ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 1 - Г.Й. Щербак - 1995
ПІДЦАРСТВО БАГАТОКЛІТИННІ (METAZOA)
РОЗДІЛ СПРАВЖНІ БАГАТОКЛІТИННІ (EUMETAZOA)
ТИП КИШКОВОПОРОЖНИННІ (CNIDARIA, АБО COELENTERATA)
КЛАС ГІДРОЇДНІ (HYDROZOA)
Підклас Гідроподібні (Hydroidea)
До цього підкласу належать кишковопорожнинні, переважна більшість яких утворює мономорфні колонії, прикріплені до субстрату. У деяких неколоніальних видів поліпи здатні плавати біля поверхні води. Найбільш поширені види мають життєві цикли з яскраво вираженим метагенезом.
Найпростіше побудовані поодинокі поліпи (ряд Hydrida). їх тіло має вигляд циліндричного мішка, що прикріплюється підошвою до субстрату, на протилежному кінці міститься ротовий отвір, оточений віночком із 4—12 щупалець. Рот веде в гастральну, або кишкову, порожнину.
Стінка тіла складається з епідерми та гастродерми, між якими замість мезоглеї залягає тонка базальна перетинка — безструктурна речовина, продукт виділення обох пластів (рис. 90).
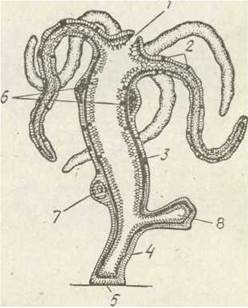
Рис. 90. Схема будови гідри:
1 — рот; 2 — щупальце; 3 — тулубний відділ; 4 —стебельце; 5 — підошва; 6— чоловічі гонади; 7 — жіноча гонада; 8 — брунька
Епідерма складається з кількох типів клітин, що виконують різні функції (рис. 91). Більшість з них — епітеліально-м'язові клітини. Тіло такої клітини входить до складу епітелію, а на своєму базальному кінці має перпендикулярний до осі клітини м'язовий відросток, що містить
одне скоротливе волокно. Всі м'язові відростки в епідермі напрямлені паралельно поздовжній осі тіла. Деякі з цих клітин мають джгутики. В епідермі є також зазолисті клітини, які виділяють клейкий секрет, особливо багато їх в області підошви. До складу епідерми входять також нервові клітину (нейрони), які поділяють на три типи: чутливі, асоціативні та рухові.
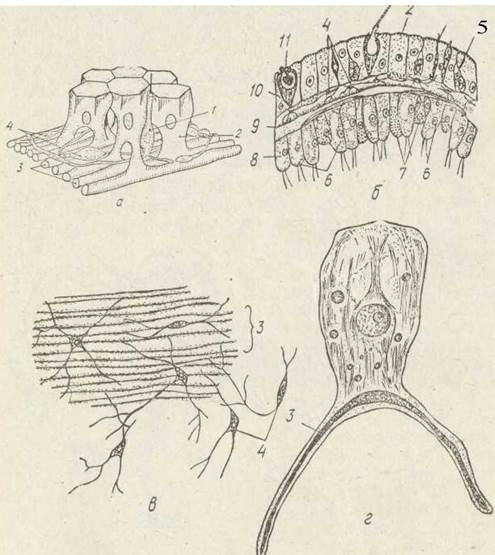
Рис 91 Типи клітин гідри — схема просторового розміщення клітин епідерми (а), розріз через стінку тіла (б), нервовий плексус (в), епітеліально-м'язова клітина (г):
1— чутлива клітина; 2 — епітеліально-м’язова клітина; 3 —скоротливі відростки епітеліально-м'язових клітин: 4 — нервові клітини; 5 — інтерстицівльні клітини; 6 —тоавні клітини; 7 — залозисті клітини; 8 — гастродерма; 9 — опорна пластина;
10 — епідерма; 11 — жалка клітина
Чутливі клітини розташовані за епітеліальними, асоціативні та рухові — під епітелієм одному рівні з м'язовими відростками. Всі нейрони мають численні відростки, що контактують між собою, а рухові нейрони — також і з м'язовими відростками. Таким чином, усі нервові клітини з'єднані в плетиво, або дифузний плексус. Він утворює скупчення клітин навколо рота й підошви, однак справжніх нервових вузлів, або гангліїв, у кишковопорожнинних не утворюється. Але навіть ця примітивна структура забезпечує пристосувальні реакції тварини. Наприклад, гідра реагує на механічні подразнення скороченням тіла, причому спочатку скорочується ділянка, що прилягає до місця подразнення, а потім, у разі тривалішого подразнення, воно поширюється на інші ділянки. Такий характер реакції властивий саме дифузійній нервовій системі. Дифузний плексус проводить збудження у всіх напрямках; поширення збудження супроводжується хвилею м'язових скорочень.
Здатність гідроїдних реагувати на механічні подразнення пов'язана з розвинутим чуттям дотику. Це виявляється, зокрема, в тому, що гідри звичайно обирають жорсткий субстрат і активно переміщуються по ньому. Спеціалізованих органів для сприймання світла в поліпів немає, однак світло приваблює гідру, подразнюючи чутливі клітини епідерми. У гідри є й специфічні хеморецептори — чутливі клітини, які реагують на поживні речовини й керують її харчовою поведінкою. Вони містяться в епідермі та ґастродермі.
У епідермі локалізовані також найбільш спеціалізовані жалкі клітини — кнідоцити (рис. 92). Вони є знаряддям нападу та захисту. В найбільш типовому випадку жалка клітина містить всередині овальну жалку капсулу, наповнену рідиною. На одному полюсі стінка капсули має впинання у вигляді порожнього, дуже тонкого відростка, який утворює закручену спіраллю жалку нитку. На зовнішній поверхні клітини розташований тонкий чутливий волосок — кнідоциль, ультраструктура якого нагадує довгий джгутик, оточений 18—22 тонкими виростами цитоплазми—мікро- ворсинками. На відміну від справжніх джгутиків кнідоциль нерухомий. У разі найлегшого дотику до кнідоциля спостерігається стрімке, подібне до пострілу, вивертання жалкої нитки. Вона встромлюється в тіло жертви й впорскує отруйний вміст капсули. Нитка вкрита напрямленими назад щипами, завдяки чому утримується в тілі жертви. Назад у капсулу жалка нитка не втягується, клітина висмикується з покриву й гине. Замість неї утворюються нові жалкі клітини. Жалкі клітини такого типу називаються пенетрантами. Інші — вольвенти — мають короткі, позбавлені шипів нитки, які обплутують здобич, у глютинантів є довгі липкі нитки, що утримують здобич. Найбільше жалких клітин міститься на щупальцях, де вони утворюють скупчення (батареї). Функціонування жалких клітин автономне й не пов'язане з нервовою системою. Є тварини (голозяброві молюски, турбелярії тощо), які, доїдаючи кишковопорожнинних, не перетравлаюють кнідоцити. Жалкі клітини мігрують до поверхні тіла і відіграють захисну роль.
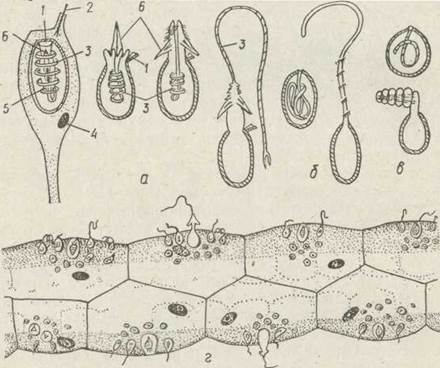
Рис. 92. Тиші жалких клітин гідри — пенетранти на різних стадіях вистрілювання жалкої нитки (а), глютинанти (б), вольвенти (б), батареї жалких клітин на щупальцях (г): 1— кришечка жалкої капсули; 2 — кнідоциль; 3—жалка нитка; 4 ядро; 5 —розширена основа жалкої нитки із щетинками, що прикриті зовні стилетами (5)
Біля основи епітеліально-м’язових клітин епідерми містяться дрібні недиференційовані клітини, що називаються інтерстиціальними (і-клітини). Вони можуть перетворюватися на інші типи клітин екто- та ентодерми, замінюючи загиблі клітини.
Епітеліально-м'язові клітини розмножуються, започатковуючи такі ж клітини, а нервові, жалкі та залозисті клітини вже настільки спеціалізовані, що втратили здатність до поділу й утворюються лише за рахунок і-клітин. Із інтерстиціальних клітин розвиваються також статеві клітини.
Гастродерма складається з епітеліально-м'язових і залозистих клітин. Перші мають таку ж будову, як і епідермальні, але їх м'язові відростки спрямовані перпендикулярно до поздовжньої осі тіла й діють як антагоністи м'язових відростків епідерми. Епітеліальні частини епітеліально-м'язових клітин мають два-п'ять джгутиків і можуть утворювати псевдоподії, тобто здатні до фагоцитозу.
Завдяки скороченню м'язових шарів поліп може видовжуватися, скорочуватися, вигинатися, його щупальця підтягують до рота здобич, яка заковтується за рахунок Координованої роботи м'язів епідерми та гастродерми.
Залозисті клітини виділяють у гастральну порожнину травний сік із ферментами, їжею гідроїдів є дрібні планктонні організми, переважно ракоподібні та їх личинки, інколи — ікра й личинки риб.
Вбита або паралізована жалкими клітинами здобич, потрапивши в гастральну порожнину, під дією травних ферментів частково перетравлюється, подрібнюється до стану маленьких часточок, які фагоцитуються епітеліально-м'язовими клітинами. Завдяки руху джгутиків вміст гастральної порожнини (вода, продукти травлення, дрібні часточки їжі) переміщується, що сприяє контакту харчових часток із клітинами фагоцитуючсго епітелію. Неперетравлені рештки викидаються з гастральної порожнини через рот, Отже, в кишковопорожнинних має місце порожнинне та внутрішньоклітинне травлення.
У складі гастродерми є також нервові клітини, які лежать поодинці або утворюють ентодермальний плексус, але більш рідкий, ніж у епідермі.
Видільної системи в кишковопорожнинних немає. Кінцеві продукти обміну речовин накопичуються в гастродермі, звідки виводяться в гастральну порожнину, а звідти через рот назовні.
Поліпи дихають усією поверхнею епідерми та гастродерми. У разі відсутності їжі в шлунку гастральна порожнина заповнюється водою, яка переміщується завдяки роботі джгутиків, що сприяє більш інтенсивному обміну.
Більшість видів морських гідроїдних поліпів (ряд tolida) утворює колонії, що складаються з величезної кількості особин. Колонії можуть бути різними за формою, найчастіше кущоподібними або деревоподібними. Вони прикріплюються до субстрату особливими відростками, які нагадують коріння. Стовбур колонії розгалужується, на кінцях гілок сидять окремі особини, схожі на гідру, які звуться гідрантами. Загальний стовбур і розгалуження, які територіальне не належать до жодного поліпа, називаються ценосарком (рис. 93).
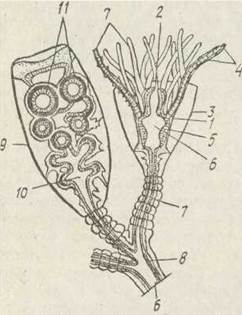
Рис. 93. Будова ділянки колонії Obelia geniculata:
1 — епідерма; 2 — рот; 3 —гідротека; 4 — щупальця; 5 — гастродерма; 6 — гастральна порожнина; 7 — тека; 8 — стовбур колонії (ценосарк); 9 — гоноска (ділянка теки, ідо вкриває благостиль); 10 — бластостиль; 11 — медузи. що відбруньковуються
Навколо ценосарка утворюється зовнішня органічна оболонка з хітиноподібної речовини — тека, що виділяється клітинами епідерми й виконує захисну та опорну функції. У деяких груп тека просякається вапном. Тека в деяких видів досягає основи гідрантів (підряд Athecata), у інших вкриває їх тіло, утворюючи захисну чашечку, або гідротеку, навколо кожного гідранта (підряд Thecaphora). У деяких видів колонії можуть досягати великих розмірів, при цьому окремі гідранти залишаються мікроскопічними.
Гідранти мають таку ж гістологічну будову, як і гідра, але їх гастральні порожнини продовжуються в загальний стовбур колонії й таким чином гастральні порожнини, всіх гідрантів сполучаються між собою. Внутрішній канал ценосарка заповнений рідиною — гідроплазмою. Це — своєрідна розподільна система колонії.
Рідина весь час рухається, і за її допомогою відбуваються перенесення та розподіл між окремими частинами колонії часточок їжі, поживних речовин і навіть окремих клітин. У останні роки вчені навчилися утримувати колонії гідроїдів у лабораторних умовах і спостерігати за їх життєдіяльністю. Завдяки цьому виявлено, що в різних видів рух гідроплазми відбувається двома різними способами. Перший спосіб — це рух. за рахунок роботи джгутиків клітин гастродерми (наприклад, у Tubularia). При цьому порожнина ценосарка поділяється поздовжньою перетинкою (мезентерієм) на два паралельних канали, якими гідроплазма переміщується з постійною швидкістю в двох протилежних напрямках.
У інших гідроїдів порожнина ценосарка позбавлена мезентерію, й гідроплазма рухається в ній почергово то в одному, то в протилежному напрямку завдяки пульсації (скороченню та розслабленню) верхівок росту (молоді бруньки), гідрантів і ценосарка колонії. Робота розподільної системи забезпечує фізіологічну інтеграцію колонії.
Отже, колонія гідроїдів — це не просто сукупність фізіологічне самостійних індивідів, а інтегрована в єдине ціле індивідуальність вищого порядку, її розподільна система забезпечує не рівномірний розподіл їжі по всій колонії, а її концентрацію в певних, найбільш важливих для існування колонії місцях. Наприклад, якщо в лабораторних умовах годувати лише окремі гідранти, їжа буде транспортуватися в місця утворення молодих гідрантів незалежно від того, де знаходяться поліпи, що одержали їжу.
Гідроїдні розмножуються нестатевим і статевим способами. Нестатеве розмноження найчастіше відбувається шляхом брунькування, рідше — поперечного або поздовжнього поділу, а також лацерації, коли від поліпа (найчастіше від його базальної частини) відокремлюється шматочок або група клітин, з якої розвивається нова особина. У процесі брунькування поодиноких поліпів на тілі утворюється горбок, у який заходить гастральна порожнина, а стінки є продовженням стінок тіла (епідерми та гастродерми). Утворюється брунька, яка росте, набуває циліндричної форми, на вільному її кінці виростають щупальця, проривається ротовий отвір. Брунька ще деякий час утримується на материнській особині, нарешті відривається й переходить до самостійного життя, прикріплюючись до субстрату. Інколи на одній гідрі тримаються кілька дочірніх особин, утворюючи тимчасову колонію.
У колоніальних гідроїдних особини, що утворилися шляхом брунькування, не відриваються від материнської особини, а продовжують жити на її тілі, розмножуючись брунькуванням. Так утворюється колонія, розміри якої збільшуються шляхом появи нових особин.
Статеве розмноження переважної більшості гідроїдних здійснюється спеціальними статевими особинами — медузами. Вони утворюються на колонії поліпів шляхом брунькування на особливих видозмінених безщупальцевих поліпах — бластостилях (див. 93), відриваються від них і переходять до вільноплаваючого життя.
Медуза за будовою відрізняється від поліпа. Вона має вигляд дзвона або парасольки. Зовнішній опуклий її, бік називається ексумбрелою, внутрішній, увігнутий — субумбрелою. По внутрішньому краю дзвона проходить подвійна складка епідерми, що має форму кільця — парус. Це — характерна риса гідроїдних медуз, яка відрізняє їх від сцифомедуз. Парус, скорочуючись, зменшує діаметр вустя дзвона, й медуза рухається за реактивним принципом, виштовхуючи воду з-під дзвона (рис. 94).
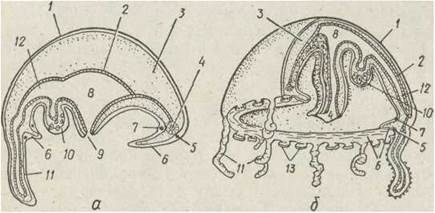
Рис. 94. Схема будови гідроїдної медузи — поздовжній розріз (а), медуза з відрізаною чвертю дзвона (б): 1 — епідерма; 2 — гастродерма, 3 — мезоглея; 4 — кільцевий канал; 5 — зовнішнє нервове кільце: 6 — парус; 7 — внутрішнє нервове кільце; 8 — шлунок: 9 — рот; 10 — гонада; 11 — щупальце; 12 — радіальний канал; 13 — статоцисти
По краю дзвона розташовані щупальця, кількість яких найчастіше кратна чотирьом. У центрі субумбрели розташоване довге ротове стебельце, на кінці якого міститься ротовий отвір. Рот веде в гастральну порожнину, вистелену гастродермою. На відміну від поліпа гастральна порожнина медузи має більш складну будову. Вона складається з невеликого центрального шлунка й радіальних каналів,
які розходяться від нього до країв дзвона та об'єднуються, там у кільцевий канал. Це — гастроваскулярна система — все, що залишається від гастральної порожнини. Решта гастродерми між каналами зростається й утворює суцільну подвійну гастродермальну пластинку. Медузи — хижаки, живляться дрібними планктонними організмами.
У медузи на відміну від поліпа між епідермою та гастродермою замість тонкої базальної перетинки міститься товстий шар мезоглеї. Мезоглея бере участь у русі медузи: під час скорочення паруса дзвін звужується, а його розширення відбувається завдяки пружності мезоглеї.
За рахунок вільноплаваючого способу життя нервова система гідромедуз дещо складніша, ніж у поліпів. Крім дифузного плексуса, в медузи на краю дзвона міститься подвійне нервове кільце (внутрішнє та зовнішнє), яке. складається з нервових клітин та їх відростків. Нервове кільце інервує м'язові волокна паруса, а також органи
чуття, розташовані поблизу. У медуз є органи рівноваги — статоцисти та органи зору — вічка, яких немає в поліпів (рис. 95).
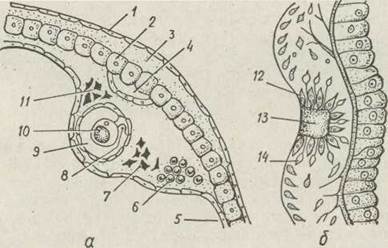
Рис. 95. Органи чуття гідроїдних медуз — поздовжній розріз через- статоцист, що розташований при основі щупальця медузи Оbеlіа (а), вічко гідроїдної медузи (б):
1 — епідерма ексумбрели; 2 — гастродерма; 3 — мезоглея; 4—кільцевий канал; 5 — щупальце; 6 — інтерстиціальні клітини;
7 — зовнішнє нервове кільце; 8—чутливий відросток статоциста; 9 — статцист; 10 — статоліт; 11 — внутрішнє нервове кільце; 12— пігментна клітина; 13 — кришталик; 14 — чутлива клітина
Статоцисти найчастіше мають вигляд ямки або міхурця, всередині якого містяться кристалики карбонату кальцію— статоліти, а стінки вкриті нервовими клітинами, що мають чутливі волоски. Ці клітини відростками зв'язані з нервовим кільцем. Зміна в положенні тіла медузи приводить до подразнення статолітами відповідних чутливих волосків, збудження передається в нервове кільце, що спричинює скорочення паруса. Статоцисти забезпечують також ритмічність роботи м'язів паруса. Під час коливань води перед штормом статоліти вібрують, що є сигналом для занурення медуз у глибинні шари води.
Очі гідромедуз мають різну будову. Вони розташовані біля основи щупалець у вигляді очних плям або очних ямок. До складу ока входять світлочутливі та пігментні клітини. Світлове подразнення сприймається світлочутливими клітинами та передається їх відростками у нервове кільце. Завдяки наявності пігментних клітин світло потрапляє на чутливі клітини тільки з одного боку. В очах більш складної будови світлочутливі та пігментні клітини лежать на дні невеликого впинання — ямки, порожнина якої заповнена прозорим виділенням епідерми — кришталиком, що концентрує світлові промені на чутливих клітинах. Незважаючи на складність будови вічок, медуза не може сприймати форму предмета, а реагує лише на інтенсивність освітлення. За рахунок цього гідромедузи можуть здійснювати вертикальні міграції —від освітлених верхніх шарів води в глибину й навпаки.
Переважна більшість гідроїдних медуз дуже дрібні, найчастіше їх діаметр становить 1—2 мм, рідше — кілька сантиметрів. Вони є хижаками, живляться найдрібнішими планктонними організмами — рачками, личинками різних безхребетних.
Медузи розмножуються за деякими винятками лише статевим шляхом; вони роздільностатеві. Статеві органи (гонади — це групи статевих клітин, які містяться між епідермою та мезоглеєю на нижньому боці дзвона під радіальними каналами або на ротовому стебельці. Статеві клітини — яйцеклітини та сперматозоїди — утворюються з клітин ектодерми.
Статеві клітини (гамети) після дозрівання виходять назовні крізь розрив стінки тіла. У воді сперматозоїди запліднюють яйцеклітини. Запліднена яйцеклітина проходить повне та рівномірне дробіння, яке в кишковопорожнинних ще не має сталої характеристики; воно дуже мінливе навіть у одного виду. В результаті дробіння утворюється суцільна багатоклітинна кулька — морула або кулька з одним поверхневим шаром і порожниною всередині — бластула. Гаструляція відбувається шляхом деламінації (у морули) або імміграції (у бластули). Внаслідок цього утворюється личинка паренхімула, поверхневий шар клітин якої, має джгутики та є ектодермою, а внутрішня аморфна маса клітин — ентодермою (див. рис. 89).
Паренхімула, яка нагадує личинку губок, виходить з яйця у воду й плаває за допомогою джгутиків. Пізніше частина ентодермальних клітин личинки руйнується й на їхньому місці виникає порожнина. На цій стадії личинка називається планулою. Планула деякий час (від кількох годин до двох діб) плаває, потім прикріплюється до субстрату переднім кінцем. На цьому місці утворюється підошва, на протилежному кінці проривається ротовий отвір, навколо нього виростають щупальця, й таким чином утворюється поліп. Ектодерма личинки диференціюється та перетворюється на епідерму поліпа, ентодерма утворює відповідно гастродерму. У поодиноких видів поліп виростає в дорослу особину й починає розмножуватися брунькуванням. У колоніальних видів шляхом брунькування з первинного поліпа утворюється колонія.
Отже, життя гідроїдних поліпів складається з правильного чергування двох поколінь, що відрізняються за будовою та способом розмноження (рис. 96). Одне покоління — поліпоїдне — веде сидячий спосіб життя, розмножується лише нестатевим способом, утворюючи поліпи та медузи. Друге покоління — медузоїдне — утворюється на колонії поліпів, веде вільнорухливе життя й розмножується статевим шляхом. Оскільки таке правильне чергування нестатевого та статевого поколінь називається метагенезом, медузи, які утворюються на колоніях поліпів, є метагенетичними. Медузи за рахунок активного руху, а також пасивно, за допомогою течій, розносяться на велику відстань від сидячої колонії й виконують функцію розселення виду. В цьому полягає біологічне значення стадії медузи.
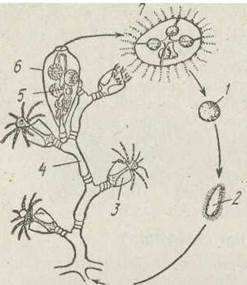
Рис. 96. Життєвий цикл Obelia:
1 — яйця; 2 —планула; 3 — гілрант; 4 — ценосарк; 5 — бластостиль; 6 — гонотека; 7 — медуза
Описаний цикл розвитку є типовим, проте в деяких гідроїдних має місце пригнічення одного з поколінь — медузоїдного або поліпоїдного — гіногенез. У разі гіногенезу медузи, які утворюються на колонії, не відриваються від неї, а залишаються жити й розмножуватися, перебуваючи в прикріпленому стані. У таких медуз недорозвинені щупальця, рот і органи чуття. Вони називаються медудоїдами. В інших видів редукція медуз продовжується й може доходити до того, що вони стають схожими на мішечки із статевими клітинами. Такі утвори називаються гонофорами, або споросаками (рис. 97). У кінці цього ряду можна поставити гідр (ряд Hydrida), в яких від медузи не залишається нічого, крім гонад. На тілі гідри під епідермою утворюються чоловічі та жіночі гонади (див. рис. 90). Більшість гідр — гермафродити. Сперматозоїди крізь розрив стінки тіла виходять у воду, проникають у іншу особину й запліднюють її яйцеклітини.
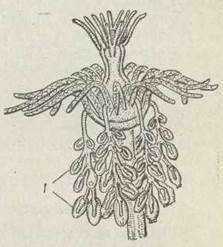
Рис. 97. Tubitlaria larynx — окремий гідрант з гонофорами (1)
Із заплідненої яйцеклітини розвиваються молоді гідри. Отже, гідра це поліп, що втратив медузоїдне покоління та став розмножуватися нестатевим і статевим шляхами. Разом із цим гідра втратила й метаморфоз — вона не має вільноплаваючої планктонної личинки. Імовірно, що це — наслідок переходу її предків до мешкання в прісних водоймах. Інший вияв гіногенезу — повна втрата поліпоїдного покоління. Є група видів (ряд Trachilida), у яких існує лише покоління медуз. Вони розмножуються статевим способом, а з запліднених яєць утворюються планули, які відразу ж. перетворюються на медуз. До підкласу Гідроподібні (Hydroidea) належить кілька рядів.
Ряд Лептоліди (Leptolida)
Це переважно морські форми, іноді трапляються й прісноводні. Найчастіше вони є колоніальними організмами. Колонії мають кущоподібну або деревоподібну форму. Для них характерний різною мірою виражений метагенез. Це найбільш численна група гідроїдних. У півічноєвропейських морях (Баренцовому, Білому) найпоширенішими є Obelia geniculata, Tubularia, Coryne. У арктичних та помірних водах північної півкулі, зокрема в Чорному морі, часто трапляється вид Rathkea octopunctata. В опріснених водах оселяється Bougainvillia megas — масовий вид, поліпи якого беруть участь у обростанні суден і портових споруд. Поширена в морях обох півкуль Obelia longissima. Жителям Приморського краю, Курильських островів та Японії добре відомі невеличкі медузки (діаметром 25 мм) Gonionemus, дотик щупалець яких викликає у людини відчуття сильного опіку, після чого утруднюється дихання та виникає загальна слабкість. Людина, яка перебуває у воді далеко від берега, може потонути (рис. 98).
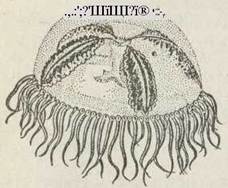
Рис. 98. Ряд Leptolida: Gonionenius
Це морські плаваючі, яскраво забарвлені поодинокі поліпи, що живуть на поверхні води тропічних морів, їх тіло сплющене, рот і щупальця спрямовані донизу. На верхньому боці особин утворюється хітиноїдна пластинка, яка виступає з води й є парусом. На нижньому боці утворюються гонофори, на яких відбруньковуються медузи (рис. 99).
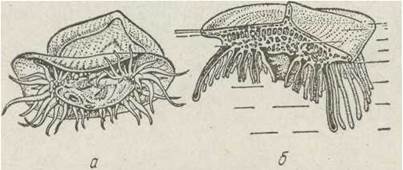
Рис. 99. Ряд Chondrophora — Velella:
а — молода особина, що захопила малька риби; б — схема будови
Існує й інша думка щодо природи цих організмів. Деякі вчені вважають, що це — колонія, яка складається з одного великого поліпа та кількох недорозвинених, прикріплених знизу поліпів. Найбільш відомий представник, ряду — парусник (Velella). Парусники живуть у субтропічних водах Світового океану. Ці досить великі поліпи (довжина паруса — до 12 см) можуть збиратися у величезні зграї, що тягнуться на десятки кілометрів.
Ряд Пдрокорали (Stylasterida)
Представники ряду є колоніальні гідроїдні поліпи, зовнішня оболонка яких (тека) має вигляд добре розвиненого вапнякового скелета. Медузоїдне покоління в них редуковане, медузи не відриваються від колонії. Гідрокорали, зокрема фіолетова дістіхопора (Distichopora violacea), беруть участь у побудові рифів.
Ряд Трахімедузи (Trachilida)
До цього ряду належать винятково морські гідроїди, що мають форму медузи. Поліпів у них немає. По всьому Світовому океану трапляються медузи роду Aglantha (рис. 100). На медузах ряду Leptolida паразитують види роду Cunina.
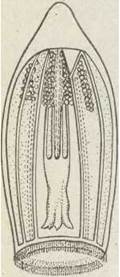
Рис. 100. Ряд Тrachilidar Aglantha
Це переважно прісноводні, рідше морські поодинокі поліпи, які не мають медузоїдного покоління. Найпоширенішою в прісних водах України є Hydra oligactis.
Більшість гідроїдів мешкає на невеликих глибинах — від літоралі до 200—250 м. Вони прикріплюються до кам'янистого грунту, різних підводних предметів, вкриваючи морське дно густими заростями. Гідроїди не бояться прибою, тому що їх колонії захищені текою, гілки гнучкі й вигинаються під дією хвиль, не ламаючись.
Зарості гідроїдних поліпів разом із бурими водоростями є особливим угрупованням, яке заселяють черви, молюски, ракоподібні, голкошкірі, що знаходять тут притулок та їжу. Вони оселяються на підводних частинах суден, вкриваючи їх суцільним волохатим шаром. Цим гідроїди завдають значної шкоди судноплавству, знижуючи швидкість суден. Вони можуть оселятися на різних спорудах і в трубах морського водопроводу, закриваючи його просвіт і перешкоджаючи надходженню води. Гідрокорали разом із справжніми коралами беруть участь в утворенні рифів.
Гідроїди є джерелом біологічно активних речовин — простагландинів, які використовуються в сучасній медицині. З гідроїдів Obelia виділено препарат обелін — індикатор фізіологічного стану клітин, що використовується для ранньої діагностики різних захворювань людини.
Гідроїдні медузи, незважаючи на малі розміри, дуже зажерливі. Вони поїдають безліч рачків і тому вважаються конкурентами планктоноїдних риб.
У прісних водоймах мешкає лише близько двох десятків видів кишковопорожнинних. Усі вони належать до підкласу Hydroidea. Серед них є кілька видів гідр ряду Hydrida. Перший дослідник гідр — французький вчений Трамбле ще в середині XVIII ст. дослідив будову гідри, її поведінку, розмноження та регенерацію. Розрізаючи гідру вздовж або впоперек, він спостерігав відновлення цілої тварини з окремих шматочків. Багаторазово оперуючи одного з піддослідних поліпів, Трамбле одержав «семиголового» поліпа. Відрізавши всі його «голови», дослідник спостерігав їх відновлення подібно до того, як відростали голови в міфічної потвори — Лернейської гідри.
Гідра — ненажерливий хижак. Вона живиться інфузоріями, планктонними ракоподібними, малощетинковими червами, нападає на мальків риб. Гідри — надзвичайно зручний об'єкт для різноманітних досліджень — фізіологічних, біохімічних, генетичних тощо.
Крім гідр, у прісноводних акваріумах трапляється прісноводна медуза краспедакуста (Craspedacusta, ряд Leptotida) — невеличка (діаметр близько 2 см) прозора медуза, по краю якої розташовані широкий парус та близько 100 тонких щупалець. Тривалий час було невідомо, як ці медузи потрапляють у акваріуми. Виявилося, що краспедакуста має досить складний і цікавий життєвий цикл. З яйця формується червоподібна личинка, що деякий час повзає по субстрату, прикріплюється до нього та перетворюється на малесенького безщупальцевого поліпа, який нагадує білу краплину на зелених водних рослинах. Поліп може розмножуватися брунькуванням. У разі підвищення температури на поліпах з'являються медузоїдні бруньки й відокремлюються медузки. Іноді з яєць формуються також щупальцеві поліпи, але на них медузоїдне покоління не розвивається (рис. 101). Вважають, що цю медузу занесено до Європи з тропічними рослинами та акваріумними рибками.
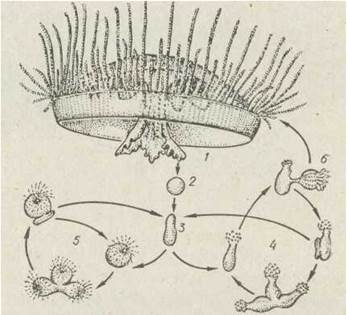
Рис. 101. Життєвий цикл краспедакуств:
1 — медуза: 2 — яйце; 3 — гтланула; 4 — безщупальцеві поліпи; 5 — щупальцеві поліпи; б—відбруньковування медузи
Серед кишковопорожнинних трапляється дуже мало паразитів. Відомо кілька видів гідроїдних медуз (ряд bectolida), які паразитують на інших видах медуз. Але найцікавішим прикладом паразитизму є Polypodium hydriforme—-паразит ікри осетрових риб (рис. 102). Поліпи живуть усередині ікринок, живлячись їх жовтком, ікринки при цьому не розвиваються та втрачають поживні якості. В одній інкринці утворюється нерозгалужений стовбур — столон — із 20—60 поліпами. Цікаво, що поліпи всередині ікри вивернуті назовні, тобто зверху в них міститься гастродерма, а всередині — епідерма. Це пов’язано з тим, що поживні речовини вони одержують іззовні — з жовтка ікри, що їх оточує. Перед виходом із ікри у воду вони вивертаються, й шари їх тіла набувають нормального положення.
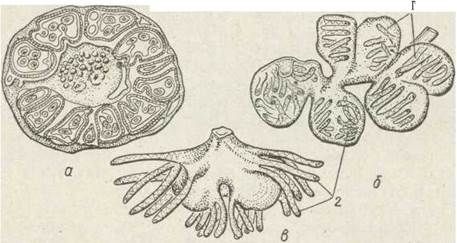
Рис. 102. Polypodium hydriforme - паразит всередині ікри (а), . фрагмент столону (б), медузоподібна стадія (б):
1 — окремий поліп; 2 — щупальця
У воді розвиваються вільноживучі стадії — столон із поліпами швидко розпадається на фрагменти з кількох особин, а потім — на поодинокі особини, медузоподібні організми з редукованим дзвоном і органами чуття. Вони малорухливі, тримаються біля дна, в них розвиваються гонади. Шляхи враження риб паразитами остаточно ще не з'ясовані, але відомо, що ембріональний розвиток і формування личинки відбувається вже в організмі риби й триває кілька років. Розвиток паразитичних стадій Р. hydriforme узгоджений із розвитком і розмноженням його хазяїна — осетрової риби.
Систематичне положення Р. hydriforme остаточно не встановлене. Обговорюється необхідність виділення для цього виду нового класу—Polypodiozoa.