Зоологія хордових - підручник - Й. В. Царик - 2013
Розділ 4. ВІДДІЛ ЩЕЛЕПНОРОТІ, або ЕКТОБРАНХІАТИ, GNATHOSTOMATA, seu ECTОBRANCHIATA. НАДКЛАС Риби РІSCES
4.1. КЛАС ХРЯЩОВІ РИБИ CHONDRICHTYES
4.1.1. Особливості організації
Хрящові риби мають низку специфічних ознак:
✵ шкіра гола або вкрита плакоїдною лускою;
✵ скелет хрящовий, осьовий - із хрящових амфіцельних хребців (також може бути представлений хордою), розвинуті дуги хребців;
✵ череп амфістилічний, гіостилічний, а в суцільноголових - аутостилічний;
✵ хвостовий плавець гетероцеркальний;
✵ спільної зябрової покришки немає, зябра відкриваються назовні самостійними отворами;
✵ немає плавального міхура;
✵ у кишечнику розвинутий спіральний клапан;
✵ у серці між шлуночком і черевною аортою є артеріальний конус;
✵ запліднення лише внутрішнє, у самців є спеціальні вирости черевних плавців - птеригоподії;
✵ трапляються яйцекладні, яйцеживородні, живородні форми.
Зовнішній вигляд і покриви. Хрящові риби відрізняються від безщелепних наявністю рухомих парних плавців, які дають змогу вправніше плавати. Це одна із давніх груп морських хижаків. Особливості будови хрящових риб розглянемо на прикладі акул, у яких вони більше ніж за 250 млн років майже не змінилися.
Непарні плавці представлені спинними і хвостовим. Передній спинний плавець (pinna dorsalis anterior) розміщений посередині тулуба, задній (pinna dorsalis posterior) - у хвостовому відділі.
На передньому краї плавців частина видів має вільний колючий шип.
Хвостовий плавець (pinna caudalis) - гетероцеркальний. Парні плавці представлені грудними (pinnae pectorales) і черевними (pinnae ventrales).
Черевні плавці змикаються внутрішніми, або медіальними, краями. У самців, як уже зазначено, на задньому кінці кожного черевного плавця розвинутий особливий м’ясистий виріст - копулятивний орган - птеригоподій (pterygopodium).
Поступальний рух тіла вперед відбувається завдяки згинанню хвостового відділу вправо і вліво. Грудні й черевні плавці виконують функцію несучих площин і спрямовують тіло вгору, донизу та вбік.
На черевному боці тіла між задніми краями основ черевних плавців у хрящових риб розміщена поздовжня широка щілина - клоака. Поряд із заднім краєм клоаки є дві черевні пори, які з’єднані з черевною порожниною.
В акул спереду наявний р о с т р у м (rostrum), на вентральному боці якого є дві носові ямки, що утворюють вхідні й вихідні отвори нюхових капсул. Очі (oculus) частково прикриті шкірними складками - повіками (palpebra). На передньому й задньому краї очей виступає біла білкова оболонка - склера (sclera), решта ока вкрита прозорою рогівкою (cornea), з-під якої просвічує райдужка (iris).
За очима наявна щілина - бризкальце (spiraculum), яка з’єднує порожнину глотки із зовнішнім середовищем.
П’ять (у деяких - шість-сім) поперечних зябрових щілин розміщені спереду від грудних плавців (у скатів - нижче, на черевному боці). Кожна зяброва щілина прикрита шкірною складкою.
Рот півмісяцевий. Зуби різної форми, частіше - трикутні, розміщені в кілька рядів. На вентральній поверхні рострума, між ротом і першою зябровою щілиною, на дорзальному боці голови, над очима помітні рядочки отворів - пори, що ведуть до канальців системи бічної лінії.
Шкіра хрящових риб утворена багатошаровим епідермісом і коріумом. В епідермісі розміщені численні одноклітинні залози, слизистий секрет яких тонким шаром укриває тіло акули. Клітини нижніх шарів епідермісу містять пігменти. Спеціальні пігментні клітини є і в коріумі. Деякі скати здатні змінювати забарвлення залежно від кольору ґрунту, на який вони лягли. Цього досягають завдяки стисканню й переміщенню пігментних клітин углиб тіла, або, навпаки, розширенням усієї пігментної клітини.
У шкірі хрящових риб формується плакоїдна луска (рис. 4.1). Луска такого типу складається з розміщеної в коріумі округлої або ромбічної пластинки й ості (виступу), вершина якого через епідерміс виходить назовні.
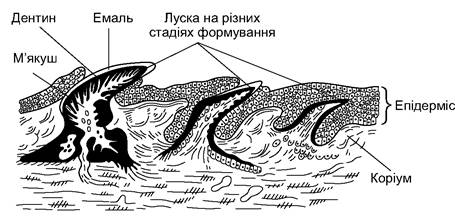
Рис. 4.1. Будова плакоїдної луски
На щелепах плакоїдні луски збільшені, набувають іншої форми й утворюють зуби. У цьому разі, перш за все, збільшується остя, зростає товщина емалі. Зношений або зламаний зуб відпадає, а на його місці виростає новий. Заміна зубів може відбуватися протягом усього життя риби. Зуби скатів переважно плоскі, притуплені - пристосовані до подрібнення захищених панциром або черепашкою тварин. У суцільно- голових окремі зуби зливаються в зубну пластинку. Зуби всіх хребетних тварин побудовані за плакоїдним типом, фактично це змінена плакоїдна луска предків. Змінені луски також утворюють плавцеві колючки у рогатих і колючих акул, голки у скатів- хвостоколів, пилкоподібні зубці в акул-пилоносів і риб-пилок. У частини скатів луски немає зовсім або є окремі збільшені луски, зубці яких перетворені на потужні шипи (морська лисиця).
Шкіра хрящових риб забезпечує їм достатньо надійний механічний захист, одночасно вона пропускає воду, а також розчинені в ній речовини.
Скелет і м’язова система. В сучасних хрящових риб у скелеті відсутні кістки. Внутрішній скелет повністю хрящовий, проте окремі ділянки можуть містити солі кальцію, що збільшує міцність елементів. Скелет поділяють на осьовий, скелет черепа (мозковий і вісцеральний), скелет парних і непарних плавців і скелет поясів парних плавців. Осьовий скелет пластинозябрових риб складається з хребетного стовпа (columna vertebralis), який поділяють на тулубовий і хвостовий відділи.
Хребці осьового скелета ззаду і спереду вгнуті (рис. 4.2). Такі хребці називають двовгнутими, або амфіцельними. У центрі тіла хребця є канал, у якому проходить хорда. Вона пронизує весь хребетний стовп, який має вигляд вервиці: розширений у місцях стикання хребців і звужений у тілах хребців. Над тілом хребців є верхні дуги. Між верхніми дугами сусідніх хребців розміщені вставні пластинки, які разом із дугами утворюють стінки хрящового каналу, в якому лежить спинний мозок.
У тулубовому відділі слабко розвинуті нижні дуги хребців утворюють поперечні відростки, до яких приєднані дуже короткі хрящові ребра. У хвостовому відділі розрослі нижні дуги правого й лівого боків з’єднані одна з одною, а між ними розташовані невеликі нижні вставні пластинки. Ця конструкція утворює гемальний канал, яким проходять хвостові артерія і вена.
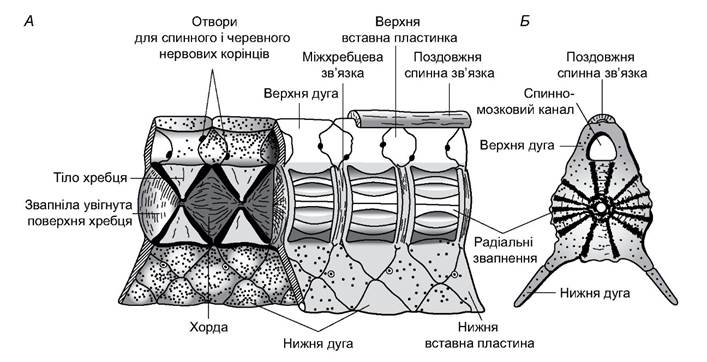
Рис. 4.2. Фрагмент хребетного стовпа акули (А) та поперечний розріз хребця (Б)
Під час онтогенезу в сполучнотканинній оболонці хорди, спочатку в кожному сегменті тіла, утворюються хрящові зачатки: два парні верхні та два парні нижні зачатки нижніх дуг. Пізніше навколо хорди починають розвиватися кільцеподібні хрящі - тіла хребців. У суцільноголових риб тіла хребців не утворюються. Сполучнотканинна оболонка хорди ущільнюється і в ній виникають кільцеві звапніння, які ззовні нагадують хребці. Передня пара хрящових зачатків розвивається у верхні дуги, які спираються на оболонку хорди, задня пара перетворюється на вставну пластинку. У хвостовому відділі розвиваються нижні дуги і нижні вставні пластинки. Ребер немає.
Череп хрящових риб розділений на два відділи: мозковий (осьовий) і вісцеральний (рис. 4.3). Мозковий череп (neurocranium) пластинозябрових риб - це суцільна хрящова коробка, яка з усіх боків оточує мозок: у склепінні черепа є затягнутий сполучною тканиною отвір - фонтанель. Передній кінець черепа витягнутий у рострум. В основі рострума з черепом зростаються парні нюхові капсули, а в задні бічні стінки вростають слухові капсули.
Потиличний відділ містить отвір, через який проходить спинний мозок. На бічних поверхнях черепа є заглибини - очниці, які захищають очі згори і з боків. Череп платибазальний: його основа широка, між стінками очниць міститься головний мозок.
Вісцеральний череп (splanchnocranium) сформувався внаслідок перетворення зябрових дуг, які у предків були опорою в ділянці глотки. У його побудові беруть участь щелепова дуга, під’язикова (гіоїдна) дуга і зяброві дуги. Щелепна дуга утворена двома парними хрящами. Лівий і правий хрящі спереду з’єднані (зрощені) один з одним. Функцію верхньої щелепи виконує піднебінно-квадратний хрящ (palatoquadratum), який із нижньою щелепою (меккелевим хрящем - cartilago Meckeli) утворює щелепний суглоб. На верхній і нижній щелепах розміщені зуби. У багатьох акул спереду щелепних дуг лежать 1-2 пари маленьких губних хрящів, які трактують як залишки двох редукованих зябрових дуг.
Під’язикова (гіоїдна) дуга розміщена одразу за щелепами. Вона складається з двох парних і одного непарного хрящів. Великий верхній парний хрящ - підвісок, або гіомандибуляре (hyomandibulare), - у пластинозябрових риб верхнім кінцем за допомогою суглоба і зв’язок рухомо прикріплений до слухового відділу мозкового черепа, а його нижній кінець рухомо з’єднаний як з обома елементами щелепної дуги в ділянці щелепного суглоба, так і з нижнім парним елементом під’язикової дуги - гіоїдом (hyoideum). Правий і лівий гіоїди з’єднані один з одним за допомогою невеликого непарного хряща - копули (copula). Такий тип з’єднання щелепної дуги з мозковим черепом - через верхній елемент під’язикової дуги - називають гіостилією.
Позаду під’язикової дуги розміщені п’ять пар (у плащоносних і багатозябрових акул - шість-сім) зябрових дуг, відділених одна від одної зябровими щілинами.
На відміну від пластинозябрових, у черепі суцільноголових роструму немає або він розвинутий слабко, нюхові капсули прирослі до мозкової коробки спереду. Зуби зрощені в зубну пластинку. Череп тропібазального типу (між очними ямками розміщена лише тонка міжочноямкова перегородка).
Скелет непарних плавців утворений різними за розміром рядами занурених у м’язи паличкоподібних хрящів - радіалій, які простягаються до основи плавця. Іноді частини радіалій зливаються у пластинки.
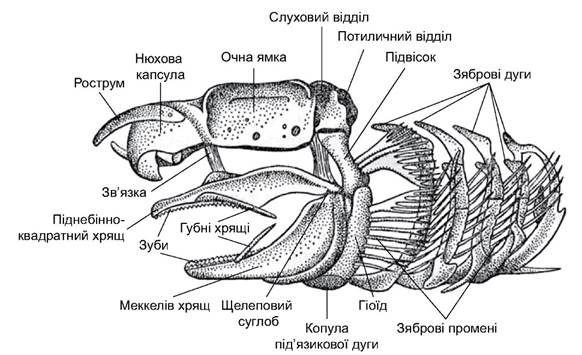
Рис. 4.3. Череп акули
Хвостовий плавець більшості хрящових риб гетеро- церкальний (рис. 4.4). Верхня лопать плавця значно більша від нижньої, до неї входить кінець хребетного стовпа.
Скелетну основу хвостового плавця утворюють видовжені верхні й нижні дуги хребців, а також ряд дорзоспінальних і вентроспінальних непочленованих хрящів. Шкірні лопаті плавця підтримувані еластотрихіями. У частини скатів і химер лопаті хвостового плавця зменшені або зовсім зникли.
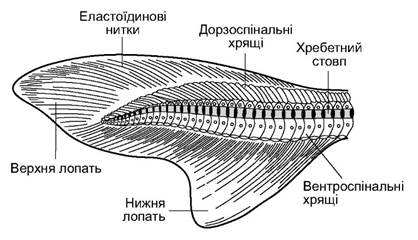
Рис. 4.4. Гетероцеркальний хвостовий плавець
Пояс передніх кінцівок (плечовий пояс) має вигляд хрящової дуги (рис. 4.5), яка лежить у товщі м’язової стінки позаду зябрового відділу. В акул і химер він ніяк не зв’язаний з осьовим скелетом, у скатів унаслідок збільшення розмірів грудних плавців верхні частини хрящової дуги пояса з’єднуються з хребтом. На бічній поверхні пояса з кожного боку є виріст - місце прикріплення скелета плавця, який має такі ж елементи, що і в непарних плавцях.
Тазовий пояс (або пояс черевних плавців) хрящових риб утворений хрящовою пластинкою (див. рис. 4.5), яка лежить у м’язах черевної стінки безпосередньо перед клоакою. До його бічних поверхонь прикріплений скелет черевного плавця.
Соматична м’язова система чітко сегментована і складається з м’язових сегментів - міомерів. Міомери відділені один від одного міосептами, які утворюють складну ламану лінію у вигляді повернутої боком латинської літери W. В інших частинах тіла м’язи можуть бути іншими. Наприклад, вісцеральна м’язова система - це шари гладких м’язів, які оточують травну трубку. У хрящових риб у ділянці щелепних і зябрових дуг гладкі м’язові волокна вісцеральної м’язової системи заміщені поперечно-посмугованими і формують достатньо складно диференційовані групи м’язів, які керують рухом щелепних і зябрових дуг.
Характерною особливістю м’язів хрящових риб є їхня відносна автономність, тобто здатність скорочуватися навіть після порушення зв’язку між м’язами й центральною нервовою системою. Зокрема, акула з відтятою головою може плавати, в іншому випадку - після видалення внутрішніх органів - нападати на здобич. Специфічним є хімічний склад м’язів хрящових риб. У них великий вміст сечовини - до 1,50-2,84% у морських і до 0,7% у прісноводних (у костистих риб - до 0,02-0,03%). Такий вміст сечовини тісно пов’язаний з регулюванням осмосу. У м’язах хрящових риб також високий вміст солей важких металів: Ті, Mn, Co, Ni. Значення цього феномена в житті акул не з’ясоване.
Загальна будова акули зображена на рис. 4.6.
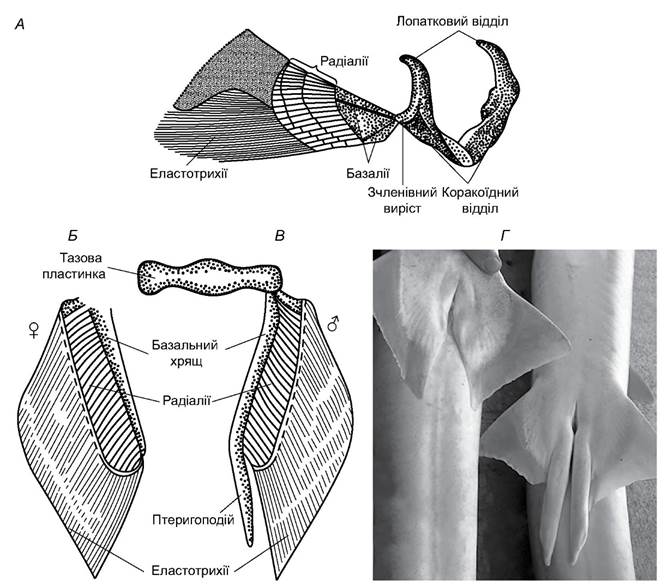
Рис. 4.5. Пояси кінцівок і скелети плавців акули: А - грудний плавець і плечовий пояс; Б, В - тазовий пояс і скелет черевного плавця самки і самця; Г - черевні плавці самки і самця
Травна система починається ротовим отвором (stoma) (рис. 4.6), який має форму щілини, обмеженої верхньою й нижньою щелепами. На дні ротоглоткової порожнини міститься язик (lingua). Це складка слизової оболонки, підтримувана непарним елементом - копулою під’язикової дуги. Такий язик не має власних м’язів, тому його рух пов’язаний із рухом під’язикового й зябрового апаратів.
Короткий м’язистий стравохід плавно переходить в об’ємний шлунок, який має V-подібну форму. За шлунком починається кишечник, у якому розрізняють три відділи: тонку кишку, в якій виділяють коротку дванадцятипалу (duodenum), куди впадають протока підшлункової залози і жовчна протока; товсту кишку, у якій є спіральний клапан із 12-13-ма обертами; пряму кишку з пальцеподібним виростом - ректальною залозою. Ректальна залоза - це орган сольового обміну. Пряма кишка впадає у клоаку. У черевній порожнині чимало місця займає трилопатева печінка, в якій багато жиру (у велетенської акули - до 70% від маси печінки).
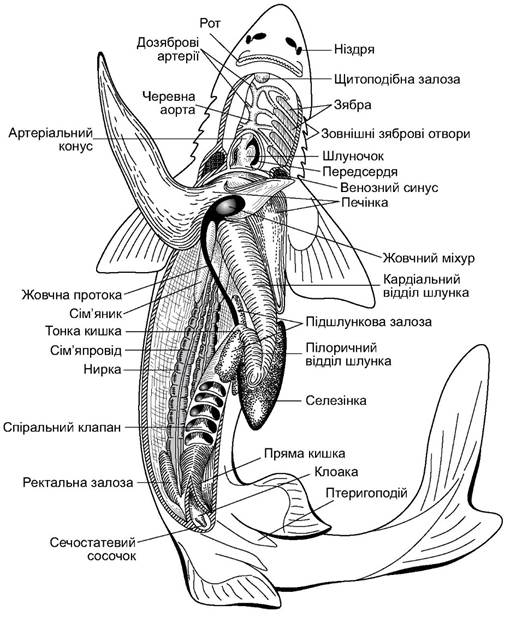
Рис. 4.6. Препарована акула (самець)
Жовчний міхур (vesica fellea) розміщений з правого боку центральної частки печінки. Його протока (ductus cysticus) з’єднана із протоками печінки (ductus hepaticus) і впадає у дванадцятипалу кишку.
Підшлункова залоза (pancreas) має дві лопаті: дорзальну і вентральну. Протока підшлункової залози також впадає у дванадцятипалу кишку.
Дихальна система хрящових риб представлена зябровими мішками, які відкриваються внутрішніми зябровими отворами в ротоглоткову порожнину, а зовнішніми - з боків голови. Зяброві мішки відділені один від одного зябровими перегородками (septum branchiale). Кожна зяброва перегородка починається на зябровій дузі, тягнеться до зовнішнього зябрового отвору і закінчується складкою, яка затуляє щілину (рис. 4.7). На зябрових перегородках є зяброві пелюстки. У багатьох хрящових риб наявне бризкальце - рудимент щілини між щелепною і під’язиковою дугами. В основі зябрових пелюсток є хрящові промені, які підтримують зяброву перегородку. Під зябровою дугою, в основі зябрової перегородки є три отвори зябрових судин: непарна широка судина - приносна артерія, якою венозна кров тече до зябер, і дві малі судини - виносні зяброві артерії, якими артеріальна кров відтікає від зябер.
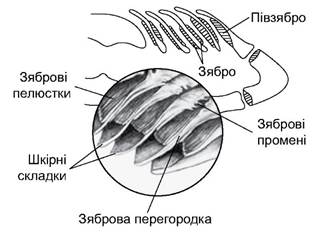
Рис. 4.7. Поперечний переріз зябер акули
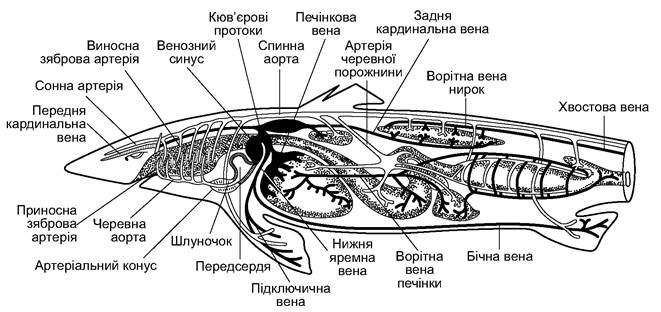
Рис. 4.8. Загальна топографія органів кровообігу акули
Серце (cor) хрящових риб, як зазначено, двокамерне, однак складається з чотирьох відділів. Задній відділ, куди впадають усі вени, називають венозним синусом. Він широким отвором з’єднаний з передсердям. Далі кров надходить у шлуночок - центральний відділ серця з товстими м’язовими стінками. Спереду шлуночка у вигляді товстої трубки лежить передній відділ серця - артеріальний конус, який переходить у черевну аорту. Серце розміщене в особливій навколосерцевій, або перикардіальній, порожнині (cavum pericardiale), яка є залишком целома. Ця порожнина відділена від черевної поперечною перегородкою і вистелена всередині тонким парієтальним листком очеревини - перикардієм (pericardium). Вісцеральний листок щільно облягає серце ззовні, його називають епікардієм (epicardium). Під ним
розміщений товстий шар м’язів, яка є основою стінки серця - міокард (myocardium). Внутрішня порожнина серця вкрита тонким листком, який називають ендокардієм (endocardium).
Від артеріального конуса відходить черевна аорта (aorta ventralis), яка розпадається на п’ять дозябрових артерій (arterii branchiales advehentes). Окислена в капілярах зябрових пелюсток артеріальна кров збирається у відзяброві артерії (arterii branchiales revehetes). Від першої відзябрової артерії відгалужується загальна сонна артерія (arteria carotis communis), яка забезпечує кров’ю голову.
Відзяброві артерії обох боків зливаються, утворюють розташовану під хребтом спинну аорту (aorta dorsalis). Від спинної аорти відходить низка артерій, які забезпечують кров’ю всі органи акули; далі спинна аорта заходить у гемальний канал хвостових хребців, її уже називають хвостовою артерією (arteria caudalis).
Венозна кров повертається до серця по венах - ширших, тонкостінних судинах. Хвостова вена (vena caudalis) збирає кров із хвоста, входить у порожнину тіла й розділяється на праву та ліву ворітні вени нирок (venae portae renalis), які підходять до нирок і розпадаються в них на капіляри. Капіляри зливаються, утворюють праву й ліву задні кардинальні вени (venae cardinalis posterior). З боків тіла йдуть парні бічні вени (venae lateralis), які збирають кров із черевних плавців і стінок тіла. У ділянці серця кожна з них зливається з підключичною веною (vena lateralis), яка несе кров від грудних плавців. Із голови венозна кров збирається у парні передні кардинальні вени (venae cardinalis anterior) і парні нижні яремні вени (venae jugularis inferior). Задні й передні кардинальні вени і нижні яремні вени кожного боку тіла зливаються в кюв’єрові протоки (ductus cuvieri), які впадають у венозну пазуху (венозний синус) серця.
Від шлунка, кишечника й селезінки венозна кров збирається в кілька вен, які перед печінкою зливаються у ворітну вену печінки (vena portae hepatica). У печінці ворітна вена розпадається на капіляри, у яких кров фільтрується й надходить до печінкової вени (vena hepatica), а нею - до венозного синуса.
Селезінка хрящових риб велика, виконує функцію кровотворного органа.
Видільна система хрящових риб - парні тулубові, або мезонефричні, нирки (рис. 4.9), які розміщені на спинному боці черевної порожнини і вкриті плівкою очеревини. Основну масу нирок становлять нефрони , утворені з мальпігієвих тілець (клубочків капілярних судин, що містяться в боуменових капсулах), від яких відходять ниркові канальці.
Ниркові канальці впадають або у вольфів канал, який виконує функцію сечоводу в самки, або у відділений від вольфового каналу сечовід - у самців. Вольфів канал у самців виконує функцію сім’япроводу. Продукти обміну надходять до нирок після фільтрації в мальпігієвих клубочках. Зазначимо, що для хрящових риб характерний високий вміст солей у тілі, що дає їм змогу зберігати осмотичну рівновагу зі середовищем (морська вода). Надлишок солей та сечовини виводиться назовні. Морські хрящові риби виділяють малу кількість сечі: від 2 до 50 мл на 1 кг маси тіла за добу. Для хрящових риб, які живуть в опріснених водоймах, це значення зростає до 250 мл на 1 кг маси за добу (риба-пила).
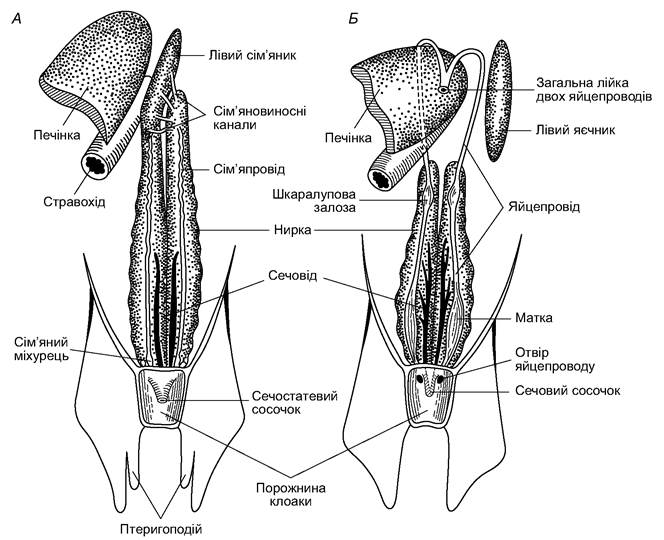
Рис. 4.9. Схема сечостатевої системи хрящових риб: А - самець; Б — самка
Статева система й особливості розмноження. У самців є парні сім’яники (testis), які звисають на брижах з боків стравоходу над печінкою (див. рис. 4.9). Протоки сім’яних канальців сім’яника впадають у ниркові канальці передньої частини нирки, яка має змінену будову і є придатком сім’яника. Протоки сім’яника (vas deferens) відкриваються у вольфів канал, який функціонує як сім’япровід. У задній його частині є сім’яний міхурець. Сім’япроводи правого й лівого боків відкриваються в порожнину сечостатевого сосочка, а той - у порожнину клоаки. Під час копуляції, внаслідок скорочення стінок сім’яного міхурця сперматозоїди потрапляють у клоаку самця, а звідти, за допомогою птеригоподіїв, - у клоаку самки.
Самки мають парні яєчники, підвішені на брижах у тих же місцях, що й сім’яники самців. У самок вольфів канал виконує функцію сечоводу. В них добре розвинуті мюллерові канали, які лежать на черевній поверхні відповідної нирки (правої, лівої).
Передні кінці мюллерових каналів виконують функцію яйцепроводів. У ділянці передньої частини нирки кожен яйцепровід має розширення - шкаралупову залозу; в задній частині яйцепроводу є помітне розширення - «матка». Правий і лівий яйцепроводи відкриваються у клоаку самостійними отворами з боків сечостатевого сосочка. Між яєчниками і яйцепроводами прямого зв’язку немає. Яйце, прориваючи яйцеву оболонку, потрапляє в порожнину тіла, а потім - у лійку яйцепроводу, де запліднюється. У ділянці шкаралупових залоз воно вкривається білковою оболонкою, а поверх неї - роговою. Остання часто має вирости. За допомогою виростів яйце може чіплятися до водоростей (рис. 4.10). Яйця великі, містять багато жовтка. Ембріональний розвиток зародка відбувається повільно. Для багатьох акул і скатів характерне яйцеживородіння: запліднене яйце затримується в матковій частині яйцепроводу до моменту виходу ембріона. Спеціального зв’язку між материнським організмом і ембріоном у цей час не виявлено.

Рис. 4.10. Яйця хрящових риб:
А - різнозубої акули; Б - котячої акули; В - пилоноса; Г- ската; Д — химери
Такий зв’язок виникає у кунячих акул (рід Mustelus) і акули-молота (рід Sphyma). У цих риб є своєрідна плацента (рис. 4.11).
Майже в усіх пластинозябрових народжена молодь підготовлена до самостійного життя.
Нервова система сучасних хрящових риб як хижаків складніша, порівняно з круглоротими і давніми костистими рибами. Вона представлена головним і спинним мозком та нервами, які відходять від них. На ділянці тулуба спинномозкові нерви утворюють складне плетиво, яке заходить у парні плавці.

Рис. 4.11.Зародок кунячої акули з жовтковою «плацентою»
Органи чуття диференційовані на систему шкірних органів бічної лінії, очі мають пристосовану для орієнтації у воді будову, статоакустичні органи - це три півколових канали.
Головний мозок містить передній мозок із покрівлею, яка утворена нервовою тканиною (рис. 4.12). Ззовні він виглядає розділеним на півкулі, однак усередині такого поділу ще немає. Проміжний мозок розділений на відділи й мозкову лійку. Середній мозок є головним центром зору, довгастий мозок - центром статоакустики й органів бічної лінії, а також вісцерального апарату (щелепого і зябрового). Мозочок відповідає за моторику всього тіла. Головні нерви в кількості десяти пар мають складне галуження.
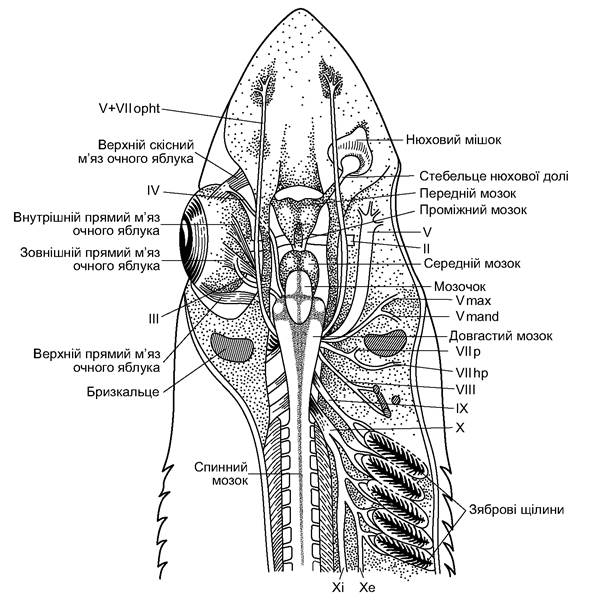
Рис. 4.12. Головний мозок і черепні нерви акули. Нерви: II-X - черепні нерви; II - зоровий; III - окоруховий; IV - бічний;
Vmax - верхньощелепна гілка трійчастого нерва; Vmand - нижньощелепна гілка трійчастого; V+VIIopht - очницеві гілки трійчастого і лицевого нервів; VIIp - піднебінна гілка лицевого нерва; VIIhm - гілка, що іннервує підвісок; VIII - слуховий; IX - язикоглотковий;
X - блукаючий; Xi - внутрішня гілка блукаючого нерва; Xe - бічна гілка блукаючого нерва
Симпатична нервова система хрящових риб, як і круглоротих, представлена ланцюжком гангліїв, між якими поки що немає прямого поздовжнього зв’язку (він з’явився лише у дводишних і костистих риб).
Як уже згадано, хрящові риби найбільше покладаються на такі системи чуттів, як нюхові органи, бічна лінія й органи зору (рис. 4.13).
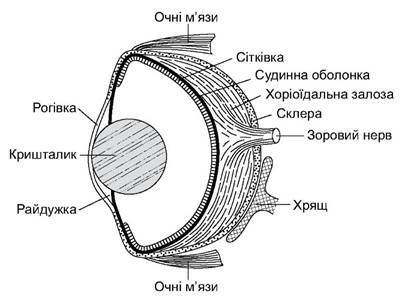
Рис. 4.13. Будова ока акули
Добре розвинуті органи нюху та бічна лінія. Завдяки бічній лінії тварини відчувають механічне переміщення води. На голові у деяких хрящових риб можуть бути скупчення ампул Лоренціні (занурена у шкіру капсула, яка відкривається назовні тонким каналом) (рис. 4.14).
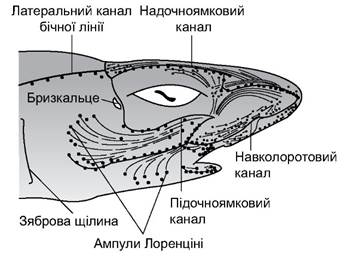
Рис. 4.14. Схема розташування органів бічної лінії
Завдяки цим ампулам риби відчувають зміну температури води (від 0,05°С) та, ймовірно, зміни параметрів електричних полів (від 0,10-0,01 мВ/см).
У деяких представників хрящових риб, зокрема, з ряду електричних скатів, є органи, які генерують електричні поля, а також електричні розряди. Ці електричні органи є структурно зміненими ділянками поперечно-посмугованих м’язів (рис. 4.15). М’язові волокна перетворюються на електричні пластинки, розділені драглистою сполучною тканиною.
Нижня поверхня пластинки накопичує негативні заряди, а верхня - позитивні. Скупчення пластинок у кількості 30-40 рядів утворюють призми, відділені одна від одної волокнистою сполучною тканиною. Кожен електричний орган складається з 350-600 призм. До нього підходять потужні гілки головних нервів (сьомої, дев’ятої та десятої пар). Електричний орган діє за принципом лейденської банки, накопичуючи електричний потенціал, який утворюється у процесі обміну в пластинках. Розряди у деяких скатів можуть сягати 60-300 В, а сила струму - 5 А. Упродовж кількох секунд скат може генерувати понад сотню таких розрядів.
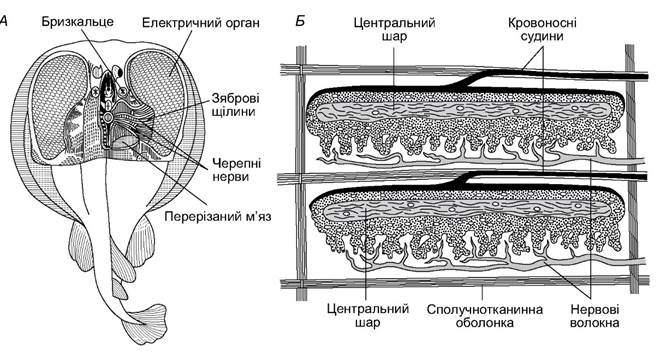
Рис. 4.15. Електричні органи ската Torpedo marmorata: А - препаровані електричні органи й головний мозок;
Б - будова електричного органа (дві електричні пластинки)