ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
РОЗДІЛ ПЕРВИННІ БАГАТОКЛІТИННІ (PROMETAZOA)
ТИП ГУБКИ, АБО ПОРИФЕРИ (SPORGIA ABO PORIFERA)
Губки — це нижчі багатоклітинні, нерухомо прикріплені до субстрату тварини, що живляться шляхом фільтрації. Вони мешкають у водоймах, переважно в морях: утворюють колонії. Відомо понад 5 тис. сучасних і понад 2 тис. викопних видів. У прісних водоймах України виявлено не більше десяти видів у Чорному морі через низьку солоність - лише 29 видів, ще менше їх у Азовському морі.
Губки не мають оформлених тканин і органів, їх- тіло складається з безлічі клітин, що виконують різні функції
та мають різну будову, й міжклітинної речовини — продукту виділення цих клітин. Отже, губки — тварини з поклітинним диференціюванням, або дотканинні організми, їх зовнішня та внутрішня будова настільки незвичайні, що тривалий час учені не знали, до якого царства живих істот вони належать, і лише в 1825 р. їх було визначено як тварин.
За формою тіла губки надзвичайно різноманітні. Поодинокі губки мають переважно кубкоподібну форму, однак колонії утворюють обростання на субстратах у вигляді кущів, корка або грудок. Розміри губок коливаються від 1—2 мм до 2 м, багато видів забарвлені спеціальними пігментами в жовтий, коринневий, жовтогарячий, червоний, зелений чи фіалковий кольори.
Тіло губок побудоване з двох шарів клітин: пінакодерми, що вкриває тіло ззовні, та хоанодерми, яка вистилає парагастральну порожнину або джгутикові камери. Між цими двома шарами розташований мезохіл (паренхіма, мезенхіма), який складається з різноманітних клітин і продуктів їх виділення, він містить також скелетні елементи. Товща мезохіла пронизана каналами, що відкриваються на поверхні невеличкими отворами — порами. Всю різноманітність будови губок можна поділити на три типи— аскон, сикон, лейкон. Це — назви родів, що -мають відповідну будову (рис. 69).
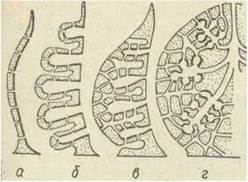
Рис. 69. Типи будови губок:
а — аскон; б —простий сикон; в — ускладнений сикон; г — лейкон
Губка типу аскон, наприклад Ascetta, має вигляд бокала чи мішечка, прикріпленого основою (підошвою) до субстрату, з отвором (оскулюмом) на протилежному верхньому полюсі. Отвір веде у внутрішню (парагастральну, або атріальну) порожнину (рис. 70).
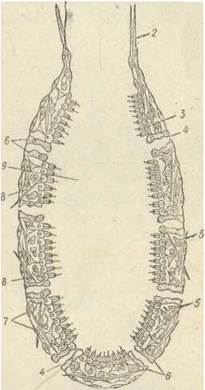
Рис. 70. Будова губки типу аскон:
1 — оскулюм; 2 —голки; 3 — хоаноцити; 4— пори; 5 — амебоцити; 6— пороцити;
7 — пінакоцити; 8 — коленцити; 9 — парагастральна порожнина
Подальші ускладнення будови губок пов'язані з розростанням мезохіла та випинанням в нього ділянок атріальної порожнини, що утворюють радіальні канали, стінки яких вистелені хоанодермою. Це сиконоїдний тип, що трапляється в багатьох поодиноких губок (Sycon raphanus).
Для більшості губок характерний лейконоїдний тип будови: стінка тіла потовщується, хоаноцити концентруються в джгутикових камерах, які містяться в товщі паренхіми (мезохіла) й зв'язані із зовнішнім середовищем і атріальною порожниною системою розгалужених канальців.
Канальці, що сполучають камеру з зовнішнім середовищем, називаються привідними, а парагастральною порожниною — відвідними. Камери можуть утворювати кілька шарів, значно збільшуючи об'єм тіла губки. Таким чином утворюється складна іригаційна .Система, що спрямовує та впорядковує потік води.
У губок типів сикон та лейкон парагастральна порожнина та канали вистелені пінакодермою, як ї зовнішня поверхня. Клітинний склад губок вивчений досить добре, а останнім часом ці відомості доповнені електронно-мікроскопічними дослідженнями (рис. 71). До складу пінакодерми входять два типи клітин: пінакоцити та породити.
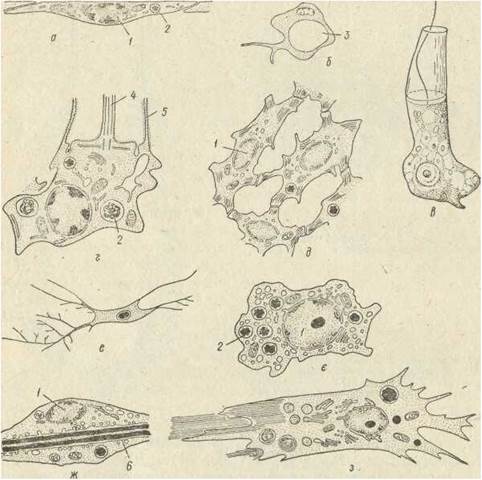
Рис. 71. Клітини губок — пінакоцит (а), пороцит (б), хоаноцит — загальний вигляд (е) і внутрішня будова (г), міоцити (д), коленцит (е), археоцит (є), склероцит (ж), лофоцит (з): 1— ядро; 2 — травна вакуоля; 3 — канал у пороциті; 4 — джгутик; 5 — мікроворсинка комірця; 6 — спікула
Пінакоцити мають сплющену або т-подібну форму з плоскою поверхневою частиною та цитоплазматичним відростком із ядром та іншими органоїдами, зануреними в товщу мезохіла (рис. 71, а).
Пінакоцити, щільно прилягаючи оди» до одного, утворюють зовнішній покрив губок і вистилають їх внутрішні порожнини (крім джгутикових камер). У деяких губок (рогових) поверхня тіла вкрита ще й захисною оболонкою — кутикулою, яка вириває шар піонакоцитів. У цитоплазмі пінакоцитів, крім ядра, великої кількості мітохондрій, лізосом, локалізовані також великі травні вакуолі (фагосоми), що свідчить про їх здатншть до фагоцитозу. Цитоплазма пінакоцитів містить також мікрофіламеній, які зумовлюють їх скоротливість. Завдяки цьому форма пінакоцитів може змінюватися.
Пороцити — це різновид пінакоцитів, клітини, пронизані поровим каналом (рис. 71,6). Пороцит здатний скорочуватися й може відкривати або закривати пору та регулювати діаметр каналу.
Хоанодерма складається з комірцевих клітин хоаноцитів. Це циліндричні клітини з одним джгутом (рис. 71, в, г.). Навколо основи джгута від поверхні клітини відходять близько 2000 ворсинок, які під оптичним мікроскопом мають вигляд комірця. Джгутик всередині цитоплазми починається базальним тільцем (кінетосомою), від якого радіальне відходять мікротрубочки. За рахунок роботи джгутиків хоаноцити створюють рух води через тіло губки. Крім того, в хоаноцитах вловлюються часточки їжі, бактерії, водорості тощо. Харчові частки захоплюються біля основи мікроворсинок, де формуються піноцитозні та фагоцитозні вакуолі. В цитоплазмі хоаноцитів постійно містяться фагосоми.
Мезохіл, або паренхіма, складається з міжклітинної речовини, густо заселеної різними типами клітин і скелетними елементами. Серед клітин мезохіла найбільш поширеними є амебоцити (археоцити), які мають амебоподібну форму, рухаються за допомогою псевдоподій і характеризуються фагоцитарною активністю (рис. 71, є). Вони захоплюють і перетравлюють бактерії та інші харчові частки, одержані від хоаноцитів або захоплені крізь просвіти в хоано- або пінакодермі. Амебоцити поїдають також старі або відмираючі клітини в мезохілі. Завдяки своїй рухливості амебоцити також розносять поживні речовини по тілу губки. Амебоцити здатні перетворюватися на інші типи клітин (хоаноцити, пінакоцити, статеві клітини тощо).
Міоцити — це спеціалізовані веретеноподібні клітини з довгими відростками клітини, які розташовані навколо оскулюма та крупних каналів іригаційної системи (рис. 71, д). їх цитоплазма містить велику кількість паралельно орієнтованих мікротрубочок і мікрофіламентів. Але м'язовими ці клітини назвати не можна, бо в них немає справжніх м'язових волокон. Крім того, вони не зв'язані з нервовими клітинами, яких у губок немає. Вчені припускають, що здатність до проведення подразнення та скорочень притаманна самим міоцитам, які формують складну сітку, з'єднуючись між .собою за допомогою довгих відростків. Висловлюється також думка, що саме сітка міоцитів виконує інтегративні функції в організмі губок.
Збудження через таку сітку проводиться в.десятки разів повільніше, ніж через нервові клітини кишковопорожнинних.
Коленцити — нерухомі клітини зірчастої або веретеноподібної форми з довгими розгалуженими відростками (рис. 71, е). Основною функцією коленцитів є виділення колагенових волокон, що спочатку містяться всередині коленцитів у особливих вакуолях, де виконують опорну, функцію, а потім виділяються в мезохіл.
Лофоцити — великі рухливі клітини, локалізовані переважно в базальній частині губки (рис. 71, з). Вони характеризуються продукуванням великої кількості колагенів пучки якого тягнуться за клітиною, що рухається за допомогою псевдцтюдій. За функціями лофоцити подібні до коленцитів, але виділяють не поодинокі, а зібрані в пучки колагенові волокна. Коленцити та лофоцити здатні до фагоцитозу, в їх цитоплазмі трапляються фагосоми.
Спонгіоцити — спеціалізовані клітини, які продукують скелетний білок—спонгін. Це дрібні клітини з добре розвиненим шорстким ендоплазматичним ретикулюмом та апаратом Гольджі; що активно синтезує речовину — попередник спонгіту. Вона міститься в особливих пухирцях, а потім виділяється з клітини. В утворенні спонгінового волокна беруть участь кілька клітин, які збираються разом і садретують епонгія,
Склероцити — клітини, що утворюють елементи мінерального скелета — спікули. Цитоплазма склероцитів містить численні вакуолі, в яких накопичується фібрилярний матеріал, з якого утворюється осьова нитка (рис. 71, ж). Потім навколо осьової нитки відкладається кремнезем. Отже, ці спікули утворюються всередині склероцитів. В утворенні вапнякової спікули беруть участь щонайменше дві клітини, а в утворенні три-, чотири- або багатоосьових опікул — шість і більше.
Крім зазначених клітин, у мезохілі губок містяться спеціалізовані амебоїдні клітини з різними включеннями в цитоплазмі (сферулярні, мікрогранулярні, рабдитні клітини тощо). Вони трапляються не у всіх губок, а лише у певних видів або екологічних груп.
Майже всі клітини можуть перетворюватися на клітини інших типів. Археоцити здатні диференціюватися в усі Інші клітинні типи. З пінакоцитів вапнякових губок розвиваються склероцити, а з їх хоаноцитів — статеві клітини, пінакоцити, амебоцити.
Подразливість у губок дуже обмежена. Вони не здатні ні до активних рухів, ні до зміни форми тіла. Лише пори та оскулум можуть дуже повільно змінювати діаметр завдяки скороченню міоцитів і пороцитів. Така нерухомість компенсується течією води крізь тіло губки. Джгутики хоалоцитів, рухаючись, забезпечують постійну течію води через пори та канальці в парагастральну порожнину, звідки вона виходить назовні через оскулум. Разом з водою до тіла губки надходять дрібні органічні частки — залишки організмів, одноклітинні водорості, бактерії, напростіші.
У більшості вапнякових губок внутріклітинне травлення відбувається в травних вакуолях хоаноцитів (фагосомах), однак частина їжі передається амебоцитам. У зви-
чайних і скляних губок вся їжа потрапляє до амебоцитів, хоаноцити її лише відловлюють, однак не перетравлюють. У деяких губок хоаноцити лише підганяють поживні частки, а їх вловлювання та перетравлення здійснюється амебоцитами в стінках каналів. Пінокицити також здатні захоплювати часточки їжі з мезохіла та перетравлювати їх. Амебоцити мігрують в товщі мезохіла, виконуючи функції розподілу їжі. Неперетравлені рештки їжі передаються мігруючими клітинами пінакоцитам, які викидають їх у вивідні канали (рис. 72).
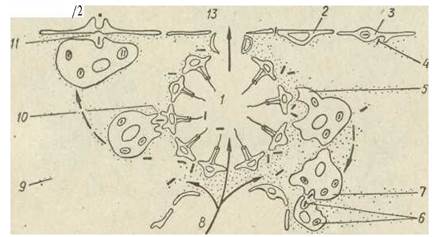
Рис. 72. Схема поглинання харчових часток у Ephydatia: 1— коаноцити; 2—породити; 3 — пінакоцити; 4 — захоплення їжі пінакоцитом з меаохіла; 5 — археоцит, що поглинає бактерії; 6 —бактерії; 7 —передача їжі від одного археоцита до другого; 8 — ввідний канал; 9 — товща мезохіла; 10 — передача їжі від хоаноцита до археоцита; 11 — те ж від археоцита до пінакоцита; 12— виведення неперетравлених решток; 13— вивідний канал
Губки дихають розчиненим у воді киснем, який проникає в клітини завдяки течії води через тіло тварини. Продукти обміну виділяються клітинами безпосередньо у воду або в міжклітинну речовину паренхіми, звідки вони також дифундують у воду й з нею через оскулум виводяться назовні. Отже, в губок всі фізіологічні функції, зокрема й подразливість, виконують, як і в найпростіших, окремі клітини, а не організм у цілому.
Переважна більшість видів губок має скелет, що є опорою тіла та стінок каналів, однак не виконує рухової функції. Він буває вапняковим, кремнеземним (з диоксиду кремнію) або роговим (із спонгіну). Склад і будову скелета покладено в основу класифікації губок. Мінеральний скелет (вапняковий чи кремнеземний) складається з великої кількості голок, або спікул, які мають різноманітну форму та по-різному розташовані в тілі губок (рис. 73). Скелет, як правило, міститься в товщі мезохіла, однак іноді гелки стирчать назовні (захисне пристосування). Спікули утворюються всередині особливих клітин — склероцитів. Після формування спікули клітина відмирає. Спікули можуть бути одно-, три- чи багатоосьовими або мати більш складну будову зірочок, шпильок, дужок, якірців тощо. Інколи спікули з'єднуються в суцільний скелетний каркас, що найбільш характерно для скляних губок.
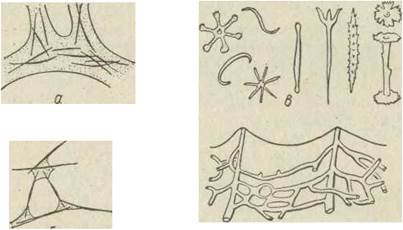
Рис. 73. Скелетні елементи губок: а — спікули в спонгіновому тяжі; б— спікули, з'єднані спонгіном; в — типи спікул; г — спонгіновий скелет туалетної губки.
У деяких губок, зокрема у викопних археоціат, скелет монолітний.
Органічний (роговий) скелет складається з колагенових волокон і спонгіну—речовини, за хімічним складом близької до шовку. Спонгінові волокна з'єднують окремі спікули в єдиний скелет.
Губки розмножуються статевим і нестатевим шляхом. Нестатеве—-вегетативне — розмноження відбувається шляхом зовнішнього чи внутрішнього брунькування, поздовжнього поділу та фрагментації.
Під час зовнішнього брунькування на тілі материнської особини утворюється брунька, в яку проростають усі шари материнського організму та продовжується парагастральна порожнина (рис. 74). У інших випадках брунька виникає зі скупчення археоцитів. У поодиноких видів (наприклад, із родів Ascetta, Sycon тощо) з бруньки формується молода особина з власним оскулумом. Певний час вона утримується на материнському організмі, потім падає на дно та починає самостійне життя. У колоніальних видів, до яких належить більшість губок, дочірні особини протягом життя зв'язані з материнською, причому часто окремі тварини зливаються, й такі колонії набувають найрізноманітніших форм (рис. 75). У цьому разі лише число оскулумів вказує на кількість особин колонії. У деяких губок індивідуальність особин пригнічується й замінюється індивідуальністю колонії. Так, колонії у багатьох видів набувають форми бокала або чаші, повторюючи форму поодинокої особини, при цьому оскулуми розташовуються правильно або навіть зливаються в один. Це — вторинно поодинокі губки, тобто колонії, що імітують форму індивидів, однак вони мають значно більші розміри та складнішу іригаційну систему.
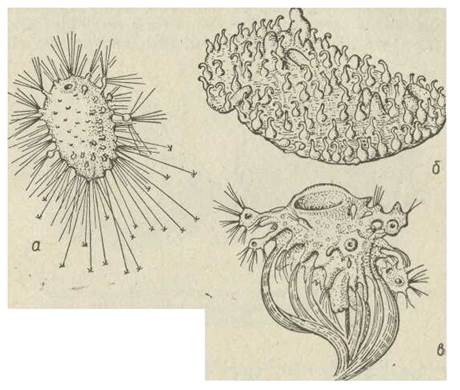
Рис. 74. Зовнішнє брунькування у скляних губок: й — Anoxycalus ijimae; б — Polymastia mammillaris; e — Tethya aurantium

Рис. 75. Колоніальні та вторинно поодинокі губки: а — Mycale ochotensis; б —Sycon ciliatum; в —Phakellia eribrosa
Внутрішнє брунькування характерне для прісноводної губки бодяги. Воно відбувається в паренхімі. Група амебоцитів, багатих на поживні речовини, відокремлюється від інших клітин за допомогою щільної оболонки складної будови, яка продукується коленцитами. Вона складається з двох шарів органічної речовини (спонгін), між якими розташований пінистий шар, що містить повітря. У деяких видів бодяг у пінистому шарі локалізовані особливі скелетні елементи — амфідиски, що мають вигляд коротеньких паличок із зіркоподібними дисками на кінцях. Вони запобігають злипанню спонгінових шарів, а пінистий шар створює надійну термоізоляцію. Така зимуюча брунька називається гемулою (рис. 76). Протягом літа в кожній губці накопичується велика кількість гемул. Восени колонії бодяги гинуть, а гемули зимують. Навесні археоцити виходять назовні через особливий отвір — пору та розвиваються в нову губку.

Рис. 76. Гемула прісноводних губок — зовнішній вигляд (а), схема внутрішньої будови (б):
1 — амфідиски; 2 - пінистий шар; 3 —амебоцити
Гемули трапляються й у деяких морських губок (Suberites, Cliona, Haliclona та ін.), але вони мають простішу будову й не містять спеціальних скелетних елементів. Унікальний приклад утворення планктонних гемул відомий у деяких чотирипроменевих губок (Alectona, Thoosa), у мезоглеї яких утворюються гемули, що вкриті кремнеземними дисками та довгими головчатими голками, за допомогою яких вони линуть у товщі води.
Розмноження фрагментацією відоме для деяких морських губок, їх тіло розпадається на кілька частин, кожна .з яких започатковує новий організм. Така частина складається з групи археоцитів, вкритих оболонкою з пінакощитів. Інколи виникають лише дві такі частини (поздовжній поділ), Статеве розмноження характерне для більшості губок. Вони бувають роздільностатевими та гермафродитами. Статеві клітини (гамети) утворюються в паренхімі внаслідок редукційного поділу археоцитів або хоаноцитів. Статевих залоз (гонад) немає: кожна клітина виникає в будь-якому місці паренхіми. Перед мейозом клітини, з яких утворюються гамети (ооцити та сперматогонії), шляхом фагоцитозу поглинають інші клітини, накопичуючи запаси поживних речовин. Сперматозоїди виходять у воду, через пори проникають у тіло інших особин, мігрують у паренхіму, де запліднюють яйцеклітини. Яйце розвивається всередині материнського організму з метаморфозом: із яйця виходить планктонна розселювальна личинка, яка згодом осідає на субстрат, перетворюючись на дорослу особину.
У різних систематичних груп спостерігаються два різних типи метаморфозу. У більшості вапнякових губок запліднене яйце ділиться повністю та рівномірно, в результаті дробіння утворюється бластула, що складається з однакових джгутикових клітин. На цій стадії зародок залишає материнський організм і стає вільноплаваючою личинкою. Згодом частина клітин втрачає джгутики та мігрує всередину личинки (імміграція). Поступово порожнина бластули повністю заповнюється безладно розташованими амебоїдними клітинами. Така личинка називається паренхімулою. Через деякий час паренхімула осідає на субстрат і з неї розвивається маленька поодинока губка (рис. 77).
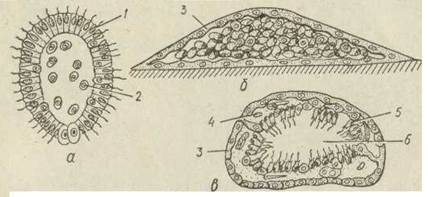
Рис. 77. Розвиток губки Clathrina blanca — паренхімула (а), личинка після осідання на субстрат (6), перетворення на дорослу губку (в):
1— кінобласт; 2 — фагодитобласт; 3 — пінакодерма; 4 — мехозіл; 5 — хоанодерма; 6 — парагастральна порожнина
У звичайних, скляних і деяких вапнякових губок яйце дробиться нерівномірно, тобто одна півкуля бластули складається з мікромерів, дрібних джгутикових клітин, а друга — з макромерів, великих безджгутикових клітин, багатих на запасні речовини (жовток). Така личинка називається амфібластулою. До субстрату вона прикріплюється полюсом з дрібними джгутиковими клітинами, й з неї розвивається молода губка, яка спочатку має асконоїдний тип будови, що згодом ускладнюється до сикону чи лейкону (рис. 78).
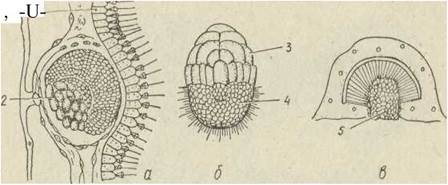
Рис. 78. Розвиток губки Sycon raphanus — зародок у тілі материнської особини (а), плаваюча амфібластула (6), прикріплення амфібластули до субстрату та початок метаморфозу (е): 1 — стінка джгутикової камери материнської особини; 2 — зародок; 3 — великі клітини; 4 — дрібні джгутикові клітини; 5— парагастральна порожнина
У багатьох невапнякових, зокрема кремнерогових, губок з яйця виходить личинка типу паренхімули, однак у неї відбувається дуже раннє диференціювання клітин: вже на стадії плаваючої личинки утворюються джгутикові камери з хоаноцитами, спікули тощо, під час осідання на субстрат зовнішній джгутиковий епітелій фагоцитується клітинами паренхіми та заміщується шаром пінакоцитів.
Таким чином, у двох типах розвитку губок переважають процеси міграції окремих клітин або вгинання шарів клітин, наприклад, у амфібластули під час осідання на субстрат джгутиковий шар вгинається всередину. В питанні про те, з якого типу клітин личинки утворюється той чи- інший шар тіла дорослої особини, серед вчених єдиної думки немає. Деякі дослідники вважають, що дрібні джгутикові клітини паренхімули та амфібластули перетворюються на хоанодерму, а великі безджгутикові клітини — на пінакодерму та клітини паренхіми. Решта стверджує, що хоанодерма та пінакодерма утворюються із джгутикових клітин, паренхіма — з безджгутикових. Отже, в обох випадках у личинок губок формуються два шари — кінобласт (джгутиковий) та фагоцитобласт (безджгутиковий), які можна порівняти з такими шарами в інших первинних багатоклітинних. Таким чином, до губок не можна застосувати фундаментальне ембріологічне поняття «зародкові листки».
Губки — один з найдавніших типів, відомий ще з докембрійських часів. Вважають, що вони виникли від ранньої фагоцители внаслідок її переходу до сидячого способу життя.
Тип Губки поділяється на чотири класи: Вапнякові губки, Скляні губки, Звичайні губки та Археоціати.
