ЗООЛОГІЯ БЕЗХРЕБЕТНИХ - Г. Й. Щербак - 2008
ПРОТИСТИ - PROTISTA
НАЙПРОСТІШІ (PROTOZOA)
АЛЬВЕОЛАТИ
ТИП ВІЙКОНОСНІ, або ІНФУЗОРІЇ (CILIOPHORA)
Переважно вільноживучі морські та прісноводні найпростіші; рідше зустрічаються мешканці ґрунту, коменсали та паразити. Тип нараховує приблизно 8 тис. видів. Розміри інфузорій порівняно великі - 50-300 мкм, окремі види досягають 3 мм.
Інфузорії відрізняються від інших найпростіших наявністю органел руху - війок, ядерним, дуалізмом, складно збудованим покривом - кортексом, який притаманний лише інфузоріям, та особливою формою статевого процесу - кон'югацією.
Будова. Форма тіла інфузорій є надзвичайно різноманітною (рис. 31). Плаваючі види мають обтічну форму: округлу, овальну, паличкоподібну; повзаючі - сплющену; сидячі - форму дзвону, сурми; вони можуть мати й розгалужене тіло, а паразитичні види - й різні відростки. Зовні клітина вкрита кортексом (рис. 32). У кортексі містяться різноманітні структури, які разом забезпечують сталість форми клітини. Зовнішній шар кортексу - пелікула - утворений плазмалемою та розташованими під нею сплющеними мішечками (альвеолами), мозаїчно з'єднаними між собою. Під пелікулою розміщуються базальні тільця (кінетосоми) війок, а також пов'язані з кожною з кінетосом утвори: смугастий філамент, спрямований до переднього кінця клітини, та пучки мікротрубочок, з яких один іде вздовж тіла до його заднього кінця, а другий - поперек клітини. Сукупність мікрофіламентів і мікротрубочок утворює єдиний цитоскелет, властивий лише інфузоріям.
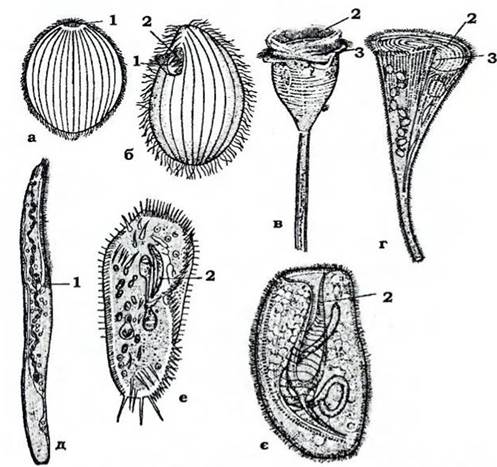
Рис. 31. Інфузорії (а, б, г-е - з Догеля; в - з Полянського): а - Holophrya; б - Telruhymena pyriformis; в - Vorticellusp.;
г - Stentor polymorphus; д - Spirostomum ambiguum; e - Stylonichia mytilus; є - Bursana truncatella:
1 - цитостом; 2 - мембранели; 3 - перистом
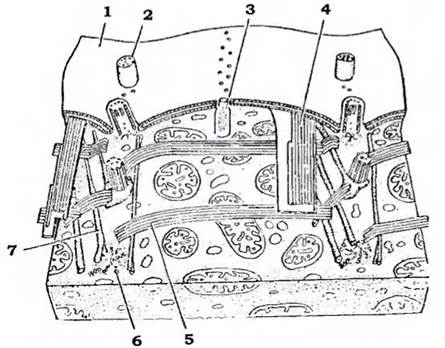
Рис. 32. Ультраструктура кортекса інфузорій (з Догеля):
1 - пелікула; 2 - війка; 3 - трихоциста; 4 - поздовжні мікротрубочки;
5 — поперечні мікротрубочки; б - кінетосома; 7 - мікрофіламент
В ектоплазмі перпендикулярно до її поверхні розташовані захисні органели - трихоцисти, які при подразненні "вистрілюють", перетворюючись на довгу нитку. Вважають, що вони містять отруйні речовини, які вбивають ворогів чи здобич. У хижих інфузорій є токсоцисти. Вони мають вигляд капсули з оболонкою з трубочок, у порожнині якої є внутрішня трубка. При полюванні ця трубка вивертається назовні, як палець рукавички, або телескопічно висувається з капсули, пронизує тіло здобичі, в яке впорскується отрута. Це паралізує чи вбиває здобич (інші найпростіші, коловертки).
Органели руху - війки - принципово не відрізняються від джгутиків, вони мають таку саму ультраструктуру. Війки можуть рівномірно вкривати всю поверхню тіла, проте часто вони концентруються на певних ділянках або об'єднуються, утворюючи цири, мембрани або мембранели. Цири - пучечки або китиці з війок, мембрани - це ряди війок, які щільно прилягають одна до одної, утворюючи пластинки, а мембранели - це щільний ряд війок (рис. 31). Війки рухаються веслоподібно, узгоджено. За допомогою війок інфузорії можуть плавати, бігати по субстрату чи "стрибати" в товщі води, відштовхуючись цирами, заганяти їжу до рота мембранелами та мембранами. Особливо складною є будова війкового апарату, пов’язаного із живленням.
Для багатьох війчастих на певних етапах життєвого циклу характерним є розвиток прикріпних апаратів. Наприклад, в інфузорій роду Vorticella прикріпне стебельце утворюється особливою органелою - скопулою, яка секретує це стебельце (рис. 31, в). А в паразитів риб - інфузорій роду Tnchodina - є особливий "присосок", утворений білковими структурами.
Ядерний апарат інфузорій складається з двох типів ядер (ядерний дуалізм): найчастіше одного великого - макронуклеуса та кількох дрібних - мікронуклеусіє. Макронуклеус багатий на ДНК і містить велику кількість ядерного матеріалу, в ньому відбувається синтез РНК, він регулює клітинний метаболізм. Мікронуклеуси є диплоїдними й беруть участь лише у статевому процесі.
Більшість інфузорій має клітинний рот (цитостом) і складну систему органел травлення (рис. 30). У деяких видів цитостом розташований на передньому кінці клітини найчастіше збоку, на дні особливого заглиблення - перистома (рис. 31) й оточений звичайними війками (Holophrya, Prorodon). У деяких видів перистом веде до вузького каналу - клітинної глотки, або цитофаринкса, що закінчується в ендоплазмі (рис. 33).
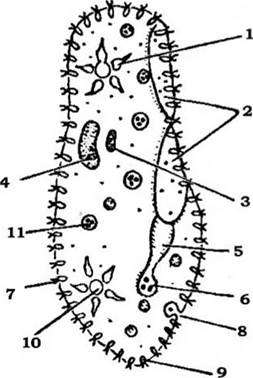
Рис. 33. Схема будови Paramecium caudatum:
1 - передня скоротлива вакуоля; 2 - перистом; 3 - мікронуклеус;
4 - макронуклеус; 5 - цитофаринкс; 6 - цитостом; 7 - війки; 8 - цитопрокт; 9 - трихоциста;
10 - задня скоротлива вакуоля; 11 - травна вакуоля
Війки перистома та цитофаринкса утворюють мембрани й мембранели, які заганяють часточки їжі до рота. У хижих інфузорій (Didinium) замість війок функціонує складне пристосування цитостома для заковтування здобичі - паличковий апарат (рис. 34). Він складається із сотень мікротрубочок, які утворюють навколо рота трубку, що виконує опорну функцію при заковтуванні великої здобичі (найпростіших, коловерток тощо). На дні цитостома грудочки їжі оточуються мембраною, й утворюється травна вакуоля, яка перемішується в ендоплазмі інфузорії, де їжа поступово перетравлюється, а неперетравлені рештки видаляються через спеціальний утвір у пелікулі - порошицю.
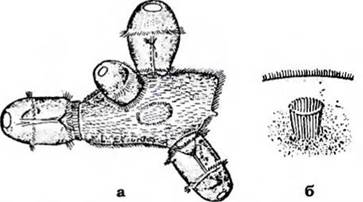
Рис. 34. Didinium nasutum, які поїдають іифузорію-туфельку (а) та їхній паличковий апарат (б) (з Догеля, зі змінами)
Сисні інфузорії (Suctoria) не мають цитостома, вони висмоктують свою здобич сисними щупальцями, в яких є трубки, складені з мікротрубочок (рис. 35). Паразитичні інфузорії з ряду Astomatida також не мають цитостома.
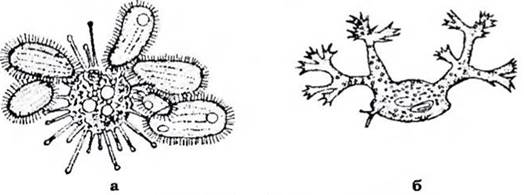
Рис. 35. Сисні інфузорії:
а - Sphaerophryu magna, що висмоктує одночасно п'ять інфузорій; б - Dendrocometes paradoxus (з Догеля, зі змінами)
Прісноводні, а часто й морські інфузорії мають скоротливі вакуолі - органели осмореіуляції та виділення. У багатьох випадках їхня будова ускладнюється наявністю зірчасто розташованих привідних канальціе з розширеннями - ампулами, в які збирається надлишок рідини. З ампул рідина надходить до скоротливої вакуолі, а з неї викидається назовні. Фіксованого положення цьому комплексу надають стрічки з мікротрубочок, які починаються від стінок видільної пори та йдуть уздовж ампул і каналів (рис. 36). Більшість інфузорій має клітинний рот (цитостом) і складну систему органел травлення (рис. 30). У деяких видів цитостом розташований на передньому кінці клітини найчастіше збоку, на дні особливого заглиблення - перистома (рис. 31) й оточений звичайними війками (Holophrya, Prorodon). У деяких видів перистом веде до вузького каналу - клітинної глотки, або цитофаринкса, що закінчується в ендоплазмі (рис. 33).
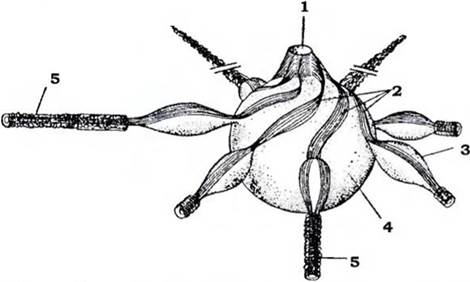
Рис. 36. Схема будови скоротливої вакуолі Paramecium (із Хаусмана): 1 - пора; 2 - стрічки мікротрубочок;
3 - ампула; 4 - скоротлива вакуля; 5 - привідні канали
Розмноження. Нестатеве розмноження відбувається шляхом поділу клітини навпіл або брунькуванням, при цьому як у мікро-, так і в макронуклеусах відбувається мітоз, хоч в останніх він носить прихований характер. Частина органел розподіляється між дочірніми клітинами, а ті, яких не вистачає, відновлюються. При брунькуванні від великої материнської клітини відбруньковується менша, дочірня (сувійки, сисні інфузорії).
Статевий процес - кон'югація - притаманний лише інфузоріям. Він не супроводжується утворенням гамет, як у більшості найпростіших, а полягає в тимчасовому попарному з'єднанні особин й обміні між ними ядрами (рис. 37). Розглянемо цей процес на прикладі інфузорії- туфельки з одним макро- та одним мікронуклеусом. Дві інфузорії з'єднуються бічними поверхнями, їхні перистоми зв'язані цитоплазматичним містком. У кожної особини макронуклеус руйнується, а диплоїдиий мікро- нуклеус поділяється мейотично, утворюючи чотири гаплоїдні ядра. Три з них руйнуються, а четверте мітотично ділиться на стаціонарне (жіноче) та мігруюче (чоловіче) ядра. Партнери обмінюються мігруючими ядрами, кожне з яких зливається зі стаціонарним іншої особини. Так утворюється дигілоїдний синкаріон. Після цього клітини розходяться. Згодом у кожному екс-кон'юганті із синкаріона шляхом послідовних мітозів утворюються вісім ядер. З них три гинуть, одне стає мікрону клеусом, а чотири - зачатками макрону- клеусів. Мікронуклеус мітотично ділиться, що супроводжується поділом клітини на дві, кожна з яких має один мікронуклеус і два зачатки макронуклеусів, і цей процес повторюється. Наприкінці утворюються чотири клітини, і кожна містить один мікронуклеус і зачаток макронуклеуса з диплоїдними наборами хромосом. У зачатку макронуклеуса більша частина хромосом розчиняється, а ті, що залишились, розпадаються на дрібні фрагменти, більша частина яких також руйнується. Потім починається інтенсивний синтез ДНК, що веде до багаторазового подвоєння фрагментів, що залишились, у сотні та тисячі разів. Отже, хоч макронуклеус і містить величезну кількість ДНК (порівняно з мікронуклеусом), його не можна вважати поліплоїдним, тому що в ньому залишається лише незначна частина (1,6 %) повного складу геному, але багаторазово помножена.
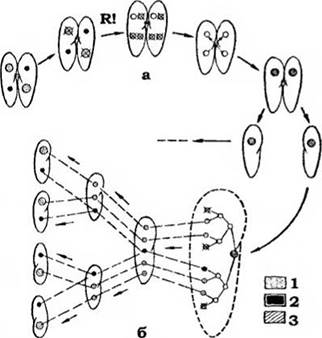
Рис. 37. Послідовні стадії змін ядерного апарату (а) та його відновлення (б) у Paramecium caudatum у процесі кон'югації:
1 - макронуклеус; 2 - мікронуклеус; З - синкаріон; R! - редукційний поділ
Якщо інфузорія не має партнера для кон'югації, статевий процес проходить у неї шляхом автогам. Її чоловіче та жіноче ядра зливаються всередині однієї клітини. Біологічний сенс кон'югації - обмін спадковим матеріалом між особинами, що веде до комбінативної мінливості. Урізноманітнення спадкового матеріалу популяції інфузорій, що мешкають у певному середовищі, підвищує здатність організмів пристосовуватись до несприятливих змін умов довкілля. Істотне значення має також відновлення макронуклеуса: при поділах кількість ДНК у ньому зменшується, що сповільнює розвиток і ріст. Життєвий цикл інфузорій відбувається з галіетичною редукцією хромосом. Тип Інфузорії поділяється на три класи.