні кінцівки, довгий хвіст-противага: у кенгуру (сумчасті), африканських стрибунців
(мідицеподібні) та різних мишоподібних - тушканчиків, піщанок тощо. У рийних видів тіло набуває валькуватої форми, кінцівки і хвіст вкорочені (ховрахи, бабаки, полівки та ін.). У разі переходу до підземного способу життя поряд із валькуватим тулубом значного розвитку набувають передні кінцівки, що беруть участь у ритті ґрунту (сумчасті і звичайні кроти й ін.).
У деревних видів пристосуванням до пересування є короткі, але сильні, з гострими кігтями, кінцівки, довгий пухнастий хвіст, який допомагає під час стрибків (вивірки, куниці та ін.). Довгі кінцівки приматів забезпечують їм пересування у кронах дерев. Опосуми, деякі види мавп, деревні мурахоїди, ящури і їжатці використовують для хапання хвіст. У сумчастих летяг, звичайних летяг, шерстокрила шкірна складка з боків тіла дає змогу робити довгі планерувальні стрибки. Характерно, що серед сумчастих трапляються життєві форми, подібні до багатьох плацентарних ссавців.
Отже, представники різних рядів, що живуть у подібних умовах і виробили близький спосіб життя, можуть мати й подібну форму тіла (конвергентна подібність). Такі групи видів називають «життєвими формами», або екологічними типами.
Покриви. Порівняно товста шкіра, як у всіх хребетних, складається з двох шарів, багата на залози і має рогові утворення - волосся, кігті та ін. (рис. 5.46). Епідерміс багатошаровий. У його основі розташований мальпігієвий шар, утворений циліндричними або призматичними, живими, здатними до розмноження клітинами. Ближче до поверхні ці клітини втрачають ядра і здатність до поділу. Вони набувають плоскої форми, роговіють, утворюють мертвий зовнішній шар. Ці клітини злущуються з поверхні, поступово їх заміщують нові з клітин мальпігієвого шару. Забарвлення шкіри зумовлене пігментами, що у вигляді зерен меланіну містяться у клітинах росткового шару, у міжклітинних проміжках і у спеціальних пігментних клітинах (меланобласти, меланофори). Епідермальний шар досягає найбільшої товщини в місцях постійного тертя під час пересування тварини; тут можуть утворюватися мозолі (підошви лап, сідничні мозолі деяких мавп, мозолі на колінах верблюдів тощо).
Дерма (коріум) є глибоким шаром шкіри ссавців. Вона багата на колагенові й еластинові волокна, що переплітаються між собою й утворюють сітчасту структуру. У глибоких шарах дерми відкладається жирова тканина - підшкірна жирова клітковина, яка у деяких ссавців (кити, дельфіни) розвинена дуже добре, і це компенсує повну редукцію волосяного покриву.
Такі властивості дерми, як еластичність, міцність (завдяки еластиновим і колагеновим волокнам), погана теплопровідність (завдяки жировій клітковині), у поєднанні з механічною міцністю рогового шару епідермісу і його похідних роблять шкіру ссавців надійним засобом захисту тіла від пошкоджень і впливу несприятливих зовнішніх чинників. Крім того, у дермі розміщені базальні частини залоз, фолікули волосся, нервові структури, що забезпечують чутливість шкіри, добре розвинені судини, капілярна сітка, яка, проникаючи у верхній сосочковий шар дерми, забезпечує живлення епідермісу і терморегуляційну функцію шкіри. У дермі можуть розвиватися гладенькі м’язові волокна.
Окрім волосся, специфічними для ссавців є молочні залози - видозмінені потові залози. Саме завдяки наявності молочних залоз цей клас хребетних отримав назву ссавці. У ссавців наявні потові, сальні й пахучі залози. Пахучі залози є похідними або потових, або сальних. Ці залози зазвичай складні; їхня базальна частина розміщена в коріумі, вивідні протоки відкриваються на поверхні шкіри або у волосяній сумці.
За будовою вивідних проток виділяють трубчасті й альвеолярні залози. Перший тип - трубчасті залози. До них належать потові залози, які переважно відкриваються на поверхню шкіри, інколи - у волосяну сумку. Внутрішній кінець потової залози має клубочок (див. рис. 5.46). У стінках таких залоз міститься одношаровий епітелій, епітеліальні клітини якого виділяють власне піт. Піт на 97-99% складається з води, у якій розчинені сечовина і креатин, леткі жирні кислоти й солі (вони є і в сечі). Отже, з потом виділяються продукти розпаду, проте основна функція потових залоз - терморегуляційна (піт випаровується й охолоджує тіло). Потовиділення регулюють теплові центри головного та спинного мозку. Добре розвинені потові залози у приматів і копитних, порівняно слабко - у псових, котових, зайцеподібних і мишоподібних. У видів зі слабким розвитком потових залоз терморегуляція відбувається по-іншому. Наприклад, у собак під час перегріву тепловіддача посилюється завдяки пришвидшенню поверхневого дихання (поліпное) і випаровуванню води зі слини висунутого язика та слизової оболонки ротової порожнини. У сиренових, китоподібних, лінивців, ящурів, кротів потових залоз немає.
Другий тип залоз - це альвеолярнізалози, до яких належать сальні залози. Вони мають гроноподібну будову, а їхні протоки відкриваються у волосяні сумки.
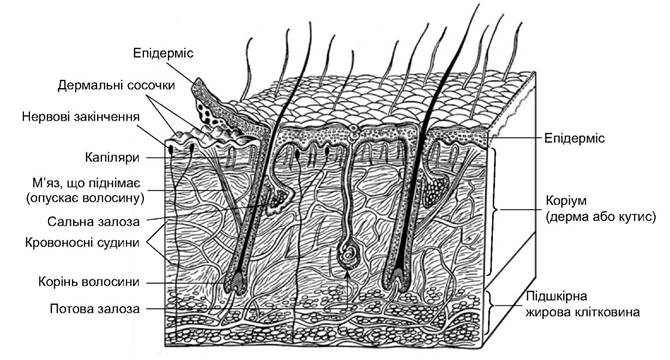
Рис. 5.46. Будова шкіри ссавців
Стінки залоз утворені багатошаровим епітелієм. У його клітинах відбувається жирове переродження з утворенням жирового секрету, що змащує поверхню шкіри та волосся, сприяючи збереженню еластичності й перешкоджаючи проникненню мікробів і грибків. Таких залоз немає у китоподібних, лінивців, деяких мідицеподібних.
Мускусні залози - шкірні залози, виділяють пахучу речовину мускус, яка слугує, головно, для приваблювання самок і мічення території, видового розпізнавання. Вони є видозміною потових або сальних залоз, іноді бувають змішаної будови, поєднуючи альвеолярну і трубчасту будову. Пахучими залозами є анальні залози багатьох псоподібних, особливо мустелових, мускусні залози кабарги, бобрів, хохулі й ондатри; передочні залози багатьох оленеподібних (олені, антилопи, вівці), копитні залози козлів тощо. Сильний смердючий секрет анальних залоз американських скунсів, деяких тхорів слугує для самозахисту. Сукупність запахів секретів мускусних, сальних і потових залоз допомагає тваринам відрізняти особини свого виду від інших, полегшує зустріч самців і самок.
Молочні залози - видозмінені потові залози - розвинені у самок усіх ссавців. В однопрохідних молочні залози зберігають трубчасту будову й розташовані групами - залозистими полями: у качконосів на череві, у єхидни у виводковій сумці. У них сосків немає і протоки залоз відкриваються у волосяні сумки; малята злизують крапельки молока з волосся. У решти ссавців молочні залози мають складнішу, гроноподібну будову; протоки молочних залоз відкриваються на сосках. У частини видів соски двома рядами розташовані від передніх кінцівок до пахвини (міди- цеподібні, хижі, мишоподібні), в інших є лише одна пара сосків (примати, сирени, слони, кажани) або тільки соски в пахвині. У більшості копитних молочні залози правого і лівого боків зливаються у розташоване в пахвині вим’я, що має два або чотири соски. Кількість сосків у різних видів ссавців варіює від 2 до 12 пар і приблизно відповідає кількості малят, які мають народитися.
Волосяний покрив - характерна особливість ссавців. У більшості звірів він розвинений на всіх ділянках тіла (його немає на губах, у деяких видів - на підошвах) і виконує термоізоляційну функцію, слугує рецептором дотику, захищає шкіру від пошкоджень і деяких паразитів, поліпшує аеро- та гідродинамічні властивості тіла, забезпечує видоспецифічність забарвлення. Лише у деяких ссавців (китоподібні, слони, носороги, бегемоти) волосяного покриву на тілі практично немає; на деякий час вони вкриваються зачатками волосся під час утробного розвитку в організмі самки.
Волосся ссавців розвивається з епідермального зачатка, який поступово занурюється в коріум (рис. 5.47). Із зовнішніх шарів епідермального зачатка формуються волосяна сумка і сальні залози; з внутрішніх шарів - власне волосина. Волосина росте завдяки розмноженню базальних клітин, що є в цибулині - основі волосини. Поступово витіснені догори клітини волосини роговіють; уся волосина, окрім базального відділу, є мертвим утвором. Сформована волосина складається зі стовбура, що виступає над поверхнею шкіри, і кореня, що розміщений у шкірі. У стовбурі волосини серцевина має порожнисту будову і складається з плоских зроговілих клітин із прошарками повітря. Особливо велика кількість повітря характерна для волосин жителів територій із холодними зимами; це збільшує теплоізоляційні властивості їхнього волосяного покриву. Пухку серцевину оточує щільний кірковий шар, утворений зроговілими, витягнутими по поздовжній осі волосини клітинами. Цей шар забезпечує міцність і пружність волосини, у ньому містяться пігменти. Ззовні кірковий шар має шкірку з плоских і прозорих рогових клітин, покладених одна на одну, мов черепиця. Корінь волосини знизу розширений у цибулину, що складається з живих клітин. Сосочок коріума з кровоносними судинами впинається в цибулину знизу, постачаючи поживні речовини до її клітин. Корінь волосини міститься у волосяній сумці, яка є впинанням епідермального шару. Найближчий до волосини шар сумки називають волосяною піхвою, а зовнішній - волосяним мішком. До його нижньої частини прикріплені гладкі м’язи, скорочення яких змінює кут нахилу волосини. У волосяній сумці розташовані й закінчення чутливих нервів.
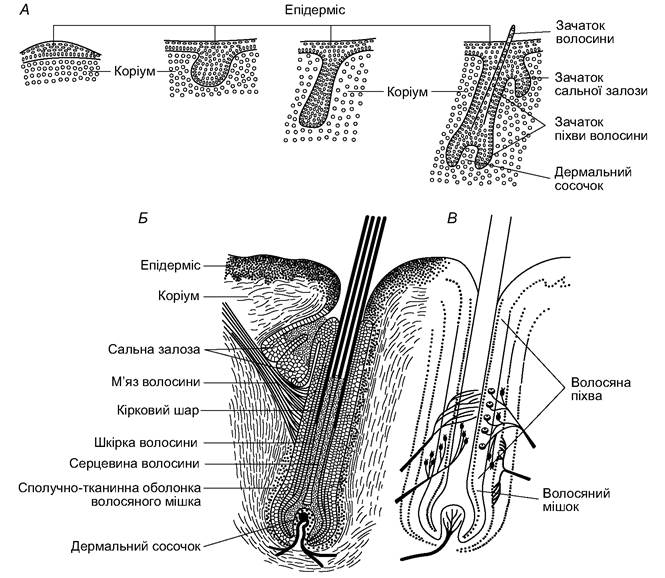
Рис. 5.47. Розвиток (А), будова (Б) та іннервація (В) волосини
У ссавців волосся неоднорідне. Найбільші, зазвичай, поодинокі волосини-щетинки, що різко виступають над хутровим покривом, називають вібрисами. Вони є органами дотику; розташовані на частинах тіла, якими тварина найчастіше торкається навколишніх предметів (кінець морди, черево, кінцівки) і які мають у волосяній сумці багато нервових закінчень. Верхній ярус хутрового покриву утворений остьовим волоссям, нижній ярус - ниткоподібними, часто спірально закрученими тонкими пуховими волосинами, що відіграють особливо важливу роль у теплоізоляції тіла. У небагатьох ссавців хутро складається з однієї категорії волосся: літнє хутро оленів і кабанів - зі самої лише ості, хутро землериїв (кріт, сліпак) - з пуху. Щетина (свині) та голки (єхидни, їжаки, їжатці) - видозмінені остьові волосини.
Звичайно волосся розташоване на тілі впорядковано. Зазвичай воно нахилене в певному напрямі, що забезпечує обтічність тіла потоками повітря і води. Головний напрям ворсу - від голови до хвоста. Характер ворсу змінюється в тих місцях, де на шкірі часто утворюються складки і розтяжки. Ворс тісно пов’язаний зі способом життя і характером руху. Наприклад, лінивці постійно висять на деревах униз спиною, і їхнє волосся укладене в напрямі від черева до спини, що можна пов’язати як із силою тяжіння, так і з впливом дощів. У підземних жителів (кроти, сліпаки), що нерідко пересуваються як переднім, так і заднім ходом, коротке волосся стоїть сторчма.
Подібний волосяний покрив мають жителі щільних трав’янистих заростей і підстилки (землерийки).
Хутро зношується і потребує заміни, яка відбувається під час линьки. У тропіках звірі линяють постійно по ходу зношування волосся (у мавп може тривати протягом усього року). Подібне явище в помірних широтах спостерігають у жителів нір, у яких швидко витираються деякі частини хутрового покриву; на цих ділянках відбувається часткова линька. У більшості видів помірних широт звичайно покриви змінюються двічі на рік - восени і навесні; линька супроводжується зміною структури хутра, а нерідко і його забарвлення. У разі зміни літнього вбрання на зимове збільшується щільність волосяного покриву, тому теплоізоляційна якість зимового волосяного покриву порівняно з літнім різко підвищується.
Кінцеві фаланги пальців більшості ссавців захищені роговими кігтями - похідними епідермісу (рис. 5.48). У деревних форм вони гострі й сильно загнуті, у рийних - видовжені та плоскіші. В усіх котових (крім гепарда) кігті втяжні; кіготь разом із кінцевою фалангою особливими сухожиллями притягується до дорзальної поверхні передостанньої фаланги і тому не затуплюється під час пересування. У багатьох приматів кігті перетворилися на нігті, прикриваючи кінці пальців тільки зверху; знизу розвинута м’яка подушечка, що підвищує дотикові властивості пальців. Ускладнення кігтів привело до утворення копит - товстих рогових утворів, що охоплюють майже цілком кінцеву фалангу. Особливо добре копита розвинені у видів, які швидко бігають (коні, антилопи, козли та ін.).
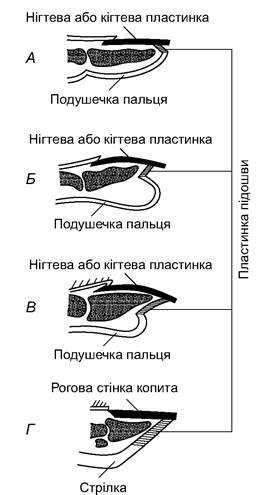
Рис. 5.48. Поздовжні розрізи через кіготь мавпи (А) і ніготь людини (Б), кіготь собаки (В) і копито коня (Г)
Унаслідок значного розростання зроговілого епітелію утворюються роги у носорогів і роги порожнисторогих - порожнисті рогові чохли, що розміщені на кісткових стрижнях і зростаються з лобовими кістками. Роги оленів - кісткові утвори, похідні дерми; їх тварини щорічно скидають. У багатьох ссавців на хвості й на кінцівках розвиваються рогові луски, подібні до лусок плазунів (сумчасті, мідицеподібні, мишоподібні). У ящурів великі, ромбічні рогові луски, черепицеподібно налягаючи одна на одну, вкривають усе тіло. У броненосців (неповнозубі) панцир утворений кістковими щитками (похідні коріума), які вкриті зверху роговими пластинками - похідними епідермісу.
Отже, шкірний покрив ссавців виконує багато функцій. Секрети шкірних залоз підтримують її еластичність, захищають від намокання і проникнення інфекцій; запах секретів відіграє важливу роль у внутрішньовидових відносинах. Роговий шар епідермісу захищає шкіру від механічних ушкоджень, зменшує втрати води. Волосяний покрив і жирова підшкірна клітковина зменшують тепловіддачу, сприяють підтриманню сталої температури тіла. Окрім того, жирові запаси у підшкірній клітковині - енергетичний резерв. Діяльність потових залоз визначає участь шкіри у водно-сольовому обміні й терморегуляції. Пігменти волосся і шкіри забезпечують видоспецифічне забарвлення тварин.
У водних ссавців шкіра і волосяний покрив підвищують гідродинамічні властивості їхнього тіла. У китоподібних дуже товста шкіра без волосяного покриву з гладким і пружним епідермальним шаром та значним коріумом, сосочки якого особливо глибоко проникають в епідерміс. Проміжки між складно переплетеними еластиновими і колагеновими волокнами коріума заповнені жиром. Така конструкція шкіри забезпечує її високу пружність: прогинаючись під тиском, шкіра гасить турбулентні завихрення, що порушують плавне (ламінарне) обтікання водою тіла тварини. Цьому ж сприяють рефлекторні хвилі скорочень підшкірної м’язової тканини, що пробігають по тілу дельфіна з прискоренням руху. Щільне хутро водних ссавців (хохуля, бобри, видри, норки та ін.) має добре розвинену підпуш із покрученого волосся. Остьове та напрямне волосся має стрілоподібну форму і виступає над підпушшю; у воді їхня верхня частина відхиляється в протилежний до руху бік і лягає на пружний шар пухового волосся. Тому волосяний покрив цих тварин утворює пружну систему, аналогічну до пружної шкіри китоподібних.
Скелет ссавців вирізняється різноманітністю будови, що відповідає значній різноманітності способів їхнього руху.
Хребет складається з шийного, грудного, поперекового, крижового і хвостового відділів (рис. 5.49). Його характерна особливість - платицельна (з плоскими поверхнями) форма хребців, між якими розташовані хрящові міжхребцеві диски. Добре виражені верхні дуги. У шийному відділі - сім хребців, від довжини яких залежить і довжина шиї; тільки у ламантина і лінивця Choloepus hoffmani їх шість, а у лінивців роду Bradypus - 8-10. Шийні хребці дуже довгі у жирафа й дуже короткі у китоподібних, які не мають шийного перехвату. До хребців грудного відділу прикріплені ребра, які утворюють грудну клітку. Грудина, що їх замикає, плоска; лише у кажанів і рийних ссавців зі значно розвиненими передніми кінцівками (наприклад, кротів) є невеликий гребінь (кіль) для прикріплення грудних м’язів. У грудному відділі - 9-24 (частіше 12-15) хребців, останні 2-5 грудних хребців мають несправжні ребра, які не доходять до грудини. У поперековому відділі від двох до дев’яти хребців з добре розвиненими поперечними відростками. Крижовий відділ утворений 4-10 зрослими хребцями, з яких тільки два перші - справжні крижові, а решта - хвостові. Кількість вільних хвостових хребців коливається від 3 (у гібона) до 49 у довгохвостого ящура. Ступінь рухомості окремих хребців різний. У дрібних тварин, які бігають і повзають, вона велика по всій довжині хребта, тому їхнє тіло може вигинатись у різних напрямах і навіть згортатись у клубок. Менш рухомі хребці грудного і поперекового відділів є у великих за розмірами тварин, що швидко рухаються. У ссавців, які пересуваються на задніх кінцівках (кенгуру, тушканчик, стрибунець), найбільші за розмірами хребці є в основі хвоста і крижового відділу хребта, а далі вперед їхній розмір поступово зменшується. У копитних, навпаки, хребці, особливо їхні остисті відростки, крупніші в передній частині грудного відділу, де до них прикріплена потужні м’язи шиї та частково передніх кінцівок.
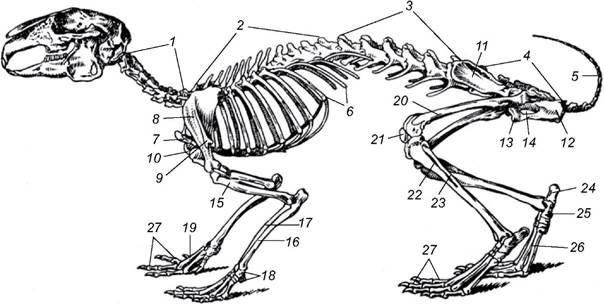
Рис. 5.49. Скелет кроля:
1 - шийні хребці; 2 - грудні хребці; 3 - поперекові хребці; 4 - крижі; 5 - хвостові хребці; 6 - ребра; 7 - руків'я грудини; 8 - лопатка;
9 - акроміальний відросток лопатки; 10 - коракоїдний відросток лопатки; 11-13 - клубовий (11), сідничний (12) і лобковий (13) відділи безіменної кістки; 14 - замикальний отвір; 15 - плече; 16 - ліктьова кістка; 17 - променева кістка; 18 - зап'ясток; 19 - п'ясток;
20 - стегно; 21 - колінна чашечка; 22 - велика гомілкова кістка; 23 - мала гомілкова кістка; 24 - п'яткова кістка; 25 - таранна кістка; 26 - плесно; 27 — фаланги
Череп ссавців синапсидного типу. В ньому наявна вилична дуга, утворена кістками: верхньощелепна-вилична-луската. Від рептилій череп ссавців відрізняється помітно більшим об’ємом мозкової коробки, зменшенням кількості кісток (завдяки їхній редукції і зростанню) і приєднанням до хребта двома виростками (рис. 5.50). Нижня щелепа утворена лише однією парною кісткою - зубною, яка безпосередньо прикріплена до виличного відростка лускатої кістки. Зчленівна кістка нижньої щелепи рептилій зменшена у розмірах і перетворена в одну з кісточок середнього вуха ссавців - молоточок (malleus). Друга частина апарату середнього вуха ссавців утворена квадратною кісткою, що перетворилась у ковадло (incus); третя слухова кісточка - стремінце (stapes) - утворилась із верхнього відділу під’язикової дуги - гіомандибуляре - уже в земноводних і зберігається у всіх наземних хребетних.
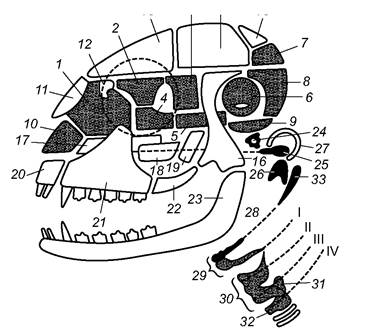
Рис. 5.50. Схема черепа ссавців (темні - кістки, які заміщуються, світлі - покривні кістки):
1 - решітчаста; 2 - очноямково-клиноподібна; 3 - крило-клиноподібна; 4 - передня клиноподібна; 5 - основна клиноподібна;
6 - кам'яниста; 7 - верхня потилична; 8 - бічна потилична; 9 - основна потилична; 10 - залишок хрящового черепа (носова перегородка); 11 - носова; 12 - слізна; 13 - лобова; 14 - тім'яна; 15 - міжтім'яна; 16 - луската; 17 - леміш; 18 - піднебінна;
19 - крилоподібна; 20 - міжщелепна; 21 - верхньощелепна; 22 - вилична; 23 - зубна; 24 - стремінце; 25 - ковадло; 26 - молоточок;
27 - барабанна кістка; 28, 29 - залишки гіоїда і зябрових дуг утворюють під'язикову кістку і хрящі гортані; 30 - щитоподібний хрящ; 31 - черпакуватий хрящ; 32 - шилоподібний відросток - залишок гіоїда, який приростає до виличної кістки, I-IV - вісцеральні дуги
У черепній коробці чотири потиличні кістки зливаються в загальну потиличну кістку (occipitale); вона оточує великий потиличний отвір і утворює два потиличні виростки для зчленування з хребетним стовпом. Вушні кістки зростаються в парну (праву і ліву) кам’янисту кістку (petrosum) (рис. 5.50, 6). Дно черепа утворюють непарні основна (basisphenoideum) і передня (praesphenoideum) клиноподібна, а попереду них у нюховій частині розвивається непарна решітчаста кістка (ethmoideum).
Міжочноямкову перегородку і передньонижню частину мозкової коробки утворюють парні основні кістки: очноямково-клиноподібна (orbitosphenoideum) і крило- клиноподібна (alisphenoideum).
Склепіння черепа формують парні покривні кістки: носові, слізні, лобові, тім’яні й непарна міжтім’яна. Значну частину бічної стінки мозкової коробки утворюють парні покривні лускаті кістки, від кожної з яких відходить потужний виличний відросток, що з’єднується з виличною кісткою, яка, відповідно, приростає до верхньощелепної кістки. Утворена так вилична дуга обмежує очну ямку знизу. Дно черепа укріплюють парні покривні кістки: піднебінні та крилоподібні, - і непарний маленький леміш. Міжщелепні кістки у багатьох видів зливаються з верхньощелепними. Ділянка середнього вуха прикрита специфічною для ссавців барабанною кісткою (tympanicum), очевидно, утвореною з кутової кістки нижньої щелепи предків.
Для всіх ссавців характерне утворення кісткового піднебіння, що відділяє носовий хід від ротової порожнини; його формують піднебінні кістки і піднебінні відростки між- і верхньощелепних кісток. Завдяки кістковому піднебінню і його дальшій частині - м’якому піднебінню (сполучнотканинній перетинці) - отвори хоан зміщуються до гортані, що дає змогу дихати, коли ротова порожнина наповнена їжею. Шви між кістками ссавців, на відміну від швів у птахів, зберігаються все життя. Зуби гетеродонтні, розміщені в альвеолах.
Плечовий пояс спрощений і з’єднаний з осьовим скелетом тільки м’язами та зв’язками. Добре розвинена велика лопатка, на зовнішньому боці якої є гребінь (збільшення поверхні для прикріплення м’язів). Коракоїд зменшений і приростає до лопатки у вигляді коракоїдного відростка (залишається як самостійна кістка лише в однопрохідних). Ключиця є лише у тих ссавців, у яких передні кінцівки зберегли можливість рухів у різних площинах (кроти, кажани, примати, ведмеді, коти й ін.), а у решти вона зникає (псові, копитні), оскільки у них передні кінцівки рухаються лише в площині, що паралельна до площини тіла. Тазовий пояс складається з двох безіменних кісток, утворених злиттям клубових, лобкових і сідничних. Таз закритий: лобкові й сідничні кістки лівого та правого боків зростаються одна з одною по середній лінії (так утворюють симфіз).
Скелет парних кінцівок зберігає типові для наземних хребетних риси п’ятипалої кінцівки. На відміну від плазунів і подібно до земноводних, у ссавців на передній кінцівці рухомий суглоб розташований між кістками передпліччя і проксимальним рядом кісток зап’ястка, а на задній - між гомілкою та проксимальним рядом кісток плесна (гомілковостопний). Вихідний тип будови кінцівок у різних рядах ссавців унаслідок пристосування до різного типу пересування набув суттєвих змін: змінилися відносна довжина відділів кінцівок, конфігурація і товщина кісток, зменшилася кількість пальців тощо (рис. 5.51).
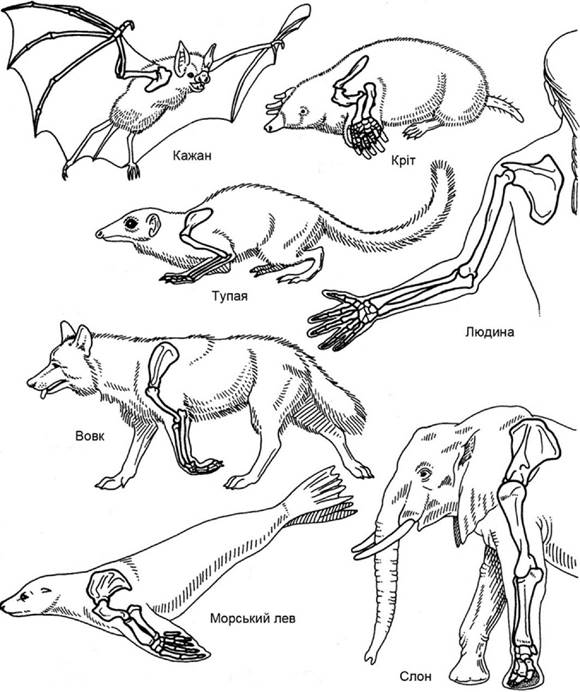
Рис. 5.51. Скелети кінцівок різних ссавців
У кажанів незвично видовжені фаланги 2-5 пальців підтримують натягнуту перетинку, утворюючи крило. Передні лапи крота - це справжнє землерийне знаряддя; п’ясток і плесно мавп пристосовані для хапання, а задні кінцівки кенгуру і тушканчиків - до стрибків; однопалі ноги коня - до швидкого бігу по ущільненому ґрунті, а ласти китоподібних і сирен з укороченням відділів і збільшенням кількості фаланг нагадують плавець кистеперої риби. Видовження кінцівок, що забезпечує пришвидшення рухів тварин, звичайно досягається видовженням кісток плесна і п’ястка (рис. 5.52). Перехід від стопо- до пальцеходіння, а в копитних - і до фалангоходіння, пришвидшує пересування.

Рис. 5.52. Видовження кінцівок за рахунок підняття і видовження кісток стопи (затемнені):
А - мавпа (павіан), стопо- ходіння; Б - собака, паль- цеходіння; В - лама, фа- лангоходіння
У ссавців удосконалена гістологічна структура кістки. Трубчасті кістки кінцівок з найбільшим навантаженням в основній частині (діафізі) мають чітко виражену щільну пошарову (зонарну) будову. Їхні стінки збудовані з остеонів - укладених одна в одну трубочок, що складаються з кісткових пластинок, скріплених щільними
пучками колагенових фібрил. Між ними розташовані кісткові клітини, а у внутрішній порожнині такого остеона, у гаверсовому каналі проходить нерв і кровоносна судина, що забезпечує клітини кістки поживними речовинами і киснем (рис. 5.53). Проміжки між остеонами заповнені вставними кістковими пластинками. Кістка такої структури відрізняється незначною масою і великою міцністю. На поверхні вона має окістя (перистом) - щільну волокнисту сполучнотканинну оболонку, у якій проходять судини і нерви, які потім проникають у гаверсові канали. Внутрішня порожнина трубчастої кістки заповнена кістковим мозком - органом кровотворення. Верхні та нижні кінці трубчастої кістки - епіфізи - утворені з губчастої маси, що сформована складною системою поперечин і пластин, розміщених уздовж лінії найбільшого динамічного напруження, стискання й розтягування (рис. 5.54). Міцність кістки залежить від ступеня її мінералізації - частки солей кальцію, якими просякнута кісткова тканина. Вона найбільша в кістках, що витримують максимальне механічне навантаження. Ступінь мінералізації відрізняється у різних тварин: у китоподібних унаслідок відносної невагомості тіла у воді в скелеті значно менше мінеральних солей, ніж у скелеті наземних ссавців.
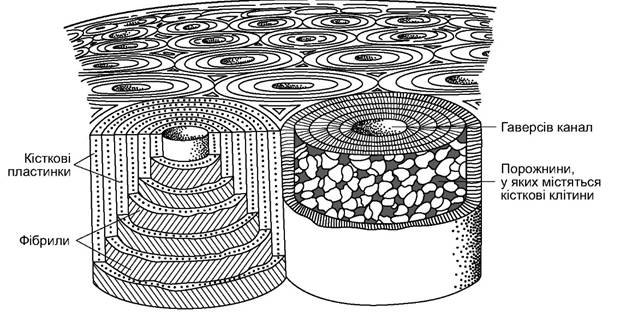
Рис. 5.53. Схема будови щільної речовини кістки. Два остеони: ліворуч - кісткові пластини і фібрили; праворуч - кісткові клітини
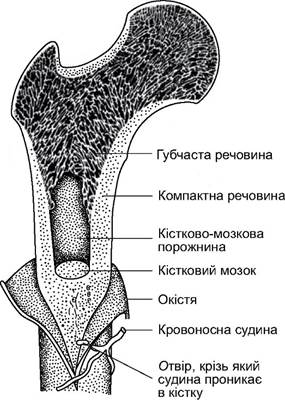
Рис. 5.54. Схема будови трубчастої кістки
М’язи ссавців високодиференційовані і суттєво відрізняється в різних рядах і родинах залежно від способів пересування. Високого розвитку і диференціювання досягла жуйні м’язи, пов’язана з захопленням і механічною обробкою їжі. Складна підшкірна м’язова тканина бере участь не тільки в терморегуляції (зміна положення волосяного покриву, згортання тіла в клубок), а й у спілкуванні тварин: вона забезпечує рухи дотикових волосин (вібрисів) і відповідає за міміку, яка відіграє важливу роль у передаванні інформації, особливо у псоподібних і приматів. Куполоподібна діафрагма - м’язова стінка - розділяє грудну і черевну порожнини; її виникнення дало змогу різко інтенсифікувати вентиляцію легень. У багатьох м’язах ссавців міститься міоглобін (гемоглобін м’язів), що забезпечує резерв кисню. Його кількість максимальна в серцевому та скелетних м’язах, що напружено працюють. Найбільший вміст міоглобіну у водних ссавців, що дає їм змогу довго залишатися під водою. У наземних ссавців (копитні, хижі, мишоподібні) кількість міоглобіну становить до 15-25% від усього гемоглобіну (кров + тканини тіла), а у китоподібних частка міоглобіну відповідає приблизно половині всього гемоглобіну. У кашалота в м’язах міоглобіну у 8-9 разів більше, ніж у наземних ссавців.
Рухи ссавців різноманітні. Швидкість пересування, як зазначено, залежить від довжини кінцівок і частоти кроків. Переміщення кінцівок під тіло, коли ліктьове зчленування обернене назад, а колінне - вперед, збільшило розмір кроку й ефективність роботи м’язів, оскільки різко зменшило витрати зусиль на підтримання тіла над ґрунтом. Перехід від стопоходіння до опори на кінці пальців не тільки сприяв видовженню кінцівки, а й збільшив відштовхувальну силу. Для прискорення руху важливим є збереження гнучкості хребта.
Із багатьох способів пересування поширені: галоп, під час якого задні ноги у стрибках заносяться далеко наперед від передніх (копитні, зайцеподібні, багато мишоподібних та ін.); крок і рись - пришвидшене переставляння кінцівок по діагоналі або попарно лівого і правого боків (інохідь). Двоноге пересування трапляється серед приматів; у тушканчиків і кенгуру воно набуває характеру рикошетного галопу на задніх кінцівках. Деревні форми лазять і стрибають. Плаваючи, водні ссавці (хохуля, видра, ондатра та ін.) працюють задніми кінцівками і частково хвостом. У китоподібних у воді інтенсивно вигинається тіло, а поштовхи відбуваються за участю хвоста; грудні плавці відіграють роль стабілізаторів. Подібно плавають і тюленеві, однак поштовх роблять задніми ластами. Вухаті тюлені на суші пересуваються порівняно легко, спираючись на передні й задні ласти, а справжні тюлені на це не здатні, завдяки особливостям будови скелета нижнього поясу кінцівок (рис. 5.55). Нарешті, кажани віртуозно опанували політ.

Рис. 5.55. Особливості локомоції ссавців:
І - типові форми руху: а - крок, б - дві фази галопу. А - кінь; Б - гепард; ІІ - рикошетний біг рівнинних жителів: В - південна піщанка;
Г- тушканчик; Д - великий сірий кенгуру; ІІІ - фази плавання дельфіна афаліни; IV - пересування на суші ластоногих: Е - звичайний тюлень; Ж - морський котик; З — морж
Швидкість пересування дуже різна. Землерийки, полівки (нориці) розвивають швидкість 4-7, слон і кріль - до 40, морський котик під час плавання - до 27 км/год. Деякі антилопи здатні пересуватися зі швидкістю до 80 км/год, таку ж швидкість, однак на коротких дистанціях, розвиває й лев. Найшвидшим ссавцем уважають гепарда: переслідуючи здобич, від досягає швидкості 105-115 км/год (розгін до 75 км/год за 2 секунди). Кажани в польоті рухаються зі швидкістю 25-30 км /год, а найліпші літуни - бульдогові кажани - до 60 км/год.
Травна система. Живлення ссавців дуже різноманітне. Вегетативні частини деревних, чагарникових і трав’янистих рослин слугують основою харчування копитних, хоботних, зайцеподібних і мишоподібних (полівки, бабаки, ховрахи, бобри); мишоподібні (особливо тушканчики, цокори, сліпаки та ін.) охоче споживають підземні частини рослин. Водними рослинами живляться сирени. Насіння і плоди є кормом для мишей, білок, бурундуків, сонь; насіння і плоди у великих кількостях поїдають ведмеді, олені, кабани, куниці. Чисельність соболів визначена не тільки забезпеченістю тваринними кормами (дрібними гризунами), а й урожаєм кедрових горішків та інших плодів. Плодами живляться крилани (Megachiroptera). Окремі спеціалізовані групи кажанів і деякі сумчасті живляться нектаром квітів, а південноамериканські кажани-вампіри ссуть кров великих за розмірами ссавців. Безхребетні становлять корм для більшості мідицеподібних і кажанів (крім криланів), а також для мишей і тушканчиків, ховрахів і білок, дрібних і навіть великих псоподібних (ведмеді). Морський планктон є основним кормом найбільших сучасних ссавців - вусатих китів. Серед псоподібних ссавців лише деякі живуть винятково з великої живої здобичі (переважно котові), тоді як більшість урізноманітнює раціон рослинними кормами. Псоподібні нерідко нападають на здобич, більшу від них, і успішно добувають її, використовуючи індивідуальні переваги - силу, раптовість нападу, спритність (мустелові, леопарди та ін.), або полюючи групами (вовки, леви).
Еволюційно виникла харчова спеціалізація, яка не тільки позначилася на будові (рис. 5.56) та функціях організму різних ссавців, а й визначила особливості їхньої поведінки, використання території, утворення і характер угруповань (стада копитних, колонії ховрахів, бабаків) і загальну рухливість. Як і у птахів, потреба в їжі у ссавців пов’язана з їхньою теплокровністю. Оскільки ж температура тіла ссавців нижча, ніж температура птахів, то вони потребують порівняно меншу кількість їжі. Найдрібніші тварини (наприклад, землерийка-крихітка масою 1,5-2,5 г) з’їдають за добу їжі у 2-4 рази більше від маси тіла, перериваючи періоди живлення сном; без корму вони не можуть прожити більше 5-8 год. Більші за розмірами ссавці споживають порівняно меншу кількість їжі, живляться у певний час доби і мають більш або менш тривалий нічний чи денний відпочинок. Ритми добової активності ссавців певною мірою визначені харчовою спеціалізацією.

Рис. 5.56. Відмінності у зовнішньому вигляді ссавців, пов'язані з особливостями живлення.
Парнокопитні: А - жирафа (живиться листям дерев); Б - бізон (живиться трав'янистою рослинністю).
Хижі: В - рись (підкрадається і вичікує здобич); Г- вовк (вистежує і переслідує здобич); Д - ласка (ловить дрібних гризунів у їхніх норах)
Добування, збирання і механічна обробка їжі відбуваються за допомогою ротового апарата - губ, язика, щелеп і зубів, а також кінцівок, а у хоботних - хобота. Форма черепа і співвідношення його основних частин - щелеп, носової частини, оч- ниць і мозкової порожнини - визначені розвитком органів чуття, мозку й щелепного апарата і тісно пов’язані зі способом добування та складом їжі. Наприклад, є відмінності в будові губ у двох видів носорогів: у білого носорога розширена морда з широкими губами сприяє живленню трав’яною рослинністю, а у чорного носорога, який поїдає головно гілки дерев і чагарників, морда трохи загострена, а на верхній губі розвинутий пальцеподібний хапальний придаток (рис. 5.57, A, Б). Вузька морда, довгий з китицею на кінці язик у нектароїдної летючої миші (див. рис. 5.57, В) або витягнута морда і довгий клейкий язик мурахоїдів, ящурів, трубкозубів - приклади подібних адаптацій.

Рис. 5.57. Форма губ у чорного (А) і білого (Б) носорогів. Вузька морда і довгий з китицею язик нектароїд- ного кажана Glossophaga (В)
Травний тракт ссавців відрізняється більшою, ніж у плазунів і птахів, довжиною та диференціюванням його відділів і різноманітністю травних залоз. Саме це забезпечує високоефективне використання різноманітної їжі. Цьому сприяє також максимальний розвиток серед хребетних тварин так званого симбіотичного травлення: використання бактерій, грибів і найпростіших кишкової трубки для перетравлювання рослинних, багатих на клітковину кормів, що їх не можуть розщепити і гідролізувати свої ферменти. Ці симбіонти слугують і додатковою поживою для хазяїна (див. нижче).
Ротова порожнина починається переддвер’ям - порожниною між м’ясистими губами (які є тільки у ссавців), щоками і щелепами. У хом’яків, бурундуків і мавп тут утворені защічні мішки, куди тварини збирають корм і потім переносять у сховища. Губи звичайно мають дотикові клітини. М’ясистих губ немає у однопрохідних і китоподібних.
Щелепи сучасних ссавців мають зуби різної форми і призначення. Зуби поділяють на різці (incisivi), ікла (canini), передкутні (praemolares) і кутні (molares). Різці, ікла та передкутні мають дві генерації (молочні замінюються на постійні), кутні - тільки одну. У тюленевих і зубатих китів зуби недиференційовані. Гетеродонтність ссавців - важливе пристосування, що вдосконалює живлення і травлення.
Різні ряди ссавців відрізняються за складом і будовою зубів. Вихідним, очевидно, був суцільний ряд із порівняно слабко диференційованих зубів. Частково він зберігся у мідицеподібних, кажанів, частково у приматів і псоподібних. Однак у кожній групі він зазнав адаптивних змін. У дрібних мідицеподібних (землерийок), що мають витягнуту вузьку морду, зубні ряди з витягнутими вперед різцями утворили своєрідний пінцет, яким зручно захоплювати й утримувати дрібну здобич (комах, червів та ін.), а горбкуватими, що мають гострі верхівки, кутніми зубами трощити хітиновий покрив. Зубний ряд псоподібних зазнав більшого диференціювання:
гострі різці, великі ікла й кутні з ріжучими краями. Ікла як засіб захоплення і придушення здобичі, досягли найбільшого розвитку у вимерлого шаблезубого тигра.
У мавп живлення переважно м’якими плодами призвело до слабшого диференціювання зубного ряду: ікла невеликі, а кутні мають горбкуваті або плоскі жуйні поверхні. У мишоподібних різці з гострими ріжучими краями; кількість різців менша (по два у верхній і нижній щелепі), ікла зникли, а корінні набули горбкуватої або плоскої жуйної поверхні з перетинками (валиками) з емалі; така поверхня здатна перемелювати найгрубші частини рослин. У мишоподібних різці, а у частини видів і корінні ростуть протягом усього життя. У слонів є лише пара різців на верхній щелепі (бивні) та по одному великому справжньому корінному у верхній і нижній щелепах; загальна кількість зубів - лише шість. У інших ссавців їх більше. Для з’ясування кількості й видової ідентифікації застосовують зубні формули, у яких відображена кількість зубів у половині верхньої та нижньої щелеп. Типи зубів позначають початковими літерами їхньої латинської назви. Зубна формула виглядає так:
кабан І 3/3 С 1/1 РМ 4/4 М 3/3=44 зуби;
заєць І 2/1 С 0/0 РМ 3/2 М 3/1=28 зубів.
У вусатих китів зуби закладаються в ембріональному стані, однак пізніше зникають. Одночасно по боках ротової порожнини, уздовж зовнішніх країв верхніх щелеп утворюються епідермальні валики, які пізніше розростаються, роговіють і розділяються на ряд трикутних пластин, основи яких занурені у тканину ясен. Кількість пластин у різних видів коливається від 160 до 500. Ці пластини, які називають китовим вусом, несуть китиці з переплетених рогових волокон і слугують для відціджування планктонних організмів: зімкнувши щелепи і піднявши язик, кит витісняє воду з ротової порожнини.
Рогові валики на піднебінні мишоподібних і копитних беруть участь у перетиранні рослинної їжі. На дні ротової порожнини лежить м’ясистий язик, що бере участь у пережовуванні й ковтанні їжі, а в копитних - і у збиранні їжі. У котових рогові сосочки на язику дають змогу здирати м’ясо з кісток жертви. Довгий, червоподібний і клейкий язик мурахоїдів - знаряддя для добування мурашок і термітів із їхніх гнізд (завдовжки 60 см і кріпиться до грудини). Слизові виділення трьох пар великих і кількох дрібних слинних залоз, що відкриваються в ротову порожнину, утворюють слину, в якій містяться ферменти, що розщеплюють крохмаль уже під час пережовування їжі. У слині рослиноїдних тварин їх більше, ніж у м’ясоїдних. У слизовій ротової порожнини і язика розміщені смакові сосочки. Пережована й достатньо змочена слиною їжа з ротової порожнини через глотку і стравохід потрапляє у шлунок. Стравохід більшості ссавців - це тонкостінна трубка з гладенькими м’язами. Лише у жуйних копитних він має поперечно-посмуговані м’язи, за їхньою допомогою їжа (жуйка) зі шлунка самовільно відригується назад у ротову порожнину для додаткового пережовування. Шлунок відокремлений від стравоходу і кишечника й добре розвинений у всіх ссавців. Його розміри, форма і будова змінюються в різних рядах (рис. 5.58). У однопрохідних шлунок має форму простого мішка і не має травних залоз. Однокамерний шлунок псоподібних, приматів і мідицеподібних за формою нагадує реторту, епітелій його стінок має численні травні залози. У живленні жуйних переважають грубі рослинні корми, тому їх шлунок складається з рубця, сітки, книжки і сичуга (рис. 5.59): перші три відділи (рубець, сітка і книжка) утворюють так званий передшлунок і вистелені багатошаровим епітелієм; у передшлунку немає травних залоз і в ньому відбувається лише бактеріальне бродіння за участю симбіонтів, які можуть існувати лише в нейтральному або слабколужному середовищі. Розщеплення рослинної їжі симбіонтами відбувається в рубці, де накопичується лише злегка пережована їжа; бродіння посилюється після повторного пережовування жуйки і змочування її слиною, яка має слабколуж- ну реакцію. У сітці та книжці продовжується бродіння і механічне перетирання їжі. Обробка шлунковим соком відбувається тільки в сичузі, який має кисле середовище.
Рис. 5.58. Схема будови шлунків ссавців:
А - єхидна; Б - кенгуру; В - людина; Г - заєць; Д - даман; Е - дельфін; Ж - бик:
1 - багатошаровий епітелій стравоходу; 2-4 - одношаровий епітелій з кардіальними (2), фундальними (3) та пілоричними (4) залозами
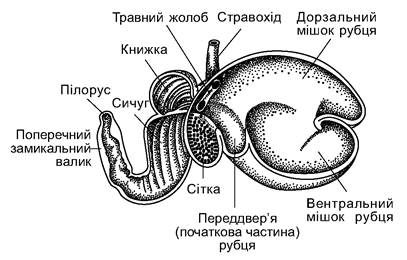
Рис. 5.59. Шлунок жуйного ссавця (вівці):
1 - стравохід; 2 - травний жолоб; 3 - дорзальний мішок рубця; 4 - вентральний мішок рубця; 5 - переддвер'я (початкова частина) рубця; 6 - сітка; 7 - книжка; 8 - сичуг; 9 - пілорус; 10 - поперечний замикальний валик
У лінивців шлунок слугує переважно місцем накопичення їжі; він сягає 20-30% від маси тіла і завжди заповнений їжею, яка повільно надходить у кишечник і просувається по ньому з малою швидкістю (інколи до тижня). Складний шлунок китоподібних, зуби яких не можуть подрібнювати їжі, забезпечує її механічну обробку під час перистальтичних рухів стінок. У мурахоїдів подрібнення їжі відбувається в пілоричній частині шлунка, яка має тверду кутикулярну вистилку зі сильно розвиненими м’язами. Шлунковий сік китоподібних і мурахоїдів вирізняється активністю і розкладає (гідролізує) навіть такі стійкі речовини, як хітин (інші тварини його зазвичай не перетравлюють).
Травні залози шлунка неоднакові. У найближчому до стравоходу відділі є кардіальні залози розгалуженої форми; у середньому відділі домінують важливі для травлення фундальні залози, які слабко галузяться, а в прилеглому до кишечника пілоричному - пілоричні залози. Шлунковий сік, що його виділяють залози, містить соляну кислоту (до 0,4-0,5%) і ферменти - пепсин (розкладає білки), ліпазу (розкладає жири) й деякі інші.
Кишечник ссавців (рис. 5.60) розділений на тонку, товсту і пряму кишки; початкову частину тонкої кишки називають дванадцятипалою кишкою; на межі тонкого і товстого відділів відходить велика сліпа кишка, що зменшується в розмірах або зовсім зникає у небагатьох м’ясоїдних тварин. Як і у шлунку, стінки в кишечнику складаються з трьох шарів: внутрішнього слизистого з травними залозами, м’язового з поздовжньо і поперечно розташованими гладкими м’язовими волокнами і зовнішньої серозної оболонки. Залози стінок тонкої кишки виділяють різноманітні травні ферменти; тут же відбувається перетравлювання і всмоктування основних поживних речовин. У товстий відділ кишечника надходить розріджена їжа з важкими для перетравлювання частинками. Там і особливо у сліпій кишці розвиваються бродильні процеси за участю бактерій, грибків і найпростіших, аналогічно до процесів у складному шлунку жуйних. У прямій кишці відбувається зворотне всмоктування води і формування калових мас.
У тварин, що живляться рослинними кормами, виникає проблема отримання незамінних амінокислот, які містяться лише у тваринних білках. Вивіркові (вивірки, ховрахи та ін.) вирішують цю проблему, енергійно поїдаючи дрібних тварин. Хом’яки та полівки, шлунок яких не має травних залоз і вистелений кутикулою ніби для захисту стінок від пошкоджень грубими кормами, використовують шлунок як бродильний чан, де розмножуються найпростіші, здатні розкладати клітковину. Потрапляючи з харчовою масою в кишечник, вони перетравлюються і дають хазяїнові необхідні білки. Ще більших масштабів аналогічні процеси набули у складних шлунках копитних, яких можна навіть назвати «протистоїдами», оскільки вони використовують інфузорій та інших найпростіших, що розмножуються у рубці, сітці та книжці. Зайцеподібні як бродильні чани мають лише сліпу і товсту кишки, проте симбіонти, які там живуть, не можуть бути перетравлені, бо в цих відділах немає потрібних ферментів (пепсину) і кислого середовища. У них виникає явище копрофагії (поїдання калових мас), оскільки лише так можуть
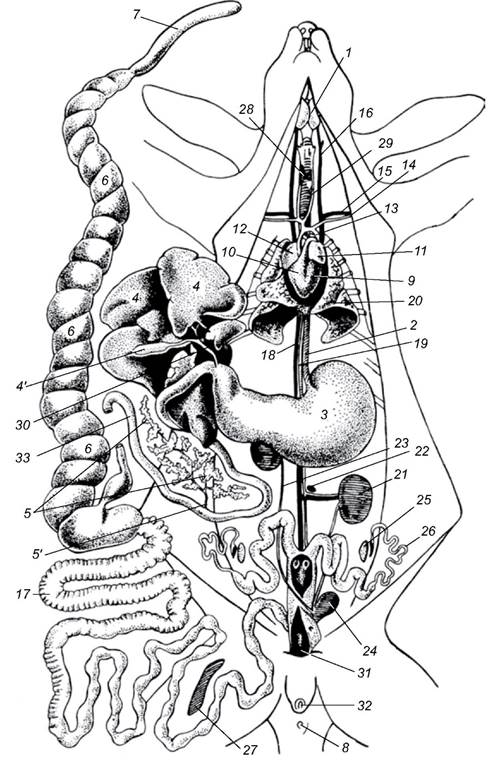
Рис. 5.60. Загальне розташування внутрішніх органів самки кроля:
1 - підщелепна слинна залоза; 2 - стравохід; 3 - шлунок; 4 - печінка (відкинута догори); 4' - жовчний міхур; 5 - підшлункова залоза;
5' - протока підшлункової залози; 6 - сліпа кишка; 7- її червоподібний відросток; 8 - анальний отвір; 9 - лівий шлуночок серця;
10 - правий шлуночок серця; 11 - ліве передсердя; 12 - праве передсердя; 13 - дуга аорти; 14 - ліва підключична артерія; 15 - ліва підключична вена; 16 - ліва яремна вена; 17 - товста кишка; 18 - задня порожниста вена; 19 - аорта; 20 - ліва легеня; 21 - ліва нирка; 22 - ліва надниркова залоза; 23 - правий сечовід; 24 - сечовий міхур; 25 - лівий яєчник; 26 - лівий яйцепровід; 27 - селезінка;
28 - щитоподібна залоза; 29 - трахея; 30 - жовчна протока; 31 - піхва; 32 - статевий отвір; 33 - дванадцятипала кишка
бути перетравлені тварини - симбіонти кишечника. У цьому разі відбувається поїдання екскрементів, найбагатших на тваринні білки.
Довжина кишечника і співвідношення його відділів у різних ссавців відповідають складу їхньої їжі. У кажанів кишечник довший від тіла в 1,5—4,0 рази, у мідицеподібних — у 2,5—4,5, у вовка — у 6,5, у мишоподібних (піщанок, морських свинок) — у 5—12, коня — у 12, а у вівці — у 29 разів довший від тіла. Живлення багатими на клітковину кормами завжди супроводжується не лише загальним видовженням кишечника, а й збільшенням відносних розмірів товстих і особливо сліпої кишки.
Травні залози — печінка і підшлункова залоза — беруть участь не лише у перетравлюванні, виробляючи активні ферменти, а й у загальних обмінних і видільних процесах, а також у їхньому гормональному регулюванні. Протоки печінки (жовчна протока) і підшлункової залози впадають у передню частину тонкої кишки — дванадцятипалу кишку. Відносний розмір печінки зменшується зі збільшенням розмірів тіла. Наприклад, печінка землерийки масою 18—20 г становить 5—6% маси тіла, а печінка кита — лише 1%. Підшлункова залоза збільшує розміри і секреторну активність зі збільшенням частки рослинних кормів.
Низка специфічних пристосувань забезпечує існування ссавців у разі сезонних змін доступних кормів. Частина цих пристосувань — спільна з іншими тваринами. Зокрема, накопичення запасів резервних речовин (жиру в підшкірній клітковині й порожнині тіла, глікогену в печінці) у сприятливі сезони (жирування) та їхнє використання в несприятливі періоди. У деяких ссавців маса жирових запасів може сягати 40% від маси тіла. У багатьох видів чітко виражена зміна кормів. Наприклад, лосі, зайці та багато інших травоїдних, які влітку живляться зеленими частинами рослин, узимку поїдають суху траву, пагони, кору дерев і чагарників. Для деяких ссавців характерне запасання їжі, подібне до запасання у суспільних комах.
У несприятливий період багато ссавців впадає у стан заціпеніння (сплячку), що є альтернативою збереження активності завдяки запасанню їжі. Сплячка пов’язана або з нестачею, або з недоступністю кормів; звичайно припадає на період літньої посухи; не простежується у тропічних лісах із достатніми запасами їжі й у тундрі, де тривалість несприятливого періоду дуже велика. Зафіксовано, що в разі особливо несприятливих умов — у сильні морози і заметілі — по кілька діб не покидають сховища, відсипаючись у гніздах, куниці, вивірки й деякі інші види.
До періоду сплячки організм готується. Відбувається перебудова багатьох фізі- олого-біохімічних процесів під впливом і контролем нейрогуморальної системи, яка відображає ендогенні ритми і реагує на сезонні зміни умов життя, передусім — погіршення можливостей добування їжі. Такий механізм забезпечує попередню підготовку організму і дає змогу пристосовуватися до конкретних умов року. Зміни поведінки і характеру обміну речовин забезпечують накопичення перед сплячкою значних запасів жиру в підшкірній клітковині й порожнині тіла, аскорбінової кислоти (вітамін С) у тканинах, глікогену і вітаміну Е в печінці. Тварини шукають або будують сховища, у яких вони проводитимуть сплячку: у печерах — кажани, у дуплах — вовчки, у барлогах — ведмеді, у норах — різні мишоподібні тощо.
Є декілька типів сплячки, а саме:
1) зимовий сон властивий ведмедю, борсуку, єноту, єноту уссурійському. У звірів під час сну зменшується інтенсивність дихання і кровообігу, температура тіла знижується лише на 2-7°С, а загальний рівень метаболізму - на 50-70% (Слоним, 1961). Потурбовані звірі легко і швидко прокидаються; переривається сон і за умов тривалих відлиг. Самки ведмедів під час зимового сну народжують малят. Тривалість зимового сну визначена погодними умовами і доступністю їжі. Наприклад, бурі ведмеді в середній смузі лежать у барлозі з листопада по квітень, а на Кавказі - лише з грудня по лютий (в деяких районах у теплі безсніжні зими взагалі не сплять);
2) справжня сплячка, що переривається з підвищенням температури. Тварини впадають у заціпеніння, що супроводжується значним зменшенням інтенсивності дихання і кровообігу, зниженням температури тіла, втрачають здатність реагувати на подразники (звуки, дотик тощо). Повільно прокидаються лише за умов довготривалих відлиг. Зокрема, у цей період кажани ловлять у печерах комах, хом’яки і бурундуки використовують зібрані запаси їжі;
3) справжня безперервна сплячка супроводжується глибшим заціпенінням, за якого інтенсивність дихання і кровообігу зменшується в 1020 разів, обмін речовин знижується у 20-40 разів, а температура тіла - до 5-1°С (Калабухов, 1956; Слоним, 1961). Найрізкіші зовнішні подразники (звуки, біль тощо) і короткочасні підвищення температури не призводять до пробудження. За період сплячки маса тіла знижується на 30-40%, причому передусім використовуються запаси жиру. Така сплячка характерна для їжаків, частини кажанів і багатьох мишоподібних (бабаки, ховрахи, тушканчики, соні). З висиханням рослинності ховрахи впадають у заціпеніння ще влітку; літня сплячка переходить у зимову, і тварина стає активною лише наступної весни, провівши у стані заціпеніння 6-8 місяців.
Дихальна система. У газообміні ссавців провідну роль відіграють легені, а також у ньому бере участь слизова поверхня дихальних шляхів і частково шкіра; крізь шкіру потрапляє лише 1% необхідного тварині кисню. Носова порожнина розділена на переддвер’я, дихальний і нюховий відділи. Функція переддвер’я - вловлювання порівняно великих часток (пилу), що проникають з повітрям. Переддвер’я більше розвинене у жителів степових і пустельних районів, де повітря особливо запилене. Дрібні часточки пилу вловлюються у дихальному відділі носової порожнини, яка вистелена слизовою оболонкою з миготливим епітелієм, що має багато кровоносних капілярів, тут відбувається не тільки подальше очищення повітря від механічних домішок, а й також його знезаражування бактерицидно активними речовинами слизу; одночасно під час проходження між складками носової раковини повітря зволожується і зігрівається кров’ю, яка проходить по капілярах, віддаючи частину кисню. Нюховий відділ, розширений виростами бічних стінок, утворює лабіринт порожнин, вистелених нюховим епітелієм. Проходячи крізь хоани у глотку, повітря потрапляє в гортань. В основі гортані є кільцеподібний перснеподібний хрящ, характерний для всіх наземних хребетних. Передню і бічні стінки гортані утворює наявний тільки у ссавців щитоподібний хрящ. черпакуваті хрящі розташовані з боків спинної частини гортані. До переднього краю щитоподібного хряща прилягає тонкий надгортанник (рис. 5.61), який прикриває вхід у гортань під час проходження їжі через глотку. Голосові зв’язки - парні складки слизової оболонки, розташовані між щитоподібним і черпакуватим хрящами. Голоси більшості ссавців обмежені переважно низькочастотним діапазоном, що, однак, не перешкоджає можливості передавання ними складної та місткої інформації.
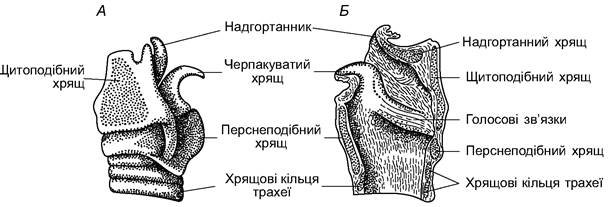
Рис. 5.61. Гортань корови: А - вигляд збоку; Б - сагітальний переріз
До гортані прилягає трахея, яку підтримують хрящові кільця. Трахея розділена на два бронхи, що відрізняються від неї лише меншим діаметром; у легенях бронхи діляться на ще дрібніші трубочки, утворюючи бронхіальне дерево (рис. 5.62). Най- дрібніші трубочки - бронхіоли - відкриваються в розширення, вистелені дихальним епітелієм - альвеоли розміром 25-400 мкм, у стінках яких галузяться численні капіляри. Така структура легень збільшує їхню дихальну поверхню, яка в 50-100 разів більша від поверхні тіла. Відносний розмір поверхні, через яку в легенях відбувається газообмін, більша у тварин із високою активністю і рухливістю.
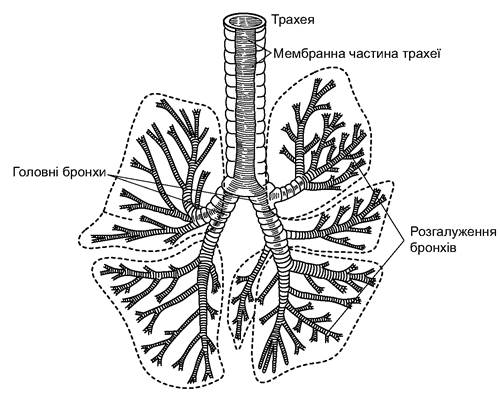
Рис. 5.62. Розгалуження бронхів у легенях ведмедя бурого
Відносне збільшення легень простежується у високогірних і водних ссавців. У китів є кільцеподібні м’язи, яка замикає вхід в альвеоли і дає змогу затримувати в них повітря навіть на значних глибинах.
Механізм дихання ссавців подвійний. У разі так званого реберного дихання за допомогою міжреберних м’язів змінюється об’єм грудної клітки; у разі діафрагмального - той же об’єм змінюється шляхом опускання і підіймання м’язової грудинно-черевної перепони - діафрагми. У різному поєднанні обидва механізми функціонують у всіх ссавців; у псоподібних переважає перший механізм, у копитних - другий. Кількість подихів у дрібних тварин із вищим рівнем метаболізму вища порівняно з великими; у коня вона дорівнює 8-16, у щура - 100-150, у миші - 200 за хвилину. Під час пересування частота дихання зростає. Дихання бере участь у терморегуляції. Часте, однак неглибоке дихання псоподібних (поліпное) збільшує випаровування з поверхні верхніх дихальних шляхів, сприяє віддачі тепла. Прискорення глибокого дихання, посилюючи газообмін у легенях, збільшує теплопродукцію організму за низьких температур.
Кровоносна система. У ссавців, як і у птахів, велике і мале кола кровообігу відокремлені (роз’єднані). Від лівого шлуночка чотирикамерного серця відходить одна ліва дуга аорти. У більшості видів від неї відділяється коротка безіменна артерія, що розділена на праву підключичну і сонні (праву та ліву) артерії; ліва підключична артерія відходить самостійно. Спинна аорта - продовження лівої дуги - відгалужує судини до м’язів і внутрішніх органів (рис. 5.63).
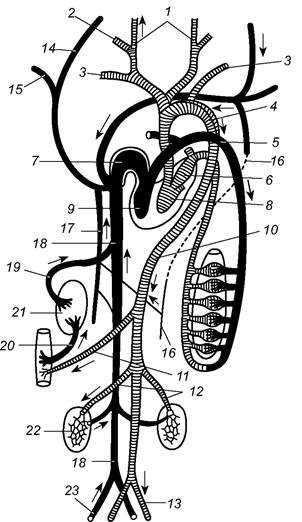
Рис. 5.63. Схема кровоносної системи ссавців:
1, 2 - зовнішня і внутрішня сонні артерії; 3 - підключична артерія; 4 - ліва дуга аорти; 5 - легенева артерія; 6 - ліве передсердя;
7 - праве передсердя; 8 - лівий шлуночок; 9 - правий шлуночок; 10 - спинна аорта; 11 - нутрощева артерія; 12 - ниркова артерія;
13 - клубова артерія; 14 - яремна вена; 15 - підключична вена; 16 - ліва непарна вена; 17 - права непарна вена; 18 - задня порожниста вена; 19 - печінкова вена; 20 - ворітна вена печінки; 21 - печінка; 22 - нирка; 23 - клубова вена
Лише у небагатьох ссавців однаково розвинені обидві порожнисті вени; у більшості видів права передня порожниста вена приймає в себе безіменну вену, утворену злиттям яремної та лівої підключичної вен. Несиметричні і рудименти задніх кардинальних вен нижчих хребетних - так звані непарні (хребетні) вени - характерні тільки для ссавців. У більшості видів ліва непарна вена з’єднана з правою непарною веною, яка впадає у праву передню порожнисту вену. Характерно, що немає ворітної системи нирок; це пов’язано з особливостями видільних процесів.
Лімфатичні судини відкриваються у венозні судини біля серця. Вони починаються лімфатичними капілярами, що збирають міжтканинну рідину (лімфу). У лімфатичній системі ссавців немає лімфатичних сердець (пульсуючі ділянки судин), проте є лімфатичні вузли (залози), функція яких - очищення лімфи від хвороботворних мікроорганізмів за участю фагоцитарних клітин - лімфоцитів. За хімічним складом лімфа подібна до плазми крові, однак бідніша на білки. У лімфатичних судинах, що контактують із травним трактом, лімфа збагачена жирами, молекули яких не можуть проникнути крізь щільні стінки капілярів кровоносних судин, та легко проходять крізь проникніші стінки лімфатичних судин. Форменими елементами лімфи є різні типи лімфоцитів (білих кров’яних тілець).
Кровотворні органи спеціалізовані. Кістковий мозок продукує еритроцити, гранулоцити і тромбоцити; селезінка і лімфатичні залози - лімфоцити; ретикуло-ендотеліальна система - моноцити.
Речовини аглютиніни, лізини, преципітини й антитоксини нейтралізують або знищують шкідливі речовини, що потрапили у кров. Дрібні еритроцити ссавців не мають ядер, що збільшує ефективність перенесення ними кисню, оскільки на власне дихання вони витрачають кисню в 9-13 разів менше від еритроцитів птахів і у 17-19 разів менше від еритроцитів амфібій. Відносний розмір серця більший у рухливіших і дрібніших тварин. У великих за розмірами видів маса серця становить 0,2-0,7% від маси тіла, у дрібних - до 1,0-1,5; у кажанів - 1,3%.
Частота пульсу за хвилину в миші дорівнює 600, у собаки - 140, у бика і слона - 24. У водних тварин кількість серцевих скорочень зменшується після занурення (у тюленя з 180 у надводному стані до 60-30 під водою), що дає змогу економніше використовувати запаси кисню в легенях і повітроносних порожнинах. Одночасно полегшується вирівнювання тиску крові, яке в разі швидкого занурення китоподібних (100-140 м/хв) помітно змінюється за короткий проміжок часу. В усіх водних ссавців різко збільшена кількість міоглобіну в м’язах. Запаси кисню в їхньому організмі розподілені так (%):
Розподіл запасів кисню людини і кита
Легені |
Кров |
М'язи |
Інші органи |
|
Людина |
34 |
41 |
13 |
12 |
Кит |
9 |
41 |
41 |
9 |
Видільна система і водно-сольовий обмін. У ссавців як основний продукт азотистого обміну виводиться сечовина: за цією ознакою ссавці стоять ближче до амфібій. Водно-сольовий обмін відбувається переважно через нирки, його регулюють гормони задньої частки гіпофіза, що впливають на діурез (сечовипускання) і зворотне поглинання (реабсорбцію) води з первинної сечі (фільтрату) в ниркових канальцях. Також у водно-сольовому обміні у ссавців беруть участь як шкіра з її потовими залозами, так і кишкова трубка.
Нирки ссавців, як і інших амніот, метанефричні; мають власні вивідні канали - сечоводи, що впадають у сечовий міхур. Протока сечового міхура відкривається у самців у копулятивний орган, а в самок - у переддвер’я піхви. У клоачних (яйцекладних) вивідні канали впадають у клоаку. Нирки бобоподібні, лежать по боках хребта. На поздовжньому перерізі нирки видно, що вона складається з двох шарів. У зовнішньому (кірковому) шарі розташований фільтрувальний апарат - гломерули. Гломерула - це клубочок кровоносної судини, яку оточує боуменова капсула. Від капсули починається вивідний канал, що має чотири відділи: проксимальний звивистий, петлю Генле, дистальний звивистий і збиральний канал. Усе разом має назву нефрон (рис. 5.64). Збиральні канали є загальними для групи нефронів: так утворюються добре помітні на розрізі піраміди, або частки, нирок. Отвори збиральних каналів відкриваються в ниркові миски, від яких починаються сечовивідні канали.
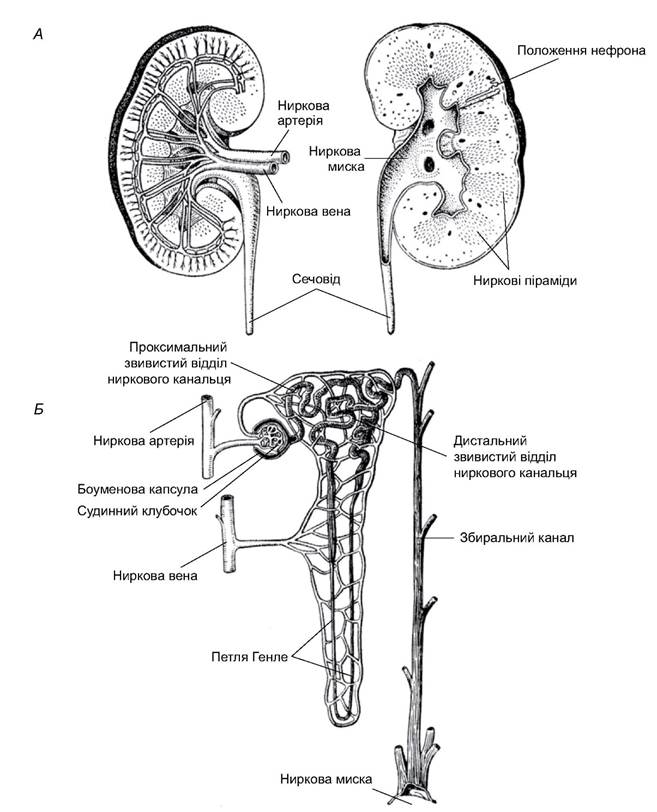
Рис. 5.64. Нирка ссавців: А - поперечний переріз нирки людини; Б - будова нефрона
Фільтрат у боуменовій капсулі представлений плазмою крові без білків, яка ще має значну кількість корисних для організму речовин (цукри, вітаміни, амінокислоти, солі, вода). Вони виводяться двома шляхами - дифузією через стінки різних відділів канальців нефронів, що підлягає дії основних фізичних законів (різниця концентрації в канальцях і навколишній тканині нирки) і активною реабсорбцією за участю ферментів, зв’язаних з мембранами клітин стінок канальців. У проксимальному звивистому відділі відбувається активна реабсорбція цукру, вітамінів, амінокислот, хлоридів і частково іонів натрію. Вода та йони натрію виводяться у петлі Генле, яка сильно розвинена у ссавців. Її частини, по яких ідуть протилежно спрямовані потоки первинної сечі, обмежені тканинами, у міжклітинних рідинах яких осмотичний тиск максимальний біля верхівки петлі (мозковий шар нирки) і мінімальний у кірковому шарі. У мозковому шарі відбувається виведення води, що дифундує в інтерстицій тканини нирки, а звідти - у кровоносні судини, внаслідок чого підвищується концентрація сечі. Виведення йонів натрію відбувається в тому ж відділі за участю ферментних систем (активна реабсорбція). Реабсорбція води й інших речовин дуже велика. Наприклад, людина за добу через ниркові клубочки фільтрує приблизно 180 л (первинна сеча), а виводить з організму за той же час лише 1-2 л (кінцева сеча). У водному балансі деяку роль відіграє і пряма кишка, стінки якої всмоктують воду з калових мас (особливо характерно для напівпустельних і пустельних тварин).
Забезпечені водою жителі зволожених біотопів мають значний водний обмін. Жителі напівпустельних біотопів більшу частину води отримують, поїдаючи соковиті частини сукулентних рослин. Їхні шкірно-легеневі втрати води мінімальні, наприклад, за температури 20°С у порівняно вологолюбного виду тамариксової піщанки - 170 см3, а в жителя посушливих біотопів великої піщанки - лише 50 см3 на 1 кг маси за 1 год. Справжні пустельні ссавці здатні поїдати майже сухі корми і практично не пити протягом усього життя, задовольняючи потреби завдяки метаболічній воді, що утворюється в організмі. Верблюди в багаті на корми і вологі сезони запасають жир, який втрачають у період посухи і недостатньої кількості корму. У процесі використання жиру утворюється певна кількість води. Під час відпочинку і сну верблюди знижують температуру тіла, що також зменшує витрату води.
Статева система і розмноження. Ссавці вирізняються високим ступенем турботи про потомство на всіх етапах його розвитку. Їм властивий розвиток в утробі самки, протягом якого зародок росте завдяки речовинам тіла матері, отримуючи їх через особливий орган - плаценту. Лише однопрохідні ссавці відкладають великі, багаті на жовток яйця.
У всіх ссавців після народження молодої тварини зв’язок з матір’ю не припиняється, оскільки дитина живиться молоком. Однак і після завершення вигодовування молоком зв’язки між батьками і потомством, зазвичай, певний час зберігаються. Це забезпечує можливість навчання молодняку - передавання потомству накопиченого батьками індивідуального досвіду. Такий зв’язок поколінь, який дає змогу накопичувати популяційно-видовий досвід, називають сигнальною спадковістю, або сигнальним передаванням інформації. Це явище поширене серед птахів і особливо характерне для ссавців. Якісно іншого рівня набуває воно у людини з її розумом і другою сигнальною системою, основою діяльності якої є виробничо-суспільні відносини.
Статеві органи ссавців побудовані складніше, ніж у інших амніот (рис. 5.65). Сім’яники парні, розташовані у задній частині черевної порожнини (клоачні, деякі мідицеподібні, неповнозубі, хоботні, китоподібні, сирени, носороги) або перемістились у калитку - шкірястий виріст, що сполучений з порожниною тіла пахвинним каналом (сумчасті, псоподібні, копитні, примати). До сім’яника прилягає витягнуте по осі тіло - придаток сім’яника (epididymis) - скупчення вивідних проток сім’яника (залишок переднього відділу мезонефричної нирки). Від придатка відходить вольфів канал - сім’япровід, що впадає біля основи статевого члена в його сечостатевий канал. Перед впаданням у канал сім’япроводи утворюють часточкові розширення - сім’яні міхурці (vesicula seminalis), секрет яких бере участь в утворенні сперми. У мишоподібних секрет виділяється після викидання сімені у статеві шляхи самки і, застигаючи, закупорює їх після копуляції.
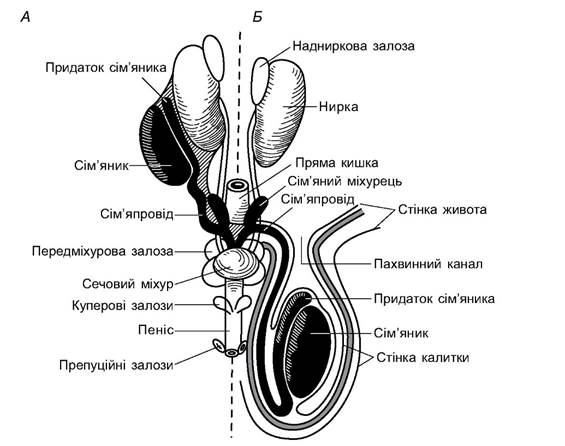
Рис. 5.65. Чоловічі статеві органи ссавців: А - сім'яники, розташовані в черевній порожнині; Б - сім'яники в мошонці
Біля основи статевого члена розміщена передміхурова залоза (prostata), секрет якої утворює рідке середовище сперми. Статевий член (penis) має печеристі тіла, що оточують сечостатевий канал; наповнення їхніх порожнин кров’ю забезпечує пружність копулятивного органа. У деяких ссавців (хижі, тюленеві, мишоподібні та ін.) його міцність збільшує спеціальна кістка (ospenis). Зі статевим членом пов’язані також куперові та препуційні залози; перші беруть участь в утворенні рідкої частини сперми (еякуляту), а другі виділяють пахучий секрет, що сприяє розпізнанню особинами протилежної статі та виникненню статевого збудження партнерів.
Парні яєчники самок завжди лежать у порожнині тіла й прикріплені до спинної стінки черевної порожнини брижами. Розмір яєчників помітно менший, ніж у решти хребетних. Парні яйцепроводи, гомологічні мюллеровим каналам, відкриваються в порожнину тіла біля яєчників лійками, що вистелені миготливим епітелієм. Верхній тонкий і звивистий відділ яйцепроводу називають фаллопієвою трубою, а нижній розширений, з м’язовими стінками - маткою. У клоачних матки відкриті в сечостатевий синус клоаки; у решти ссавців клоака зникла. У сумчастих нижня частина яйцепроводів перетворена в парні піхви, які інколи зростаються кінцями і відкриваються в переддвер’я піхви (сечостатевий синус). У плацентарних ссавців нижні ділянки яйцепроводів зливаються в непарну піхву (vagina). Якщо кожна матка відкривається у піхву самостійним отвором, то її називають подвійною (багато мишоподібних, слони та ін.); у двороздільній матці права та ліва матки зрослися кінцями і відкриті у піхву загальним отвором (мишоподібні, деякі псоподібні, свині), а у дворогій - зрослися приблизно на половину своєї довжини (мідицеподібні, хижі, копитні, китоподібні). У частини лиликоподібних і приматів права та ліва матки злиті в єдину просту матку, в яку відкриваються права та ліва фаллопієві труби (рис. 5.66).
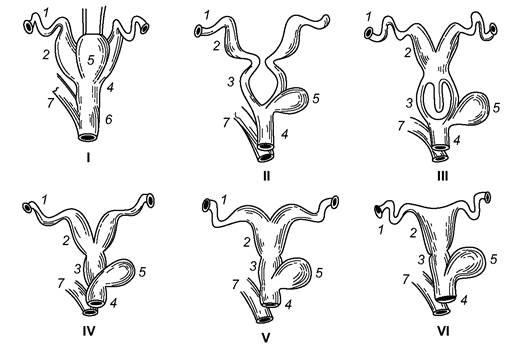
Рис. 5.66. Схема еволюції яйцепроводів у ссавців:
І - клоачного; ІІ - нижчого сумчастого (опосум); ІІІ - вищого сумчастого (кенгуру);
IV - VI - плацентарного ссавця з подвійною (ІV), дворогою (V), простою (VI) матками:
1 - фаллопієва труба; 2 - матка; 3 - піхва; 4 - сечостатевий синус; 5 - сечовий міхур; 6 - клоака; 7 - пряма кишка
Піхва переходить у короткий розширений сечостатевий синус (переддвер’я піхви), у який відкривається і протока сечового міхура. У стінці переддвер’я піхви розташований невеликий виріст із печеристими тілами - клітор, гомологічний копулятивному органу самців. Переддвер’я піхви відкриті назовні щілиноподібним сечовим отвором, обмеженим згортками шкіри - статевими губами. Розташовані тут шкірні залози виділяють пахучий секрет.
Яйцеві клітини - оогонії - утворюються в яєчниках із зачаткового епітелію; вони ростуть і перетворюються в ооцити; кожен ооцит оточують фолікулярні клітини, утворюючи первинний фолікул. До часу розмноження ооцит починає рости, накопичуючи жовток, а його оболонка розростається і між нею та яйцеклітиною - ооцитом - утворюється заповнений рідиною простір. Фолікул перетворюється у гра- афів міхурець, що видається на поверхні яєчника. Потім оболонка міхурця тріскає і яйцеклітина випадає в порожнину тіла (відбувається овуляція), звідки переходить у фаллопієву трубу, де й відбувається запліднення. Утворений на поверхні яєчника рубець заповнюють кров’яні згустки, він проростає лютеїновими клітинами, що містять жовтий пігмент і виконують гормональні функції. Так у яєчнику утворюється жовте тіло, що виділяє гормон прогестерон. У разі запліднення яйцеклітини і настання вагітності розвивається жовте тіло, а прогестерон стимулює імплантацію зародка в стінку матки, одночасно затримуючи утворення нових граафових міхурців і овуляцію до закінчення вагітності.
Оболонки зародка сумчастих ссавців у період розвитку лише прилягають до стінки матки, викликаючи частковий гістоліз її поверхневих шарів. У частини видів утворюється зачаток зародкової частини плаценти внаслідок зростання зовнішньої стінки алантоїса зі серозою, однак без утворення ворсинок, що проникають у товщу стінок матки. Живлення зародка відбувається завдяки жовтку яйця і секреторним виділенням клітин стінки матки. У плацентарних ссавців виникає справжня плацента: зрослі ділянки серози і зовнішньої стінки алантоїса утворюють хоріон (зародкову частину плаценти), вирости (ворсинки) якого занурені у стінку матки; у цьому місці матка розпушується й утворює материнську частину плаценти. Саме тоді посилюється гістоліз тканин матки, і кровоносні судини зародка тісніше контактують зі судинами материнського організму. Це полегшує живлення зародка речовинами крові матері (гемотрофне живлення). Утворення плаценти називають імплантацією зародка. Плацента може мати різну будову: її називають дифузною, коли ворсинки розміщені рівномірно по всьому хоріону, або часточковою, коли вони зібрані в окремі групи, чи дискоїдальною, коли ворсинки зосереджені в одному місці, утворюючи диск і т. д. Ступінь зв’язаності зародкової та материнської плацент неоднаковий, за цією ознакою розрізняють відпадну і невідпадну плаценти. У першому випадку зв’язок настільки глибокий, що під час пологів частина стінки матки відривається і виводиться назовні у вигляді посліду.
У одних видів дитинчата ссавців народжуються безпорадними, в інших - здатними до активних дій. Новонароджені сумчасті особливо дрібні. Вони недорозвинені, їх доношують у спеціальних сумках. Тому у сумчастих тривалість вагітності невелика: опосум - 12 днів, гігантський кенгуру - 38-40 днів. Чим більші розміри тварини, тим зазвичай триваліший період вагітності. Залежить він і від екології виду. Мідицеподібні народжують недорозвинених малят після 13-19 днів вагітності; вагітність триває 54-73 дні; у тварин, що народжують малят у норах із теплими гніздами, вагітність нетривала (хатня миша - 18 днів, сіра полівка - 16-23; ондатра - 25-26 днів); більша вона у великих за розмірами бабаків (30-40 днів). У звірів, які не мають добре влаштованих сховищ, вагітність триваліша: у нутрії - близько 130 днів, леопарда - 120, бурого ведмедя - 200 днів. Найтриваліша вагітність у тварин, малята яких відразу після народження слідують за самкою: у копитних - 150 (свині) - 290 (бики) днів; у білого носорога - 540 днів, у слонів - понад 600 днів; довготривала вона у тюленевих (морж - 200 днів) і китоподібних (270-365 днів).
Інколи вагітність продовжується латентним періодом, коли зародок на деякий час припиняє розвиток. Така діапауза дає змогу відкласти пологи до сприятливого періоду. Вона виявлена для так званих діестральних видів, тобто тих, що мають два періоди статевої активності. Наприклад, у борсука перша тічка проходить у липні- серпні, а друга - у жовтні. Запліднені в липні яйця перебувають у спокої до глибокої осені, а запліднені в жовтні розвиваються без діапаузи; дитинчата народжуються навесні. У горностая запліднені під час весняної тічки самки приводять малят через два місяці, а запліднені в серпні-вересні - тільки через 8-9 місяців. У козулі запліднені в першу тічку (липень-серпень) яйця сповільнено розвиваються до грудня, а запліднені під час осінньої тічки (листопад-грудень) затримки розвитку не мають. У куниці й соболя справжня тічка припадає на середину - кінець літа, а друга (несправжня) - на початок весни. Вона пов’язана з імплантацією діапаузованих яєць (пологи відбуваються навесні).
Швидкість постнатального (після народження) розвитку ссавців на перших стадіях пов’язана зі складом молока, яким мати вигодовує малят. Молоко ссавців має всі необхідні для розвитку речовини: білки, жири, вуглеводи, вітаміни, солі. Швидкість розвитку тим вища, чим більший у молоці вміст білків і жирів.
Розмноження ссавців ритмічне, тобто впорядковане у часі. Його можливості визначені тривалістю статевих циклів, які розпочинаються після статевого дозрівання. Терміни статевого дозрівання широко варіюють. Найплодовитіші дрібні тварини: полівки досягають статевої зрілості у віці 1,5-2 місяці, миші - 2-3 місяці, ондатра - 5 місяців, зайці - близько року. Більші за розміром звірі починають розмножуватися пізніше: вовки, куниці, лисиці й соболі - на другому році життя, тигри, ведмеді, багато видів тюленів і китоподібних - на 3-4-му році, олені - на 2-4-му році життя, носороги і слони - у 10-15 років. Статева зрілість у великих за розміром мавп настає у віці 10-12 років. Частота розмноження та його періодичність пов’язані зі смертністю і тривалістю життя. Висока плодючість - важливе пристосування дрібних ссавців, які фізіологічно нестійкі та мають численних і небезпечних ворогів. Вона компенсує їхню високу загибель. Багато мишоподібних потенційно здатні розмножуватися протягом цілого року, навіть у суворих умовах Арктики. Наприклад, зимове розмноження існує у норвезького лемінга. Багато разів його реєстрували також у сибірського лемінга. Зимове розмноження спостерігали у мишей і полівок, піщанок і хом’яків у лісовій, степовій та пустельній зонах. Щурі й миші хатні в людському помешканні також здатні розмножуватися протягом усього року. Велика амплітуда коливань плодючості і смертності дає змогу пояснити дуже мінливу чисельність названих вище тварин. Решта видів зазвичай розмножується в певний період року. Тривалість репродуктивного циклу залежить від тривалості вагітності. Пологи завжди припадають на сприятливий для вигодовування молоді період, переважно на весну. У період розмноження овуляція і тічка самок циклічно повторюються до запліднення. Такі статеві естральні цикли звичайно складаються з чотирьох фаз: передтічки (prooestrus), тічки (oestrus), післятічкового періоду (metaoestrus) і періоду спокою (disoestrus). Естральний цикл у дрібних мишоподібних триває близько 6 днів, у собак - 9-14, у копитних (кіз, овець, корів) - 17-24 дні.
Готовність до розмноження і приуроченість часу парування до певного періоду забезпечуються складним регуляторним механізмом, який діє за сигналами зовнішнього середовища. У помірних і високих широтах таким сигналом є зміна довжини світлового дня. Для видів із весняною тічкою сигнальне значення має збільшення довжини дня (багато тварин), для ратичних, які утворюють пари восени, - його скорочення, а для вовків - короткий зимовий день. Зміна світлового режиму впливає на нейрогіпофіз, який стимулює виділення передньою часткою гіпофіза гонадотропних гормонів, що, відповідно, впливають на гонади. У низьких широтах значення сигнального фактора можуть мати й інші періодичні явища природи, що передують наближенню сприятливого для розмноження періоду (в пустелях і саванах - опади, у тропічних лісах - дозрівання основних кормових рослин або їхніх плодів тощо). Навіть у тропічних лісах із їхнім постійним теплим і вологим кліматом розмноження ссавців зазвичай сезонне, хоча у різних видів припадає на певні періоди року.
Загальна плодючість ссавців, завдяки високому рівню і різноманітним формам турботи про потомство, невелика. У дрібних мишоподібних з високою смертністю виводок може сягати 12 малят (звичайно 5-6), а розмноження триває протягом усього року (до шести приплодів на рік). Зайці та вивірки дають 2-3 приплоди на рік, кожен із яких зазвичай становить від трьох до восьми малят (до 12). Вовки, лисиці, коти, соболі, куниці, горностаї розмножуються раз на рік і мають виводки з 3-6 малят. Раз на рік народжують 1-2 малят порожнисторогі, олені, тюлені й дельфіни. Слони, вусаті кити, тигри, деякі великі котові розмножуються раз на 2-3 роки, народжуючи зазвичай 1-2 малят. Плодючість змінюється залежно не лише від зовнішніх умов (забезпеченість їжею, погода), а й від щільності (чисельності) популяції: у разі її зростання збільшується частка ялових (не беруть участі в розмноженні) самок і зменшується розмір виводків.
Тривалість життя слонів становить 70-80 років, псових - 10-15, великих котів і китів - 30-40, вивіркових - 8-10, дрібних мишоподібних - 1-3 роки.
Псоподібні та мишоподібні в період розмноження утворюють пари або складні сім’ї (парцели мишоподібних, прайди левів). Такі групи з кількох самців, самок і молодих особин краще забезпечують виховання потомства. Вони часто зберігаються і після періоду розмноження. У стадних оленеподібних, тюленевих, китоподібних у період парування утворюються тимчасові «гареми», які складаються з кількох самок на чолі зі самцем-домінантом. Після парування самки з молодняком інколи утворюють окремі угруповання. Окремо тримаються поза періодом розмноження самці й самки кажанів.
Ендокринна система ссавців принципово подібна до гормональної системи решти хребетних тварин. Відмінності полягають не стільки у складі й кількості гормонів, скільки в посиленні їхньої активності та спеціалізації. Особливо це стосується гормонів, які беруть участь у здійсненні статевого циклу, вагітності й лактації (у самок - естрогени, пролактини, у самців - андрогени). Тісний взаємозв’язок гормональної діяльності ендокринних залоз між собою і центральною нервовою системою забезпечує узгоджений перебіг усіх життєвих процесів організму (в тому числі інтенсивності й характеру метаболізму, їхнього підлаштування до змінних умов зовнішнього середовища). Як і у птахів, розвиток вторинних статевих ознак у ссавців визначений не лише статевими відмінностями в характері генотипу, а й гормонами статевих залоз.
Нервова система. Головний мозок ссавців, зберігаючи спільні з іншими хребетними риси, відрізняється принциповими особливостями, що виділяють його в особливий кортикальний тип.
У головному мозку ссавців найбільшого розміру і складності досягає передній мозок. Більша частина мозкової речовини зосереджена в корі півкуль, тоді як смугасті тіла порівняно невеликі. Кора переднього мозку формується внаслідок розростання нервової речовини стінок бічних шлуночків. Утворене в такий спосіб мозкове склепіння називають вторинним, або неопаліум (neopallium); його зачатки з’являються
у амфібій, більше помітні у рептилій і птахів. Неопаліум складається з нервових клітин і безм’якотних волокон (сіра речовина мозку). Обидві півкулі з’єднані між собою комісурою з білих (мієлінових) волокон. Комісуру називають мозолистим тілом. Тіла нейронів у корі півкуль розташовані пошарово, утворюючи своєрідні екранні структури (рис. 5.67). Така організація мозку дає змогу просторово відображати зовнішній світ на підставі інформації, що надходить від органів чуття. Екранні структури характерні для найважливіших мозкових центрів ссавців, тоді як у інших хребетних вони трапляються зрідка, переважно в зорових центрах. Нова кора великих півкуль слугує центром вищої нервової діяльності, який координує роботу інших відділів мозку (рис. 5.68). Лобові частки відповідають за спілкування тварин, у тому числі акустичне; у людини вони пов’язані з мовою, тобто з другою сигнальною системою.

Рис. 5.67. Пошарове розташування нервових клітин у корі півкуль переднього мозку ссавців
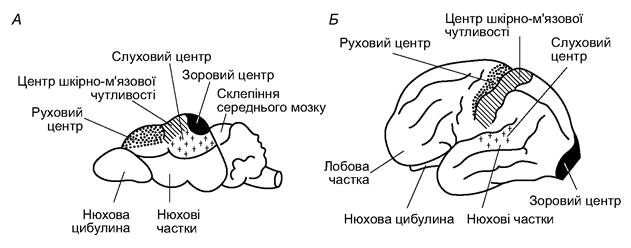
Рис. 5.68. Функціональні центри кори головного мозку землерийки (А) і людини (Б)
Кора майже всіх ссавців утворює більшу чи меншу кількість борозен, які збільшують її поверхню. У найпростіших випадках є тільки одна сильвієва борозна, яка відділяє лобову частку від скроневої; пізніше з’являється роландова борозна, яка розділяє лобову і потиличну частки, та ін. У приматів і зубатих китів кількість борозен особливо велика. Неопаліум ссавців більше, ніж комплекс кори середнього мозку птахів, забезпечує вищу нервову діяльність, нагромаджуючи сліди поодиноких збуджень і їхніх поєднань, збагачуючи так звану оперативну пам’ять. Це дає змогу на її підставі вибирати в новій ситуації оптимальне рішення. Найчастіше вони є новими комбінаціями вже відомих елементів. Виникнення у ссавців вищих асоціативних центрів - нової кори - не привело до ліквідації центрів, що керують інстинктивними актами, а лише підпорядкувало їх вищому контролю.
Інші частини переднього мозку мають порівняно менші розміри, однак також виконують важливі функції. Нюхові частки розташовані в передній частині знизу (нюхові цибулини і передні базальні ядра). Смугасті тіла виконують функцію регулювання інстинктивних реакцій під контролем кори півкуль.
Середній мозок малий; його дах розділений поперечними борознами і має назву чотиригорбкове тіло. Передні горби утворюють слабко виражену зорову кору, задні підконтрольні передньому мозку і слугують слуховими центрами (рис. 5.69). Мозочок великий і складається зі серединного черв’яка та прилеглих до нього півкуль із бічними придатками. Як і в інших ссавців, з мозочком пов’язані підтримка м’язового тонусу, пози, рівноваги, рухів частин тіла. Тісні зв’язки мозочка з корою півкуль свідчать про існування над ним вищого контролю.
Рис. 5.69. Схема зв'язків між відділами головного мозку ссавців:
1 - нюховий нерв; 2 - нюхова цибулина; 3, 4 - базальні ганглії; 5 - неопаліум; 6-9 - дорзальна (6) і вентральна (7) частини, медіальне
(8) і латеральне (9) колінчасте тіло зорового горба; 10 - зоровий нерв; 11, 12 - передні (11) (зорові) та задні (12) (слухові) горби даху середнього мозку; 13 - ретикулярна формація середнього мозку; 14 - руховий пірамідальний тракт; 15 - слуховий нерв; 16 - слуховий центр довгастого мозку; 17 - чутливі нерви шкіри і м'язів голови; 18 - чутливі ядра стовбура мозку; 19 - чутливі нерви шкіри і м'язів тіла; 20 - чутливі (дорзальні) роги спинного мозку; 21 - чутливі ядра довгастого мозку; 22 - ретикулярна формація довгастого і спинного мозку; 23 - рухові ядра стовбура мозку; 24 - рухові нерви до м'язів голови; 25 - рухові (вентральні) роги спинного мозку;
26 - рухові нерви (до м'язів тіла)
Довгастий мозок дає початок більшості головних нервів (V-XII). У ньому розташовані центри дихання, роботи серця, травлення тощо. З боків порожнини четвертого шлуночка відокремлюються пучки нервових волокон, які йдуть до мозочка й утворюють його задні ніжки. Нервові тракти зв’язують довгастий мозок зі спинним.
Як і в інших хребетних, відносні розміри головного мозку збільшуються зі зменшенням розмірів тіла і збільшенням напруження терморегуляції. Так, у великих мідицеподібних маса головного мозку становить близько 0,6% від маси тіла, а у дрібних - до 1,2, у великих китоподібних - близько 0,3%, а у дрібних - до 1,7%. Маса мозку приматів - 0,6-1,9% від маси тіла, у людини - близько 3%. У всіх ссавців маса переднього мозку перевищує масу інших відділів головного мозку: у різних групах вона становить 52-72% від загальної маси головного мозку; у приматів цей показник зростає до 76-80%, а в людини - до 86%.
Співвідношення мас головного і спинного мозку максимальне в людини (45:1), високе у приматів і китоподібних (10-15:1), нижче у псоподібних, мідицеподібних (3-5:1) і оленеподібних (2,5:1). У рептилій воно завжди менше 1. Спинний мозок за участю провідних шляхів (біла речовина) пов’язаний з рушійним центром кори півкуль, який виконує вищий контроль над руховими актами й управління складними рухами.
Ссавці мають 12 пар головних нервів; розвивається XI пара - додаткові нерви (nervus accessorius). Крім іннервації основних органів чуття (нюх, зір, слух) і м’язової системи, головні нерви беруть участь в утворенні вегетативної нервової системи, яка контролює так звані вегетативні процеси, що не підпорядковані вольовому контролю. Парасимпатична нервова система утворена черепними нервами довгастого мозку і спинномозковими нервами крижового відділу. Симпатична складається з нервових вузлів спинномозкових нервів шийного, грудного і поперекового відділів хребта (рис. 5.70). Основні системи органів мають закінчення обох систем.
Рис. 5.70. Схема вегетативної нервової системи ссавців:
1, 2 - парасимпатичні центри в середньому і довгастому мозку (1) та крижовому відділі спинного мозку (2); 3 - симпатичний стовбур; 4 - шийні вузли; 5 - зірчастий вузол; 6 - черевний вузол; 7, 8 - передній (7) і задній (8) брижові вузли; 9 - тазовий вузол; 10 - око;
11 - слинні залози; 12 - щитоподібна залоза; 13 - легеня; 14 - серце; 15 - печінка; 16 - шлунок; 17 - селезінка; 18 - підшлункова залоза; 19 - нирка; 20 - надниркова залоза; 21, 22 - тонка (21) та товста (22) кишка; 23 - сечовий міхур; 24 - матка; 25 - пряма кишка. Темні лінії - іннервація парасимпатичної системи, штрихові - іннервація симпатичної системи
Спинні стовбури білої речовини складаються з висхідних до головного мозку волокон, що несуть імпульси від органів чуття й ентерорецепторів (аферентна інформація). У черепних стовбурах переважають волокна, які несуть імпульси від мозку до м’язів та інших виконавчих органів (еферентна інформація). Контроль вищих центрів головного мозку над роботою спинного мозку досягає у ссавців найбільшого рівня.
Паралельну іннервацію пояснюють протилежно спрямованим впливом. Якщо імпульси однієї з них мають збуджувальний вплив на функцію органа, то імпульси іншого органа звичайно гальмують їх. Антагоністичний вплив, удосконалюючи регуляцію, значно розширює здатність витримувати пригнічувальні або дуже збудливі зовнішні впливи (стрес), збільшуючи шанси виживання організму в широкому діапазоні умов.
Органи чуття. Органи чуття по-різному розвинені в окремих рядах ссавців. Найважливішими для жителів відкритих просторів є зір, для нічних і присмеркових тварин (жителів лісових і чагарникових біотопів, норників і жителів водойм) - нюх і слух.
Нюх ссавців ефективніший, ніж у інших наземних хребетних. Велика роздільна здатність хеморецепторів дає змогу розрізняти окремі специфічні речовини (запахи) або їхнє поєднання, характерні для виду, групи особин або індивідів. У різних рядах і в окремих видів ссавців гострота нюху неоднакова. Сумчасті, мідицеподібні, мишоподібні, неповнозубі, більшість псоподібних і оленеподібних - так звані макросматики, відзначаються високорозвиненим нюхом; його використовують для орієнтації у просторі, пошуку їжі, у міжвидових і внутрішньовидових зв’язках. Більшість приматів і низка інших ссавців мають менш чутливий нюх (мікросматики).
Органи нюху розташовані у верхньо-задній частині носової порожнини, де виникає складна система раковин, вкрита слизовою оболонкою з нюхового епітелію з рецепторними клітинами та характерними волосками. Аксони цих клітин об’єднуються у групи, утворюючи волокна, що входять у нюхові цибулини, які через ланцюг нейронів з’єднані з центрами головного мозку. Складність будови нюхових раковин відповідає гостроті нюху (рис. 5.71).
Рис. 5.71. Поперечний розріз задньої частини носової порожнини ссавців:
А - мікросматик (людина); Б - макросматик (козуля)
У китоподібних наявність нюху та смаку заперечували, називаючи їх аносматиками. Недавні дослідження довели, що дельфіни мають пахучі залози, які відкриваються біля анального отвору; тварини здатні визначати за слідами секрету напрям табуна, що пройшов; вони сприймають запах крові як сигнал небезпеки. У ротовій порожнині вусатих китів є парні заглибини на кінці верхньої щелепи, гомологічні якобсоновому органу інших хребетних. В основі язика зубатих китів розташовані довгасті ямки, що нагадують смакові сосочки інших ссавців. Очевидно, за їхньою допомогою кити розпізнають запахи й орієнтуються, розрізняючи течії з різним хімізмом. Мозок китоподібних, хоч і відрізняється редукцією нюхових часток, однак зберігає в корі півкуль структури, пов’язані з аналізом хімічних сигналів.
Слух у житті ссавців відіграє важливу роль. Цьому відповідає і складне влаштування голосового апарату, здатного видавати різноманітні звуки, часто утворювати складні поєднання звуків, організовані в часі. За шириною звукового діапазону ссавці переважають птахів, широко використовуючи як ультразвуки (понад 20 кГц), так і низькі частоти. Слух і звукова сигналізація обслуговують найважливіші життєві явища - пошуки їжі, виявлення небезпеки, розпізнавання свого і чужого видів, різницю індивідів у групі (стаді або зграї), відносини батьків і малят та багато іншого. Особливості слуху відрізняються у різних рядах. Наприклад, для ехолокації кажани використовують переважно ультразвукові частоти в межах 40-80 кГц, проте видають і низькочастотні звуки до 12 Гц (не чутні для нашого вуха інфразвуки). Ще ширшим є звуковий діапазон вусатих китів - від кількох герців до двохсот кілогерців. Вусаті кити видають звуки низької частоти (1-2 кГц), значної сили і тривалості. Здатністю до ехолокації наділені мідицеподібні (землерийки) і деякі мишоподібні, що ведуть нірний спосіб життя. Різні діапазони використовує один вид для різних цілей: ехолокація та пошук здобичі - на високих і ультразвукових частотах, спілкування з особинами свого виду - на порівняно низьких.
Орган слуху ссавців складається з трьох відділів: зовнішнього, середнього і внутрішнього вуха. Зовнішнє вухо (вушна раковина) і зовнішній слуховий прохід - це своєрідна звукова антена, фільтр, здатний вбирати і посилювати біологічно важливі для виду звуки, послаблюючи сторонні шуми. Цю ж роль відіграє середнє вухо (рис. 5.72), у якому три слухові кісточки: молоточок, ковадло і стремінце - утворюють систему важелів, яка передає коливання більшій за розміром барабанній перетинці на меншу за площею мембрану овального віконця внутрішнього вуха, посилюючи ці коливання.
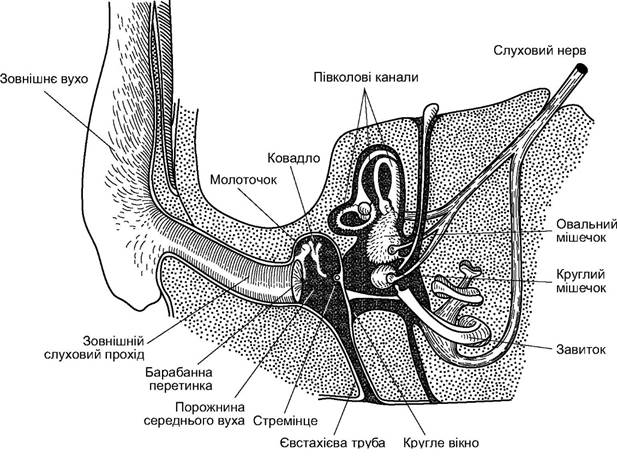
Рис. 5.72. Схема органа слуху ссавця
Кістковий барабан, у якому міститься порожнина середнього вуха, утворює ряд камер - резонаторів, що посилюють біологічно важливі звуки. У деяких видів, особливо норників, вони іноді заповнені губчастою кістковою масою, що гасить сторонні шуми. Середнє вухо євстахієвою трубою з’єднане з задньою частиною ротової порожнини, що забезпечує вирівнювання тиску повітря по обидва боки барабанної перетинки.
Внутрішнє вухо розташоване в товщі скроневої кістки (у її кам’янистій частині) і складається з вестибулярного та слухового відділів. Вестибулярний відділ охоплює три півколові канали й овальний мішечок; він є органом рівноваги і сприйняття просторового положення тіла. Слуховий відділ утворений круглим мішечком і пов’язаним із ним завитком, у якому розташований корті їв орган; функції цього органа полягають у первинному аналізі, переважно частотному, і кодуванні звукових сигналів, які після обробки передаються у слуховий центр (аналізатор) мозку. Завиток - спірально вигнута перетинчаста трубка, яка лежить у кістковому
футлярі, заповнена ендолімфою. У центрі завитка по всій його довжині йде базальна мембрана, на якій упоперек натягнуті фібрили (слухові струни). До них дотикаються чутливі клітини кортієвого органа, що сприймають коливання слухових струн, налаштованих на різну частоту. Імпульси, сприйняті чутливими клітинами, передаються нейронам, аксони яких утворюють слуховий нерв. Такий механізм забезпечує тонкий аналіз частотного спектра і часової організації звукового сигналу, що сприйнятий зовнішнім вухом і переданий посиленим через середнє до внутрішнього вуха.
Коливання голосових зв’язок верхньої гортані у ссавців приводить до утворення звуків. Ультразвукові сигнали кажанів генеруються апаратом рота або носа. У китоподібних в утворенні звуків беруть участь гортань у цілому, краї черпакуватих хрящів, повітряні мішки носового проходу і зовнішнє дихало. Окрім голосу, деякі ссавці використовують механічні звуки: клацання і скрегіт зубами (псоподібні, деякі мишоподібні, оленеподібні, примати), стукіт рогами, удари ногами об землю (багато норних тварин, копитні), шум від тертя голок (їжатець) тощо.
Зоровий аналізатор є третім основним органом чуття ссавців. Для деяких звірів, що ведуть переважно денний спосіб життя і живуть у відкритих біотопах, більша частина навколишньої інформації надходить через зоровий канал. Значення зору зменшується у жителів лісів, заростей і трав’яного покриву. У норників очі деколи перестають функціонувати, заростаючи шкірою (деякі кроти, сліпаки), або реєструють лише зміну освітлення (сліпушок, прометеєва полівка). У китоподібних очі задіяні лише для ближньої орієнтації. Очі ссавців розташовані або по боках голови, забезпечуючи майже кругове поле зору, за якого бінокулярний зір обмежений невеликим сектором, або фронтально. У другому випадку загальний огляд скорочується, однак поле бінокулярного зору збільшується. Перший тип переважає у копитних і мишоподібних, що постійно очікують нападу ворогів; другий характерний для мавп, які ведуть деревний спосіб життя і яким необхідно точно визначати відстань під час стрибків із гілки на гілку, а також для частини псоподібних, особливо котячих, які, нападаючи з засади, мають точно фіксувати відстань до жертви. Відносний розмір очей зростає у тварин із гострішим зором і у тварин із нічною активністю.
Око ссавців має зовнішню оболонку (склеру) з волокнистої тканини. У передній частині склера переходить у прозору рогівку. Під склерою лежить судинна оболонка з кровоносними судинами, що постачають необхідні речовини. Між склерою і судинною оболонкою у деяких тварин є шар клітин із кришталиками, що утворює дзеркальце (tapetum), яке відбиває промені та зумовлює «світіння» ока відбитим світлом (псоподібні, оленеподібні). Потовщуючись, судинна оболонка спереду переходить у райдужну і війчасте тіло (м’язи), за допомогою якого відбувається акомодація ока зміною форми кришталика. Райдужна оболонка відіграє роль діафрагми, регулюючи освітленість сітківки зміною розміру зіниці. Кришталик лінзоподібної форми порівняно малий у денних ссавців і різко збільшується у тварин із нічним способом життя. До внутрішнього боку судинної оболонки прилягає сітківка, що складається з зовнішнього пігментного і внутрішнього світлочутливого шарів. Колбочки не мають жирових крапель. Відмінності між видами зводяться до варіацій у співвідношенні паличок і колбочок, коливаннях загальної кількості рецепторних клітин і їхньої кількості на одне волокно зорового нерва. У норних тварин кількість рецепторних клітин і волокон нерва мінімальна: у сліпака в усій сітківці - 800 тис. рецепторів і 1 900 волокон у зоровому нерві (співвідношення 420:1). У нічних видів і жителів заростей воно вище: у їжака - 6,7 млн рецепторів на 8 400 волокон (760:1), у мишака жовтогрудого - 19,6 млн і 28 800 (680:1). Ще більше це число у жителів відкритих ландшафтів: наприклад, у зайця сірого - 192,6 млн рецепторів і 167 400 волокон (115:1), у макаки-резуса (примати) - 124,4 млн рецепторів на 1,2 млн волокон (105:1), а в кажана (лиликоподібні) - лише 8,9 млн рецепторів на 6 900 волокон (1130:1). Кількість рецепторних клітин, що в середньому припадають на одне нервове волокно зорового нерва, найменша у приматів; це дає змогу виявити більше деталей в об’єктах.
Багато ссавців здатні розрізняти кольори, проте, очевидно, слабше, ніж птахи. З цим пов’язане в середньому менш різноманітне забарвлення ссавців. Водночас ссавці розпізнають особливості форми предметів або їхніх частин, а також рухи, позу і міміку. Це забезпечено не ускладненням будови сітківки, а зоровим аналізатором у головному мозку, який у ссавців складніший, ніж у інших хребетних. Провідну роль відіграє зоровий центр кори півкуль переднього мозку, тоді як значення
зорової кори середнього мозку (передніх горбів чотиригорбкового тіла) зменшене. Перенесення основної обробки зорової інформації у центр півкуль переднього мозку відкрило можливості не тільки для візуальної орієнтації у просторі, а й для ускладнення і збагачення зорових зв’язків між особинами. Виникли у ссавців і набули широкого використання мова форми, поз, жестів і міміки. Це відіграє роль у врегулюванні відносин у популяціях і утворенні угруповань з узгодженою поведінкою (див. нижче). Забарвлення та форма тіла тварин під впливом природного добору набули маскувального значення (криптичне забарвлення) або були демонстрацією у разі погрожувальної поведінки.
На ділянках тіла, що часто стикаються з навколишніми предметами, ссавці мають особливо довгі й жорсткі волосини - вібриси. Їхні корені пов’язані з закінченням нервів. Особливого розвитку вібриси досягають звичайно на морді.