ЗООЛОГІЯ БЕЗХРЕБЕТНИХ - Г. Й. Щербак - 2008
ЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
ПІДЦАРСТВО СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA)
РОЗДІЛ ТРИШАРОВІ (TRIPLOBLASTICA), або БІЛАТЕРАЛЬНІ (BILATERIA) ТВАРИНИ
ПІДРОЗДІЛ ЛИНЯЮЧІ (ECDYSOZOA)
ТИП ЧЛЕНИСТОНОГІ (ARTHROPODA)
ПІДТИП ТРАХЕЙНОДИШНІ (TRACHEATA)
НАДКЛАС ШЕСТИНОГІ (HEXAPODA)
КЛАС КОМАХИ (INSECTA), АБО ВІДКРИТОЩЕЛЕПНІ (ECTOGNATHA)
Більшість видів комах - мешканці суші, вони населяють усі континенти, включаючи Антарктиду, зустрічаються в безводних пустелях, високогірних зонах вічних снігів, лісах і степах. Комахи опанували всі типи наземних біоценозів, а також ґрунти. Частина їх живе у прісних водоймах. Комахи - в основному вільноживучі тварини, але серед них є й паразити.
Кількість видів комах ще точно не встановлено: за різними підрахунками вже описано від 1 до 3 млн сучасних видів, і щорічно вчені відкривають ще кілька тисяч нових. Видовий склад комах України вивчено ще не достатньо: вважають, що їх не менш ніж 40 тис. видів.
Розміри комах коливаються від 0,25 мм до 26 см.
На відміну від класу Покритощелепні, в комах ротові придатки не занурені в ротову капсулу, звідси назва класу - Відкритощелепні. На голові є фасеткові очі та прості вічка. У більшості комах на другому та третьому сегментах грудей є крила. Рудименти черевних ніжок видозмінені у статеві придатки на кінці черевця.
Будова. Голова має вигляд суцільної капсули, на ній у дорослих комах розташовані пара складних (фасеткових) очей, одне-^гри простих вічка, пара вусиків (антен) і ротові придатки (рис. 358, 359).
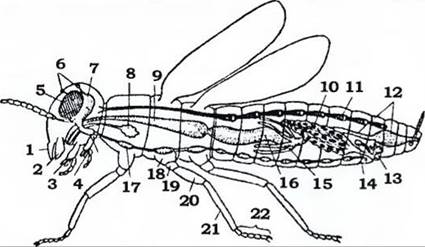
Рис. 358. Схема організації комахи (із Хадорна):
1 - верхня губа; 2 - мандибула; 3 - максила; 4 - нижня губа; 5 - фасеткове око; 6 - прості вічка;
7 - мозок; 8 - слинна залоза; 9 - воло; 10 - яєчник; 11 - серце; 12 - задня кишка; 13 - придаткові залози; 14 - сім'яприймач;
15 - мальпігієві судини; 16 - середня кишка; 17 - черевний нервовий ланцюжок; 18 - тазик; 19 - вертлюг; 20 - стегно;
21 - гомілка; 22 - лапка
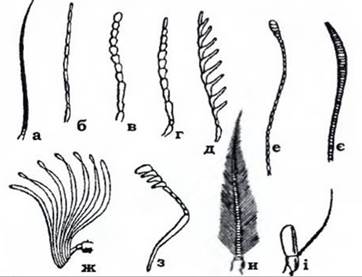
Рис. 359. Типи вусиків комах (з Догеля): а - щетинкоподібний; б - ниткоподібний; в - чоткоподібний; г - пильчастий; д - гребінчастий;
е - булавоподібний; є - веретеноподібний; ж - пластинчастий; з - колінчастий; и - пірчастий; і — щетинконосний
Ротові придатки складаються з непарної верхньої губи, пари нечленистих верхніх щелеп (ліандибул), пари нижніх щелеп (макігил) і непарної нижньої губи. Нижні щелепи та нижня губа двочленикові, мають по парі щупиків - органів дотику й смаку. До ротового апарату належить також м'язистий виріст ротової порожнини - гіпофаринкс. Залежно від характеру їжі розрізняють кілька типів ротових апаратів.
Гризучий ротовий апарат (рис. 360, а), найменш спеціалізований, пристосований для подрібнення твердої їжі. Верхня губа коротенька; мандибули мають жуйний край із зубцями на внутрішній поверхні. Максили складаються з основного членика, стовпчика, на якому є пара нижньощелепних щупиків, і двох жуйних лопатей - зовнішньої та внутрішньої.
Нижня губа має основний членик, або підборіддя, і дистальний, на якому є дві пари жуйних лопатей і пара нижньогубних щупиків.
У комах, які живляться рідинами, утворюється сисний хоботок. Так, у бджолиних ротовий апарат гризу- чо-лижучий (рис. 360, б); хоботок утворений нижніми щелепами й нижньою губою; верхні щелепи втратили жувальну функцію й беруть участь лише в побудові сот. У метеликів сисний ротовий апарат: зовнішні лопаті нижніх щелеп витягнулись у довгий хоботок, решта ротових частин редукована (рис. 361, а). У мух ротовий апарат лижучий·, м’який хоботок утворений нижньою губою, на кінці якої розвинений оригінальний фільтруючий апарат, що складається з великої кількості хітинових трубочок псевдотрахей.
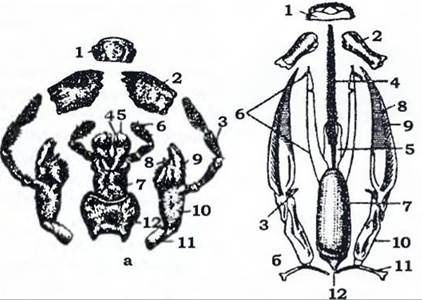
Рис. 360. Ротові апарати комах:
а - гризучий чорного таргана (з Аверенцсва); б - гризучо-лижучий джмеля (з Богданова-Катькова):
1 - верхня губа; 2 - верхня щелепа; З - нижньощелепний щупик; 4, 5 -внутрішня та зовнішня лопаті нижньої губи, відповідно;
6 - нижньогубний шупик; 7 - підборіддя; 8, 9 - внутрішня та зовнішня лопаті нижньої щелепи, відповідно; 10, 11 — стовпчик та основний членик нижньої щелепи, відповідно; 12- підпідборіддя
Комахи, які смокчуть рідини живих організмів, проколюючи їхні покриви, мають колючо-сисний ротовий апарат (рис. 361, б), в якому колюча частина хоботка складається з п’яти довгих голочок (мандибули, максили й гіпофаринкс), які вкладаються в жолоб видовженої нижньої губи, а зверху прикриті довгою верхньою губою, що зрослася в тоненьку трубочку. При укусі комар проколює шкіру жертви голками хоботка, а через трубочку верхньої губи, як по капіляру, кров надходить до рота. Такий ротовий апарат мають клопи, рівнокрилі, воші, блохи, довговусі двокрилі. У Гедзів щелепи й верхня губа мають вигляд ножів, які розрізають шкіру тварин: такий ротовий апарат зветься ріжучо-сисним.
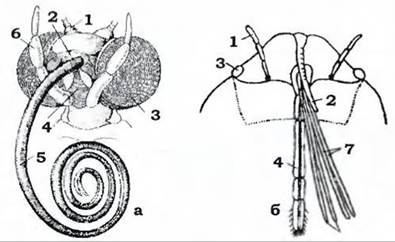
Рис. 361. Голови: а - метелика із сисним ротовим апаратом (з Догеля); б - клопа з колючо- сисним апаратом (із Шванвича):
1 - антена; 2 - верхня губа; 3 - фасеткове око; 4 - нижня губа; 5 - хоботок (нижні щелепи);
6 - нижньогубний щупик; 7 - верхні щелепи; 8 - нижні щелепи
У личинок комах з повним перетворенням типи ротового апарату часто відмінні від таких в імаго: наприклад, у гусені - гризучі, а в дорослих метеликів - лижучі. Імаго багатьох комах (одноденки, оводи, частина метеликів, у тому числі шовковичний шовкопряд та ін.) не живляться, і їхні ротові органи редуковані.
Груди складаються з передньо-, середньо- та задньогрудей. Кутикула кожного сегмента - це кільце, що поділяється на чотири склерити: спинний - тергіт, грудний - етерніт і два бічні - плейрити.
Кутикула грудей утворює внутрішні впинання, до яких прикріплюються м'язи ніг і крил.
Кожен грудний сегмент має пару ніг. Нога складається з п'яти члеників: базального членика - тазика, вертлюга, стегна, гомілки та
лапки з кількох члеників, останній членик закінчується одним чи двома кігтиками. Залежно від способу життя в комах розвинулися різні типи ніг: ходильні, бігальні, копальні, хапальні, стрибальні, плавальні тощо (рис. 362).
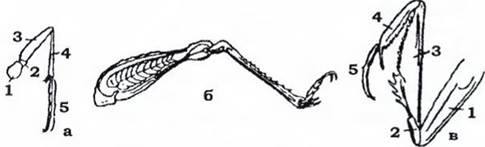
Рис. 362. Типи та будова ніг комах (з Аверенцева): а - бігальна туруна; б - стрибальна сарани; в - хапальна богомола: 1 - тазик; 2 - вертлюг; 3 - стегно; 4 - гомілка; 5 - лапка
Більшість комах мають крила. Це бічні складки покривів тіла, розташовані на середньо- та задньогрудях. Зазвичай їх дві пари: передні та задні. Крило складається з двох стінок - верхньої й нижньої. Кожна стінка утворена шаром гіподерми, зовні вкритої більш-менш розвиненою кутикулою (рис. 363). Між стінками є вузенька щілина (частина міксоцелі), заповнена гемолімфою. Крило має систему хітинових трубочок - жилок. Жилки виконують опорну функцію; у них також міститься гемолімфа, проходять трахеї й до клітин крила - нерви. Кількість і взаємне розташування жилок лежать в основі систематики комах.
У частини комах (прямокрилі, твердокрилі та ін.) передні крила перетворені на потовщені надкрила, які не беруть участі в польоті. Вони призначені для захисту ніжних задніх крил, складених під ними. У двокрилих задні крила перетворені на булавоподібні органи рівноваги - дзижчальця. У багатьох ґрунтових комах (робочі мурашки, терміти та ін.), а також у паразитів (воші, блохи) крила зникають, а в первин- нобезкрилих (Apterygoto) їх ніколи й не було.
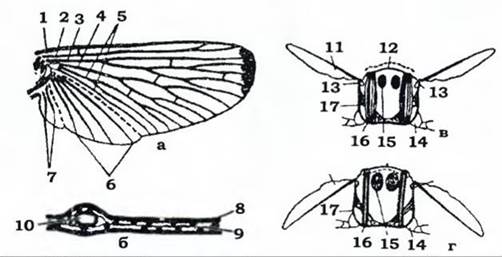
Рис. 363. Схема будови та роботи крил комахи (із Шванвинара - загальний вигляд крила; б - поперечний зріз; в - стадія піднятого крила; г- стадія опущеного крила: 1-7 - жилки: костальна, субкостальна, радіальна, медіальна, кубітальна, анальні, югальні, відповідно; 8 - кутикула; 9 - гіподерма; 10 - трахея; 11 - крило; 12 - тергіт; 13 - плейральний стовпчик; 14 - основа ноги; 15 - поздовжній м'яз; 16 - дорсовентральний м'яз; 17 — плейрит
Черевце в найбільш примітивних комах складається з 11 сегментів і тельсона, проте найчастіше їх буває вісім-дев'ять; у вищих груп (перетинчастокрилі, двокрилі) їхня кількість може зменшуватися до чотирьох- п'яти. На восьмому й дев'ятому сегментах розміщені зовнішні статеві придатки - геніталії: копулятивний орган у самців та яйцеклад у самок. Черевні кінцівки відсутні або видозмінені. Наприклад, у щетинкохвостих на всіх сегментах черевця є нечленисті придатки - гріїфельки; у щетинкохвостих, тарганів, одноденок на кінці черевця - членисті придатки - церки. Видозміненими кінцівками вважають також яйцеклад самок, який у жалячих перетинчастокрилих перетворюється на жало.
Покриви комах, як і всіх інших членистоногих, складаються з кутикули, гіподерми та базальної мембрани (рис. 287). Кутикула утворює зовнішній скелет і має поверхневий дуже тонкий шар - епікутикулу та внутрішній товстий - прокутикулу. Епікутикула дуже тонка й містить у своєму складі ліпоїди та воскоподібні речовини; во
на захищає комах від висихання. Прокутикула утворена двома шарами: екзокутикулою та ендокутикулою, що прилягає до гіподерми. Екзокутикула тверда, забарвлена пігментами, містить молекули хітину, пов'язані з білками, дубильні речовини, які роблять її твердою (склеротизують), а також пігменти. Ендокутикула м'яка, складається з молекул хітину, пов'язаного з білками, і має шарувату будову, що надає їй еластичності. Екзокутикула краще розвинена на тергітах і етернітах сегментів, у надкрилах жуків, мандибулах, члениках кінцівок; ендокутикула - в місцях зчленування кінцівок, крил, у міжсегментних мембранах.
Усю товщу прокутикули пронизують вертикальні порові канальці, всередині яких проходять тоненькі відростки гіподермальних клітин. Завдяки поровим канальцям здійснюються зв'язок гіподерми з кутикулою та утворення епікутикули під час линянь. Кутикула комахи, яка щойно перелиняла, м'яка й безбарвна. У результаті склеротизації вона твердне; паралельно із цим процесом відбувається меланізація
- синтез в екзокутикулі пігментів.
Гіподерма складається з одного шару призматичних клітин, між якими розмішені одноклітинні або багатоклітинні шкірні залози, а також спеціалізовані клітини, які утворюють волоски та сенсили. Зовнішня поверхня клітин гіподерми вкрита помітними лише під електронним мікроскопом мікроворсинками, що входять у порові канальні кутикули. Найважливішою функцією гіподерми є секреторна, бо вона виділяє речовини, з яких утворюється нова кутикула під час линянь. Від порожнини тіла гіподерму відділяє базальна мембрана, яка не має клітинної будови.
У комах розрізняють структурне та пігментне забарвлення. Перше - пов'язане з особливостями поверхневої структури кутикули й зумовлене оптичними ефектами інтерференції, дифракції та розсіювання світла. Це металічне, блискуче й переливчасте забарвлення деяких жуків і метеликів, особливо тропічних. Друге - зумовлено пігментами, які найчастіше містяться в екзокутикулі, рідше у клітинах гіподерми чи жирового тіла. У багатьох комах спостерігається поєднання структурного й пігментного забарвлення. Основні пігменти комах - меланіни (коричневі, чорні), каротиноїди, птерини (жовті, червоні), антрахінони (червоні).
Похідними покривів можуть бути: скульптурні (шипики, виступи, борозеньки, ямки тощо) та структурні (волоски, щетинки, лусочки крил метеликів тощо) придатки. Останні є або чутливими придатками, пов’язаними з нервовими клітинами, або термоізоляційними, що
створюють густий покрив, подібний до волосяного (у джмелів, деяких метеликів тощо).
Порожнина тіла - міксоцель - поділена двома поздовжніми горизонтальними перегородками - діафрагмами на три відділи (синуси): верхній, або перикардгальний, в якому розташована спинна кровоносна судина (рис. 364); нижній, або перинейральний, де міститься черевний нервовий ланцюжок; середній, вісцеральний, що лежить між діафрагмами; в ньому розташовані травна, видільна та статева системи, а також більша частина жирового тіла. Порожнина тіла заповнена гемолімфою.
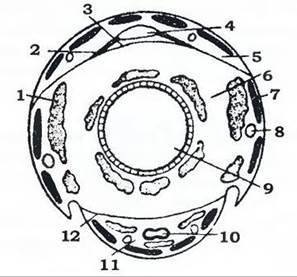
Рис. 364. Схема розташування серця та діафрагм на поперечному зрізі через тіло комахи (з Догеля, зі змінами):
1 - жирове тіло; 2 - крилоподібні м'язи; 3 - спинна діафрагма; 4 - серце; 5 - перикардіальний синус;
6 - перивісцеральний синус; 7 - поздовжній м'яз; 8 - трахея; 9 - кишечник; 10 - нервовий ланцюжок;
11 - перинейральний синус; 12 - черевна діафрагма
Мускулатура. М'язи комах є виключно поперечносмугастими. М'язова система диференційована, наприклад, у гусені метеликів налічується близько 2 тис. різних м'язів. Розрізняють скелетні м'язи, які рухають тіло та його придатки, і м'язи внутрішніх органів (вісцеральні). Кожен скелетний м’яз пркріплений до двох різних склеритів, і скорочення м'яза викликає зміщення склеритів один щодо одного.
Відносна сила м'язів комах дуже велика. Комахи можуть переносити вантаж, маса якого в 14-25 разів перевищує масу тіла. Стрибаючі комахи (сарана, блохи) за один стрибок долають відстань, яка в сотні або навіть тисячі разів перевищує довжину їхнього тіла. Більшість м’язів комах належать до синхронних: на один нервовий імпульс м’яз відповідає одним скороченням. Максимальна кількість скорочень таких м'язів не перевищує 30-40 за секунду. У двокрилих і перетинчастокрилих політ забезпечується асинхронними ("швидкими") м'язами. На кожен нервовий імпульс такий м'яз відповідає кількома (від 5 до 20) скороченнями, й кількість скорочень досягає 100 і більше, а в деяких дрібних двокрилих і перетинчастокрилих - навіть 1 тис. за секунду.
Травна система. Передня кишка складається з ротової порожнини, глотки, стравоходу, вола та м’язистого шлунка (рис. 365). У ротову порожнину відкриваються одна-дві пари слинних залоз. Перша пара залоз виділяє травні ферменти, а друга - може змінюватись на шовковіддільні (в гусені метеликів).
У передньому відділі кишечнику відбувається механічна обробка їжі та часткове її перетравлення ферментами слинних залоз. Залежно від типу живлення будова цього відділу варіює. Так, в імаго двокрилих і лускокрилих замість вола є сліпий мішкоподібний виріст стравоходу - харчовий резервуар, в якому деякий час зберігається рідка їжа, а у бджіл - "медовий шлунок" - сліпий виріст вола, де накопичується нектар, перетворюючись на мед. Жувальний шлунок м'язистий, вкритий всередині товстою кутикулою з гострими зубцями або товстими щетинками. Тут перетирається тверда їжа (прямокрилі, таргани, жуки) або фільтрується рідина (бджоли).
Середня кишка мас вигляд прямої трубки, інколи мішкоподібна, рідше має складну будову й поділяється на кілька відділів (у клопів). У багатьох комах середня кишка утворює пілоричні придатки - пальцеподібні вирости, призначені для збільшення всисної поверхні кишечнику й у деяких комах служать притулком для симбіотичних мікроорганізмів. У багатьох комах епітелієм середньої кишки безперервно виділяється тоненька плівка - перитрофічна мембрана, яка огортає їжу. Вона захищає стінку кишки від механічних ушкоджень твердими частинками їжі, а також полегшує проникнення травних ферментів до харчової грудки.
У багатьох комах спостерігається позакииисове травлення. Таргани змочують свою їжу слиною, а хижі комахи (туруни, жуки-плавунці) відригують у тіло здобичі травний сік середньої кишки та всмоктують уже перетравлену рідку їжу. Личинки мух, що живуть у трупах і гної, через анальний отвір виділяють травний сік із ферментами, які не лише перетравлюють субстрат, але й убивають і лізують гнильні бактерії та гриби.
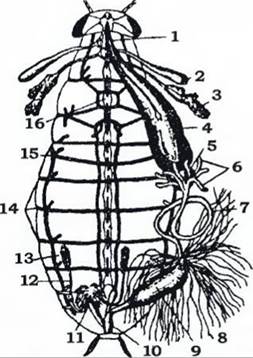
Рис. 365. Внутрішня будова самця чорного таргана при розтині зі спинного боку (з Аверенцева):
1 - стравохід; 2 - резервуар слинної залози; 3 - слинна залоза; 4 - воло; 5 - жувальний шлунок; 6 - пілоричні придатки середньої кишки;
7 - середня кишка; 8 - мальпігієві судини; 9 - товста кишка; 10 - пряма кишка; 11 - придаткові залози; 12 - сім'япровід; 13 - сім'яник;
14 - дихальця; 15 - трахея; 16 - черевний нервовий ланцюжок
Задня кишка складається з тонкої, товстої та прямої (ректум) кишок. У стінках прямої кишки є вирости - ректальні сосочки, які активно поглинають воду та мінеральні йони (Na+, К+, СІ), вилучаючи їх із вмісту задньої кишки й переводячи в гемолімфу, що забезпечує функцію осмореіуляції.
Типи живлення комах с надзвичайно різноманітними: серед них є фітофаги та зоофаги, в тому числі хижаки, паразити та кровососи, сапрофаги, некрофаги, копрофаги, є види, що живляться роговими утворами шкіри хребетних (шерсть, пір'я тощо), мертвою деревиною, воском тощо. Є також поліфаги. Способи живлення комах також є дуже різними: одні поїдають тверді речовини, другі поглинають рідини, треті є фільтраторами. Цим зумовлено різноманітність ротових апаратів. Кишковий тракт, хоч і модифікується залежно від способу живлення та складу їжі, але значно менше, ніж ротові органи.
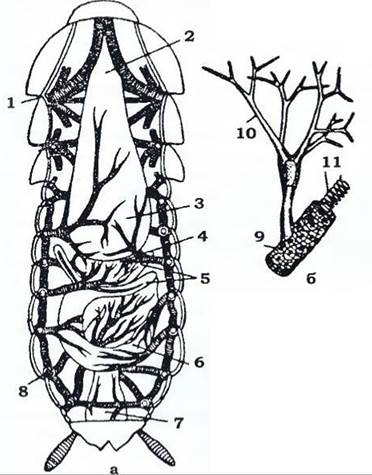
Рис. 366. Трахейна система чорного таргана (з Догеля):
а - загальний вигляд зі спинного боку; б - закінчення трахей з трахеолами
1 - грудні стигми ; 2- стравохід; 3 - воло; 4 - жувальний шлунок; 5 - пілоричні придатки;
6 - середня кишка; 7 - пряма кишка; 8 - черевні стигми; 9 - трахея; 10 - трахеоли; 11 - тенідій
У деяких комах травлення відбувається за допомогою симбіотичних мікроорганізмів. Наприклад, комахи, що живляться деревиною (терміти, деякі таргани, личинки пластинчастовусих жуків), не мають ферменту, який розщеплює клітковину. Його продукують симбіотичні найпростіші (джгутикові ряду Hypermastigida), бактерії та дріжджові гриби, що живуть у задній кишці.
Органи виділення. Виділення здійснюється мальпігієвими судинами, задньою кишкою, уратними клітинами жирового тіла, перикардіальними клітинами. Основними видільними органами є мальпігієві судини та задня кишка, які функціонують як єдине ціле. Кількість мальпігієвих судин коливається в різних комах від 2 до 200. Основний продукт виділення - нерозчинні кристали сечової кислоти. Зневоднені кристали сечової кислоти разом із фекаліями виводяться назовні через анальний отвір. Уратні клітини жирового тіла, перикардіальні та гіподермальні клітини вилучають з гемолімфи продукти обміну й нагромаджують їх без виведення назовні.
Дихання відбувається через трахейну систему (рис. 366). Зсередини трахеї вистелені тоненькою кутикулою, яка має спіральні потовщення - тенідії, що запобігає спаданню трахей. Від дихалець відходять короткі поперечні трахеї, з'єднані між собою трьома парами поздовжніх трахейних стовбурів, від яких ідуть розгалуження до всіх органів і закінчуються тоненькими трубочками діаметром 1-2 мкм - трахеолами. Кінці трахеол лежать на поверхні окремих клітин або входять усередину них. Кисень з трахеол безпосередньо дифундує у клітини, а вуглекислий газ - з тканин у трахеоли. У комах, що добре літають, на поздовжніх трахейних стовбурах утворюються розширення - повітряні мішки. Вони не мають тенідіїв і можуть змінювати об'єм. Повітряні мішки беруть участь у вентиляції крилової мускулатури під час польоту й виконують аеростатичну функцію через зменшення питомої маси тіла.
У багатьох водних й ендопаразитичних комах трахейна система замкнена, без дихалець, і вони дихають через покриви киснем, розчиненим у воді чи в рідинах тіла хазяїна. У водних личинок можуть розвиватися трахейні зябра - тонкостінні вирости шкіри, в які продовжуються трахеї, утворюючи там густу сітку (рис. 367). Розчинений у воді кисень через зяброву поверхню переходить у трахейну систему в газоподібному стані. У личинок одноденок, жуків-вертячок, волохокрильців тощо такі зябра метамерно розташовані на черевці; у німф різнокрилих бабок трахейні зябра розмішені всередині задньої кишки.
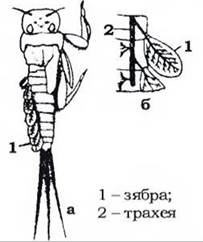
Рис. 367. Трахейні зябра личинки одноденки (а) та схема їхньої трахеації (б) (з Кузнецова)
На відміну від інших членистоногих у комах транспорт кисню до тканин відбувається безпосередньо за допомогою трахей, без участі кровоносної системи.
Кровоносна система дуже редукована через майже повну втрату функції транспорту газів. Від неї залишається спинна судина, розташована в перикардіальному синусі (рис. 364). Задня її частина - серце складається з кількох послідовних камер, кожна з яких має пару бічних отворів - остій з клапанами. Через них гемолімфа потрапляє з перикардія всередину серця. Камери сполучені між собою отворами, в частини комах - з клапанами, що не дозволяють крові рухатись назад. Задній кінець серця замкнений, передній продовжений у трубчасту
аорту, яка відкривається в міксоцель поблизу голови. До верхньої діафрагми та нижнього боку кожної камери прикріплена пара крилоподібних м’язів. Пульсацію серця зумовлено еластичністю його стінок, а також роботою крилоподібних та інших м'язів. Частота серцевих скорочень у різних комах коливається від 10 до 150 за хвилину. Біля основи антен, ніг, крил є додаткові пульсуючі органи, які нагнітають у них гемолімфу. Найчастіше це м'язові ампули та скоротливі перетинки.
Гемолімфа комах складається з рідкої плазми й клітин - гемоцитів, які або плавають у плазмі, або нерухомо осідають на поверхні внутрішніх органів. Серед них є фагоцити та клітини, які нагромаджують поживні речовини й транспортують їх у тканини. Гемолімфа виконує низку важливих функцій: транспорт поживних речовин, гормонів та інших біологічно активних речовин, а також продуктів обміну до відповідних органів, тканин і клітин; захист організму від інфекційних та інвазійних захворювань; підтримка сталості хіміко-фізичних властивостей внутрішнього середовища організму. Важливою є й механічна функція - внаслідок гідростатичного тиску гемолімфи змінюється форма органів з м'якою кутикулою: розправляються крила в імаго після виходу з лялечки, розкручується хоботок метеликів тощо. Гемолімфа багатьох комах отруйна й служить для захисту від ворогів. За небезпеки вона виділяється через суглоби кінцівок і вусиків назовні. Це явище відомо в жуків-сонечок, листоїдів. Жук шпанська мушка має в гемолімфі отруйну речовину - кантаридин, яка викликає запалення шкіри й загальну інтоксикацію у хребетних.
Жирове тіло разом з гемолімфою утворює внутрішнє середовище організму. Це пухка тканина мезодермального походження, яка складається із численних лопатей між внутрішніми органами (рис. 368, б). Переважну більшість клітин жирового тіла становлять трофоцити. У них нагромаджуються резервні поживні речовини - жири, білки й глікоген. Крім того, як уже згадувалось, у ньому є уратні клітини, в яких нагромаджуються кристали сечової кислоти.
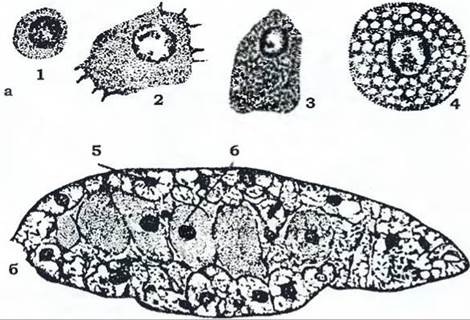
Рис. 368. Гемоцити (а) та жирове тіло (б) чорного таргана:
1 - недиференційована клітина; 2 - фагоцитуюча клітина;
З, 4 - клітини з білково-глікогенними та жировими резервами; 5 - жирові клітини; 6 - клітини із симбіотичними бактеріями
Основною функцією жирового тіла є накопичення резервів поживних речовин на личинковій фазі й забезпечення ними організму на час метаморфозу, діапаузи, голодування, дозрівання статевих продуктів. При цьому жирове тіло не лише пасивно нагромаджує поживні речовини, але й здійснює проміжний обмін речовин. У його клітинах відбуваються процеси біосинтезу й перетворення білків, жирів, вуглеводів. У самок під час розмноження жирове тіло синтезує специфічні білки - вітелогеніни, які необхідні для утворення жовтка в яйцях, що розвиваються. У багатьох комах у жировому тілі є особливі клітини - міцетоцити, у цитоплазмі яких живуть симбіотичні мікроорганізми - бактерії та гриби, які продукують деякі вітаміни та інші біологічно активні речовини, необхідні комахам.
Деякі комахи, наприклад жуки-світляки (родина Lampyridae), мають органи свічення - видозмінені ділянки жирового тіла. У них міститься речовина - люциферин, яка під впливом нервового імпульсу та дії ферменту люциферази здатна світитись. У дорослих комах взаємні світлові сигнали певної частоти призначені для зустрічі особин різної статі.
Нервова система. У головній капсулі розміщуються надглотковий ганглій, навкологлоткові конективи й підглотковий ганглій. Надглотковий ганглій, або мозок, складається з трьох злитих разом гангліїв: протоцеребрума, дейтоцеребрума й тритоцеребрума (рис. 369). Про тоцеребрум розвинений краще, ніж інші, й має найскладнішу будову. У ньому розрізняють кілька гангліозних центрів, серед яких найбільш розвиненою є пара стебельчастих, або грибоподібних тіл, які є вищим асоціативним і координуючим центром нервової системи. Вони досягають найбільшого розвитку в комах зі складними формами поведінки, особливо в перетинчастокрилих. Крім того, у протоцеребрумі розташована пара великих зорових часток, які іннервують складні очі. Дейтоцеребрум містить парні шошні центри, він іннервує антени. Тритоцеребрум іннервує верхню губу, з ним пов'язана також вегетативна (симпатична) нервова система.
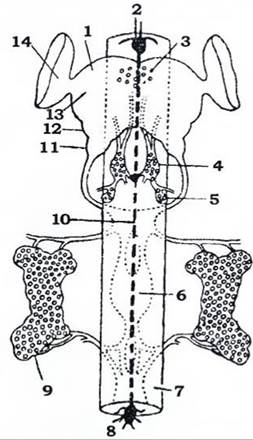
Рис. 369. Схема ендокринної та вегетативної (стоматогастричної) систем комах (з Проссера й Брауна):
1 - надглотковий ганглій; 2 - фронтальний ганглій; 3 - нейросекреторні клітини; 4 - кардіальні тіла; 5 - прилеглі тіла;
6 - ганглії черевного нервового ланцюжка;7 - кишечник; 8 - шлунковий ганглій; 9 - проторакальні залози; 10 - поворотний нерв;
11 - тритоцеребрум; 12 - дейтоцеребрум; 13 - протоцеребрум; 14 - зорова частка
Підглотковий ганглій складається з трьох парних гангліїв, які іннервують ротові органи й слинні залози. Черевний нервовий ланцюжок у прямокрилих, тарганів складається з трьох грудних і восьми черевних гангліїв; у вищих груп відбувається злиття всіх гангліїв у два-три або навіть один великий ганглій, наприклад, у вищих мух і жуків.
Крім центральної, в комах добре розвинена вегетативна нервова система, яка іннервує внутрішні органи (рис.369).
З нервовою системою тісно пов'язана ендокринна система, до якої належать парні кардіальні та прилеглі тіла, які розташовані в голові позаду мозку, нейросекреторні клітини та проторакальні (передньогрудні) залози. Ендокринні органи синтезують, нагромаджують і виділяють у гемолімфу гормони, які регулюють усі фізіологічні процеси в організмі.
Органи чуття дуже різноманітні, що пов’язано із загальним високим рівнем організації та складною поведінкою комах, яка вимагає точної інформації про навколишній світ.
Морфологічну й функціональну основу чутливості комах становлять нервово-чутливі одиниці - сенсили. Вони або розкидані по різних частинах тіла, або зібрані у скупчення - органи чуття.
До механорецепторів належать органи дотику, слуху й сейсмічного чуття. Дотикові сенсили (рис. 288) розкидані по всьому тілу, найбільше їх на антенах, ногах, яйцекладі тощо. Органи слуху й сейсмічного чуття сприймають звуки й коливання середовища. Сенсили цих органів звуться сколопідіями. їхню основу становить нервова клітина, чутливий відросток якої (довгий нерухомий джгутик) оточений по всій довжині кутикулярним чохлом - сколопсом. Дистальний кінець джгутика входить у канал шапочки, оточеної шапочковою клітиною (рис. 370). Натискання шапочки на дистальний кінець джгутика збуджує чутливу клітину.
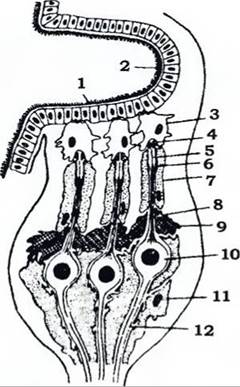
Рис. 370. Група із трьох сколопідіїв у тимпанальному органі сарани
(з Тищенка): 1 - тонка й жорстка ділянки тимпанальної мембрани;
2 - товста й м'яка ділянки тимпанальної мембрани; 3 - шапочкова клітина; 4 - шапочка; 5 - нерухомий джгутик; 6 - сколопс;
7 - облямовуюча клітина; 8 - дендрит чутливої клітини; 9 - фіброзна клітина; 10 - чутлива клітина; 11 - гліальна клітина;
12 - аксон чутливої клітини
Сколопідії входять до складу хордо- тональних і тимпанальних органів.
Хордотональні органи - це сукупність сколопідіїв, натягнутих між двома ділянками кутикули.
Вони розташовані на різних частинах тіла - у ротовому апараті, антенах, грудях, ногах, крилах, черевці та його придатках і сигналізують нервовій системі про рухи тіла та його придатків. Деякі хордотональні органи чутливі до вібрацій і сильних звуків.
Тимпанальні органи є спеціалізованими органами слуху. Вони подібні до хордотональних, але відрізняються від останніх тим, що сколопідії в них прикріплюються до витонченої у вигляді барабанної перетинки ділянки кутикули й сприймають її коливання під дією звукових хвиль. Слух розвинений не в усіх комах. Найчастіше слухові органи мають ті з них, які самі здатні створювати звуки. У саранових вони лежать по боках першого сегмента черевця, У коників і цвіркунів - на гомілках передніх ніг, у співочих цикад - в основі черевця. Найскладнішу будову мають тимпанальні органи коників (рис. 371).
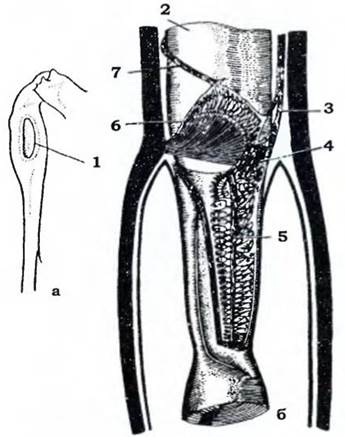
Рис. 371. Тимпанальний орган коника (з Догеля): а - загальний вигляд; б - поздовжній зріз:
1 - отвір тимпанального органа; 2 - головна трахея; 3 - слуховий нерв;
4 — проміжний орган; 5 - слуховий гребінь; 6 - підколінний орган; 7 - нерв
Терморецептори й гігрорецепторн здатні сприймати зміни температури та вологості. Вони розміщені на різних частинах тіла, в основному на антенах. їхні кутикулярні частини мають вигляд ямок або горбків. Одна й та сама сенсила може бути і термо-, і гігрорецептором, якщо вона має кілька чутливих клітин.
Хеморецепторні сенсили нюшні та смакові - зустрічаються на різних придатках тіла: антенах, ротових кінцівках, лапках ніг, церках, яйцекладі. Для кутикулярних частин хеморецепторних сенсил характерна наявність однієї або кількох пор, через які молекули хімічних речовин проникають до відростків чутливих клітин. Інша особливість - це наявність у сенсилі не однієї, а кількох чутливих клітин, кожна з яких реагує на подразнення певною групою речовин. Так, до складу рецепторів смаку, як правило, входять три або чотири чутливі клітини. Одна з них реагує на розчини солей, друга - збуджується розчинами цукрів, третя - чистою водою. До складу деяких дистантних хеморецепторів входять більше 10 чутливих клітин.
Комахи мають надзвичайно розвинений нюх, особливо на специфічні речовини, які їх приваблюють (атрактанти) або викликають відразу (репеленти). Серед атрактантів особливо важливе значення в житті комах мають харчові й статеві атрактанти. Перші полегшують їм пошук їжі, інші, які виділяються самками, дозволяють самцям знаходити їх на великій відстані. Відомо, наприклад, що незапліднені самки деяких метеликів здатні приваблювати самців на відстані 3-9 км.
Органи зору комах представлені трьома типами очей: складними, або фасетковими, очима, латеральними та дорсальними вічками. Фасеткове око складається з великої кількості фоторецепторів - оматидіїв, а кожне латеральне та дорсальне вічко відповідає окремому фоторецептору.
Фасеткові очі мають майже всі дорослі комахи й личинки комах з неповним перетворенням. Вони розташовані по боках голови й тісно пов'язані з добре розвиненими зоровими частками мозку. Око складається з окремих зорових сенсил - оматидіїв, кількість яких може досягати кількох сотень і навіть тисяч. Оматидій має вигляд сильно витягнутого конуса, зверненого основою до поверхні ока, і є сукупністю світлозаломлюючого, світлочутливого та світлоізолюючого елементів (рис. 372).
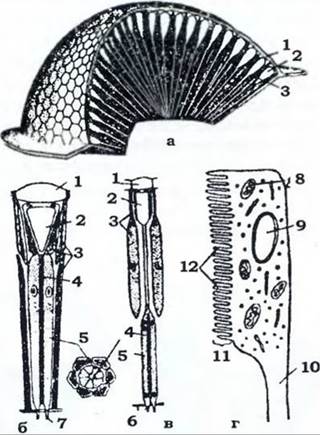
Рис. 372. Схема будови фасеткового ока комахи (а), оматидій денної (б) та нічної комах (в), окрема світлочутлива клітина (г)
(а-в - з Догеля, г - з Тищенка):1 - кришталик, що входить до складу рогівки; 2 - кристалічний конус;
З - пігментні клітини; 4 - рабдом; 5 — чутливі клітини; 6 - базальна мембрана; 7 аксони чутливих клітин; 8 - мітохондрія;
9 - ядро; 10 - аксон; 11 - рабдомер; 12 - мікроворсинки
Світлозаломлювальний апарат оматидія складається з кришталика й кристалічного конуса, які в сукупності виконують роль лінзи.
Світлочутливий апарат складається з кількох (найчастіше восьми- дев'яти) чутливих зорових (ретинальних) клітин з нервовими відростками, які зв'язують їх з мозком. Вони розташовані по колу, як частки апельсина. Кожна зорова клітина має по всій довжині внутрішньої поверхні особливу структуру складної мікроскопічної будови - рабдомер, в якій є зорові пігменти. Рабдомери всіх зорових клітин щільно прилягають один до одного й утворюють зорову паличку, або рабдом. Саме в рабдомах здійснюється фоторецепція - перетворення світлового сигналу на нервовий імпульс. Світлоізолюючий апарат складається з кількох пігментних клітин, які оточують кристалічний конус і ретинальні клітини оматидія. Вони ізолюють ці утвори в суміжних оматидіїв, запобігаючи проникненню променів світла до рабдомів через сусідні оматидії. У денних комах пігмент розміщений рівномірно по всій довжині пігментних клітин і повністю ізолює оматидіїв від сусідніх (опозиційний зір). Завдяки цьому до кожного оматидія можуть доходити лише ті промені, які йдуть через кришталик і збігаються з поздовжньою віссю оматидія, вони й досягають рабдома. Поле зору кожного оматидія дуже мале й сприймає лише малу частину предмета, який комаха роздивляє. Але велика кількість оматидіїв дозволяє різко збільшити поле зору взаємним прикладанням (апози- цією) один до одного. У результаті з окремих найменших частин складається, як у мозаїці, єдине загальне зображення.
У нічних комах, які активні за дуже слабкого освітлення, пігмент здатен переміщуватись і нагромаджуватись лише у верхніх частинах пігментних клітин, завдяки чому промені світла потрапляють на раб- доми не одного, а кількох сусідніх оматидіїв, що істотно підвищує чутливість ока до світла (суперпозиційний зір).
Латеральні (бічні) вічка (рис. 373, а) зустрічаються в личинок комах з повним перетворенням, а також в окремих імаго, наприклад, бліх, самців червеців. Вони містяться по боках голови в кількості до 30 з кожного боку. При переході комахи в дорослий стан вони руйнуються й замінюються на фасеткові очі. Дорсальні вічка зустрічаються в дорослих комах (як правило, в таких, які добре літають) й у личинок комах з неповним перетворенням. Найчастіше три дорсальні вічка розташовані у вигляді трикутника на верхній поверхні голови. Вічко має одну лінзу, під якою розміщені численні світлочутливі й пігментні клітини (рис. 373, б). Дорсальні вічка іннервуються не від зорових часток мозку, як фасеткові та латеральні, а від центральної частини про- тоцеребрума. Вони не виконують функції органів зору, але підвищують світлочутливість фасеткових очей і беруть участь у зоровій орієнтації комах, особливо під час польоту.
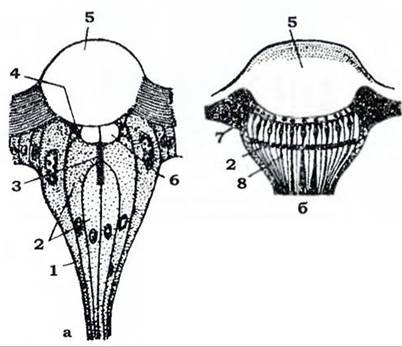
Рис. 373. Схема будови вічок комах
(з Мазохіна-Поршнякова й Тищенка): а - латерального; б - дорзального
1 - мантійна клітина; 2 - зорові клітини; 3 - рабдом; 4 - корнеагенні клітини; 5 - рогівка (кришталик);
6 - кристалічне тіло; 7 - пігментована гіподерма; 8 - пігментні клітини сітківки
Комахи мають кольоровий зір, найдосконаліший він у бджолиних і денних метеликів. Проте в комах, на відміну від людини, видима частина спектра захоплює також зону ультрафіолету (короткі хвилі); довгохвильова частина його, навпаки, коротша й закінчується на жовто-гарячому, не доходячи до червоного.
Комахи мають унікальну здатність до сприйняття поляризації світла. Денне світло поляризоване, проте людина не здатна сприймати цю поляризацію. Комахи, завдяки такій здатності, отримують змогу орієнтуватися по небу навіть тоді, коли воно затягнуте хмарами (астронавігація).
Статева система (рис. 374). У самки є пара яєчників, пара яйцепроводів і непарний яйцепровід (піхва), у який впадають придаткові залози. Часто є сім'яприймач та яйцеклад - кутикулярний пристрій для відкладання яєць у твердий субстрат. Кожен яєчник складається з яйцевих трубок, яких у різних комах може бути від однієї до 100, а інколи й більше, наприклад, у термітів їх більше 2400. Придаткові статеві залози виділяють секрет для приклеювання яєць до субстрату або утворення навколо групи яєць захисної оболонки - оотеки (таргани, богомоли) або ворочка (саранові).
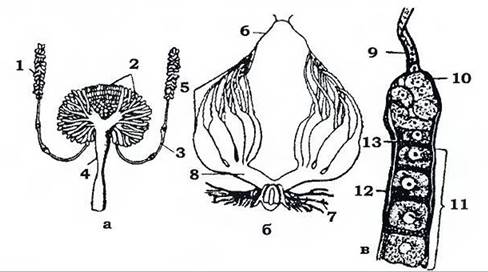
Рис. 374. Статева система таргана (із Шванвича): а - чоловіча; б - жіноча; в - кінцевий відділ яйцевої трубочки: 1 - сім'яник;
2, 7 - придаткові залози; 3 - сім'япровід; 4 - сім'явипорскувальний канал; 5 - яйцеві трубочки; 6, 9 - кінцева нитка; 8 - яйцепровід;
10 - гермарій; 11 - вітелярій; 12 - оболонка; 13 - фолікулярний епітелій
У самця є пара сім'яників, пара сім'япроводів, сім'явипорскувальний канал, придаткові залози й копулятивний орган. Часто копулятивний орган утворює зовнішні кутикулярні статеві придатки - геніталії, будова яких має видову специфічність і використовується в систематиці комах. Придаткові залози, які відкриваються у сім'явипорскувальний канал, у багатьох комах призначені для утворення сперматофора, який вводиться у статевий отвір самки чи прикріплюється до нього.
Розмноження. Як правило, для комах характерний чіткий статевий диморфізм, який виявляється в розмірах тіла, забарвленні, розмірах вусиків тощо, інколи особливо різко, наприклад, у самця жука- оленя, розвиваються величезні мандибули, а в жука-носорога - ріг. Проте часто обидві статі майже однакові й розрізнити їх можна лише за будовою статевих придатків (геніталій). У деяких комах (наприклад, попелиці) має місце партеногенез.
У комах над зародком, що розвивається, утворюються дві оболонки - зовнішня, або сероза, та внутрішня - амніон (рис. 375); вони захищають зародок від зовнішніх впливів.

Рис. 375. Один із способів утворення зародкових оболонок у комах (з Іванової-Казас):
а - зародок перед утворенням оболонок; б - початок їх утворення; в - завершення: 1 - головний кінець зародка; 2 - сероза; 3 — амніон
Метаморфоз має епіморфний характер: з яйця виходить личинка з усіма сегментами, яка більш-менш відрізняється від дорослої особини - імаго. Розрізняють два основні типи метаморфозу - неповне та повне перетворення.
За неповного перетворення, або геміметаболії, комаха проходить такі фази розвитку: яйце, личинка (яка зветься німфою) та імаго (рис. 376). З яйця виходить личинка, зовні схожа на дорослу особину. Вона має фасеткові очі, такий самий, як і в дорослих, ротовий апарат, а на більш пізніх стадіях - зовнішні зачатки крил. Німфи ведуть такий самий спосіб життя, що й дорослі комахи. Окрему модифікацію становлять так звані наяди - личинки одноденок, бабок і веснянок; вони також схожі на імаго, але живуть у воді й мають спеціальні личинкові органи - зябра, маску (личинки бабок) тощо. Ріст і розвиток личинки супроводжуються періодичними линяннями; з кожним линянням личинка за розмірами, пропорціями тіла, розвитком крил наближається до дорослої фази; під час останнього линяння вона перетворюється на імаго. У деяких нижчих комах (щетинкохвосток й одноденок) розвиток відбувається за типом архіметаболії: з яйця виходить передличинка з майже гомономною метамерією й недорозвиненими очима та придатками (антенами, хвостовими нитками тощо), яка живиться ембріональним жовтком. Вона линяє й перетворюється на личинку.
За повного перетворення, або голометаболії, комахи про ходять фази: яйце, личинка, лялечка, імаго. З яйця виходить личинка, яка різко відрізняється від імаго більшою гомономністю метамерії, відсутністю зовнішніх зачатків крил, слабшим розвитком органів чуття. Вона не має фасеткових очей, антени її значно коротші, часто ротовий апарат у неї інший, ніж у дорослої комахи (наприклад, у гусені метеликів - гризучий, а в імаго - сисний) (рис. 378). Личинки живуть в інших умовах, ніж дорослі. Більшість органів личинки має тимчасовий характер, наприклад, черевні ноги в личинок метеликів і пильщиків, шовковидільні залози в гусені метеликів, личинок деяких ос, їздців, пильщиків тощо. Зустрічаються червоподібні малорухомі личинки з м'якими покривами, інколи вони навіть не мають голови (личинки мух), часто безногі або зі слабо розвиненими грудними ногами (личинки жуків, перетинчастокрилих). Інші є більш рухливими, з трьома парами грудних і двома-вісьмома парами коротеньких черевних ніг (гусінь метеликів, личинки пильщиків). Рідше зустрічаються рухливі личинки з твердими покривами, добре розвиненими грудними кінцівками й ротовим апаратом з гострими щелепами. Це хижі личинки жуків-турунів, плавунців, сітчастокрилих. Личинка росте, кілька разів линяє й на останній стадії припиняє живлення, стає нерухомою, линяє востаннє й перетворюється на лялечку. Часто перед залялькуванням личинка оточує себе коконом із шовку чи часточок субстрату, скріплених шовком або виділеннями мальшпєвих пильщиків, їздців). Кокон захищає лялечку від несприятливих впливів. Багато личинок жуків і гусені метеликів заляльковуються у ґрунті, утворюючи порожнину - комірку, або колисочку, часто укріплену екскрементами. Іноді місцем залялькування є стебла рослин, скручене листя. Буває й відкрите залялькування, наприклад, у денних метеликів.
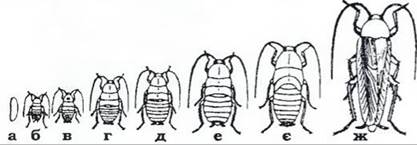
Рис. 376. Неповний метаморфоз прусака Blattella germanica (із Шванвича): а - яйце; б-є - шість личинкових стадій (німфи); ж — імаго
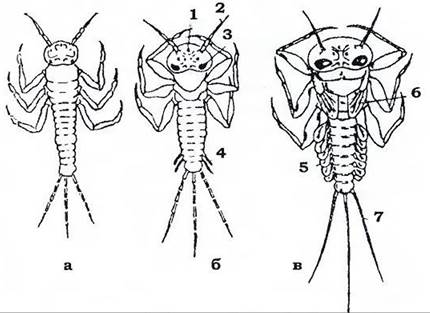
Рис. 377. Окремі фази архімєтаболії одноденок: а - "личиночка’'; б - личинка; в - німфа:
1 - дорзальне вічко; 2 - антена; 3 - фасеткове око; 4 - зачатки зябер; 5 - зябра; б - зачатки крил; 7 - хвостові нитки
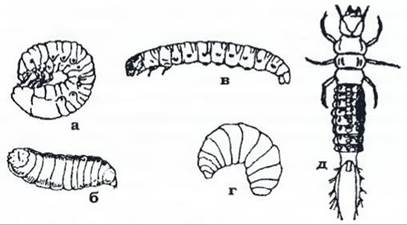
Рис. 378. Личинки комах з повним метаморфозом:
а - травневого хруща; б - мухи; в - метелика судин (у метеликів, (гусінь); г - бджоли; д - жука-туруна (камподеоподібна личинка)
Лялечка найчастіше нерухома (за деякими винятками) й нездатна до живлення, вона живе за рахунок нагромаджених личинкою резервів. Розрізняють три типи лялечок: відкриті, покриті й приховані (рис. 379). Лялечка нерухома, але на цій фазі відбуваються інтенсивні процеси внутрішньої перебудови, яка включає гістоліз і гістогенез. За гістолізу личинкові органи руйнуються шляхом фагоцитозу, автолізу (самоперетравлення), а також за допомогою ферментів. Паралельно з гістолізом відбувається гістогенез - побудова імагінальних органів, які формуються з особливих зачатків - ілюгінальних дисків. Це невеличкі скупчення недиференційованих клітин, які закладаються ще під час ембріонального розвитку або в ранніх личинок і розміщені в певних місцях тіла личинки. Є диски, з яких утворюються крила, кінцівки, певні ділянки кишечнику, фасеткові очі тощо. Упродовж личинкового розвитку вони ростуть,
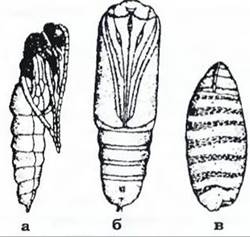
Рис. 379. Типи лялечок:
а - відкрита (їздця); б - покрита (метелика); в - прихована (мухи)
Метаморфоз комах відбувається під контролем гормональної (ендокринної) системи (рис. 369). Нейросекреторні клітини мозку виробляють активаційний гормон, через довгі аксони цих клітин він надходить до кардіальних тіл, а з них - у гемолімфу. У личинок активаційний гормон стимулює роботу проторакальних залоз, які починають виділяти гормон линяння - екдизон. Останній впливає на клітини гіподерми, викликаючи спочатку синтез ферментів, які розчиняють стару кутикулу, а пізніше - синтез матеріалу для побудови нової кутикули. Крім того, екдизон стимулює ріст і диференціацію всіх тканин личинки, особливо в гонадах, імагінальних дисках тощо. Паралельно з екдизоном при личинкових линяннях виділяється ювенільний (юнацький) гормон, який гальмує метаморфоз, тобто перешкоджає линянню личинки й переходу її в дорослу фазу. За високої концентрації ювенільного гормону гальмується розвиток імагінальних органів, і линяння личинки приводить до утворення личинки наступної стадії. Зниження концентрації ювенільного гормону спричинює линяння личинки й перетворення її на лялечку (або останню личинкову стадію за неповного метаморфозу). Припинення надходження ювенільного гормону в гемолімфу зумовлює линяння лялечки (або останньої личинкової стадії за неповним метаморфозом) в імаго.
Політ. Комахи - єдина група безхребетних тварин, здатна до активного польоту. Політ комах - це складний фізіологічний процес, який включає роботу крил, м'язів, гангліїв нервової системи, органів чуття, дихальної системи тощо.
Крило комахи під час руху опирається на виріст плейрита - стовпчик, який розташований поблизу від місця його прикріплення. Завдяки цьому крило стає двоплечим важелем, довге плече якого - це крилова пластинка, а коротке - невеличка ділянка біля його основи (рис. 363). Рух крил під час польоту забезпечується м'язами непрямої дії, які безпосередньо не прикріплюються до основи крила. При скороченні дорсовентральних м'язів опускається тергіт, який тисне на основу крила, підіймаючи крилову пластинку вгору. Поздовжні м'язи, розташовані дорсально вздовж грудей, при скороченні вигинають тергіт догори, при цьому крило опускається. Таким чином забезпечується поступальний рух. Важливе значення при цьому має пружність кутикули тергітів, які після скорочення м'язів набувають початкової форми. М'язи прямої дії приєднані безпосередньо до основи крила та склеритів грудей. За їхньою допомогою крило повертається вздовж поздовжньої осі, змінюючи кут нахилу, а також відводиться вперед чи назад, що регулює швидкість і напрямок польоту. Лише в бабок (ряд Odonata) у польоті беруть участь майже виключно м'язи прямої дії. Під час руху крил уперед і вниз у горизонтальній площині виникає підйомна сила, а при русі назад і вгору крило стає вертикально, й створюється пропелерний ефект. Велика частота помахів забезпечує поєднання підйомного та поступального ефектів, що й дозволяє комасі летіти вперед.
У більшості комах у польоті беруть участь або задні (коли передні крила перетворені на надкрила), або передні (у двокрилих) крила. Часто під час польоту передні та задні крила зчіплюються й діють як єдине ціле (метелики, перетинчастокрилі). У бабок кожне крило діє автономно, чим досягається виключна швидкість і маневреність польоту.
Швидкість польоту в комах є різною: у кімнатної мухи вона не перевищує 8 км за годину, у сарани - 12,6 км, у джмеля - 18 км, у бражника - 54 км, а в деяких бабок - навіть до 100 км за годину. Кількість помахів крила за секунду коливається від 5-10 (денні метелики) до кількох сотень у двокрилих, а у комарів-дзвінців (родина Chironomidae) і дрібних комарів-мокреців (родина Ceratopogonidae) - до 1 тис. помахів, що пояснюється здатністю крилових м'язів відповідати кількома скороченнями на один нервовий імпульс.
Поведінка. Комахи характеризуються складними формами поведінки. Перш за все, це стосується комплексу інстинктів турботи про нащадків. У найпростіших випадках це відкладання самками яєць біля харчового субстрату личинок або просто в нього (більшість метеликів, мухи, жуки тощо); складніша форма - створення запасів їжі для потомства та будова тих чи інших схованок, де розвивається молодь. Найскладніша поведінка в суспільних комах, які живуть великими сім'ями. Це терміти, деякі бджоли, оси, джмелі та мурашки. Регуляція соціальної поведінки суспільних комах здійснюється за допомогою феромонів. Феромони - це біологічно активні речовини, які виділяються тваринами у зовнішнє средовшце й викликають в особин того самого виду специфічну реакцію (певну поведінку або процес розвитку). Наприклад, мандибулярні залози бджолиної матки виділяють так звану "царську речовину", яка виконує множинну функцію: пригнічує в робочих бджіл розвиток яєчників і рефлекс будівництва "царських комірок" для вирощування майбутніх маток, приваблює робочих бджіл до матки, приваблює самців (трутнів) під час шлюбного польоту тощо. Аналогічні феромони виробляють самки мурашок і самки та самці термітів. У суспільних комах виник спадково запрограмований код, завдяки якому комбінації невеликої кількості (не більше 10) різних феромонів забезпечують координацію складної діяльності особин одного виду. У мурашок виділення феромонів можна розглядати навіть як своєрідну "хімічну мову". У різних частинах їхнього тіла розміщені численні залози, що виробляють різні феромони.
Усі перелічені форми поведінки не потребують навчання, це генетично закріплені інстинкти. Проте велику роль у поведінці комах, особливо суспільних, відіграють також умовні рефлекси, які виявляються в запам'ятовуванні місцезнаходження джерела їжі, дороги до нього та назад до гнізда. Медоносних бджіл можна навчити прилітати на той чи інший кольоровий субстрат або намальовану на ньому фігуру. Навіть у таких одиночних комах, як таргани, можна виробити умовні рефлекси.
Клас Insecta поділяється на два підкласи.