ЗООЛОГІЯ БЕЗХРЕБЕТНИХ У ТРЬОХ КНИГАХ - КНИГА 2 - Г.Й. Щербак - 1996
ТИП КІЛЬЧАСТІ ЧЕРВИ (ANNELIDA)
КЛАС МАЛОЩЕТИНКОВІ (OLIGOCHAETA)
Малощетинкові живуть у прісних водоймах, ґрунті, рідше (близько 200 видів) у морях. Більшість олігохет — риючі форми, деякі живуть на поверхні дна серед решток, що гниють; невелика кількість видів паразитує на зябрах ракоподібних. Відомо близько 5000 видів. У прісних водоймах України виявлено близько 200 видів олігохет, у фауні Чорного та Азовського морів — 33 види. Ґрунтові олігохети тут вивчено недостатньо, відомо близько 35 видів дощових червів. Більшість малощетинкових має розміри від 0,5 мм до 40 см, а деякі види тропічних земляних червів (Megascolides australis) досягають 3 м.
За загальним планом будови олігохети близькі до Polychaeta. Це виключно полімерні кільчаки, що втратили параподії та більшість придатків простомію. Вони мають гомономну метамерію, проте на певних сегментах тіла у статевозрілих особин є кільцеподібне розширення — поясок, епідерміс якого виділяє речовину кокона. Гермафродити, статеві органи розташовані лише у певних сегментах тіла. Розвиток прямий, без метаморфозу.
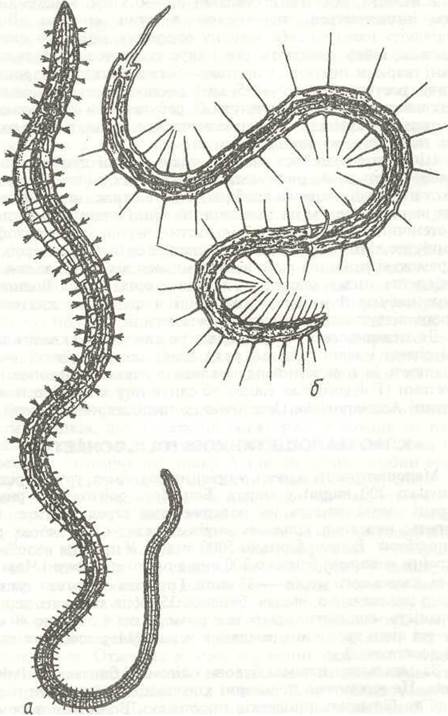
Рис. 19. Олігохети:
а - трубочник Tubifex tubifex (родина Tubificidae); б - Stylana lacustris (родина Naididae)
Тіло олігохет більш-менш витягнуте, майже циліндричне (рис. 19). Простомій, як правило, не має придатків та очей, і лише в деяких морських видів є парні очі (родина Naididae). Зрідка простомій буває витягнутий у хоботок. Перистомій відсутній, рот розташований на першому сегменті тулуба (рис. 20). Кількість сегментів коливається від 5—6 до 500— 600. Параподії редуковані, на їх місці залишаються парні щетинки. У кожному сегменті, крім першого, є, як правило, чотири пучечки щетинок —два спинні та два черевні. Іноді спинні пучечки редуковані, або, рідше, можуть взагалі зникати.
У кожному пучечку, як звичайно, є пара щетинок, а у водних форм — від 2 до 10—15, причому різноманітної форми (голкоподібні, гачкоподібні, пірчасті тощо). У деяких тропічних форм (рід Pheretimania) щетинок буває до 150, вони утворюють віночки навколо кожного сегмента. Щетинки беруть участь у рухах червів: риючі форми впираються ними у стінки нірки, водяні за їх допомогою плавають. На статевих сегментах є особливі щетинки, розміщені поблизу від чоловічих статевих отворів. Вони розширюють отвори сім'яприймачів партнера при копуляції. Крім того, біля отворів сім'яприймачів є особливі гострі й товсті ножеподібні щетинки, які мають поздовжній жолобок. Вони з силою впираються в шкіру партнера і, ймовірно, спрямовують сперму до сім'яприймачів.
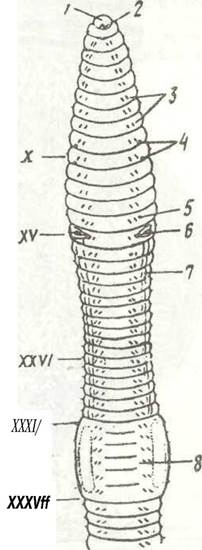
Рис. 20. Передня ділянка тіла дощовика Lumbricus terrestris із черевної сторони:
1 - простомій; 2 - рот; 3 - черевні щетинки; 4 - отвори сім'яприймачів; 5 - жіночий статевий отвір; 6 - чоловічий статевий отвір; 7 - сім'яна борозенка; ,8 - поясок; 9 - X - XXXVII - сегменти
Шкірно-м'язовий мішок (рис. 21) має характерну для кільчаків будову й складається з тонкої еластичної кутикули, шкірного епітелію (епідермісе) та двох шарів м'язів: тонкого кільцевого й товщого поздовжнього. Між мускульними волокнами рихло лежать клітини сполучної тканини. У місцях відходу щетинок утворюються впинання кутикули та епідермісу — щетинконосні мішечки. Шкіра олігохет багата на залозисті клітини, що лежать між епітеліальними. Слиз, який вони виділяють, змащує тіло, в ґрунтових олігохет робить його вологим, що є необхідною умовою для шкірного дихання. Численні слизові та білкові одноклітинні залози розташовані в епітелії пояска; вони виділяють речовину кокона.
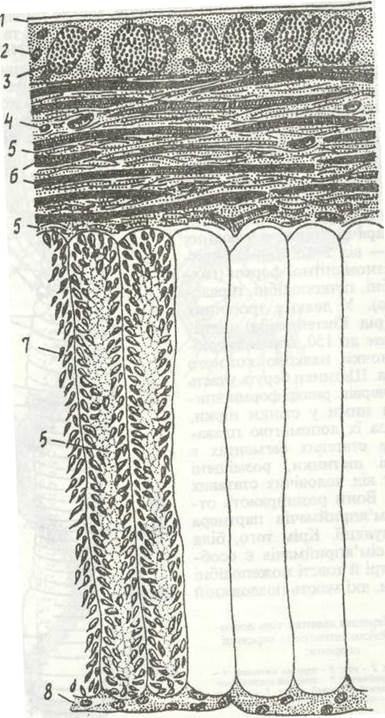
Рис. 21. Будова шкірно-м'язового мішка Lumbiicus terrestris:
1 кутикула; 2 - епідерміс; 3 - слизова залоза; 4 - ядро сполучнотканинної клітини; 5 - сполучна тканина; 6- кільцеві м'язові волокна; 7 - поздовжні м'язові волокна; 8 - перитонеальний епітелій
Під шкірно-м’язовим мішком міститься шар перитонеального епітелію, що щільно прилягає до нього і вистилає вторинну порожнину тіла — целом (рис.22).
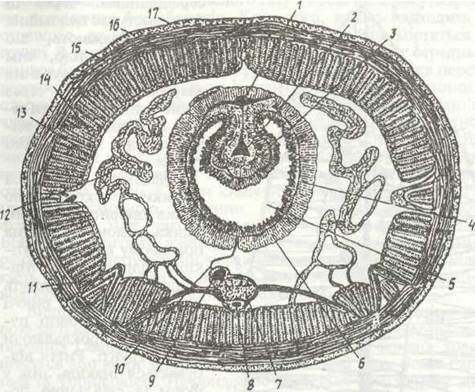
Рис. 22. Схема поперечного зрізу через середню частину тіла Lumbricus terrestris:
1- спинка кровоносна судннв; 2- судини тнфлоіоля; 3 - тифлозоль. 4 - кишковий й судинний плексус; 3 - середня кишка, б - споратогенні клітини; 7 черевний нервовий ланшожок; 8 - субневральна судина; 9 - черевна кровоносна судина; 10 - мезентерій; 11 - щетинконосний мішок; 12 - метапефридін; 13 - целомічниіі епітелій: 14 - поздовжня мускулатура; 15 - кільцева мускулатура; 16 - епідерміс; 17 — кутикула
На середніх та задніх сегментах багатьох ґрунтових видів (Lumbricus та ін.) поблизу від міжсегментних боріздок е непарні дорзальні пори, що зв'язують целом із зовнішнім середовищем. Через ці пори виступає целомічна ріцина. Вважають, що вона змащує тіло, чим полегшує пересування тварини в грунті. Через пори задньої частини тіла з порожнини тіла виділяються екскреторні тільця.
Олігохети рухаються завдяки скороченню м'язів шкірно-м'язового мішка, целомічна рідина при цьому виконує функцію гідроскелета. Малощетинкові можуть повзати, плавати; риючі форми безперервно рухаються, прокладаючи ходи в ґрунті. Риття відбувається інакше, ніж у форм із суцільною порожниною тіла (наприклад, приапулід). Септи між сегментами не дають змоги рідині перекачуватись через усе тіло, вона рухається лише в межах 2—5 сегментів завдяки отворам у дисепиментах.
Травна система починається ротовим отвором, що веде в мускулясту глотку, куди відкриваються численні слинні залози. Глотка переходить у вузький стравохід, який на задньому кінці може розширюватись у воло, а далі йде м'язистий шлунок (рис. 23). Ці відділи належать до ектодермальної передньої кишки. У видів, котрі пропускають через кишечник ґрунт, є три пари вапнякових залоз, які відкриваються послідовно одна в одну та парою отворів — у стравохід. Вони забиті кристаликами вуглекислого кальцію, їхня основна функція — вилучення з крові карбонатів, а також зв'язування надлишку вуглекислого газу в крові шляхом утворення розчинних бікарбонатів. Крім того, вапно надходить у стравохід, де нейтралізує гумінові кислоти, що містяться в ґрунті й шкідливі для органів травлення. Стінки середньої кишки багаті на війчасті та секреторні клітини. Тут їжа перетравлюється та всмоктується. У ґрунтових олігохет на спинній стороні середньої кишки є поздовжнє жолобоподібне вип'ячування в кишкову порожнину — тифлозоль, за рахунок якого значно збільшується поверхня кишки (рис. 22, 23). Він закінчується поблизу заднього кінця тіла, де середня кишка переходить у коротеньку задню, що відкривається анальним отвором на пігідії. Стінки кишечника зовні вкриті хлорагогенною тканиною, клітини якої виконують видільну функцію: в них нагромаджуються продукти дисиміляції. Більшість олігохет живиться рослинними та тваринними рештками, пропускаючи через свій кишечник велику кількість ґрунту. Мешканці водойм живляться мікроскопічними водоростями, найпростішими, бактеріями.
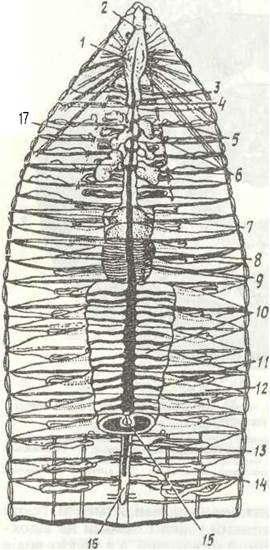
Рис. 23. Внутрішня будова Lumbricus terrestris (розтин зі спинної сторони):
1 - глотка; 2 - надглотковий ганглій; 3 - стравохід; 4 - серця; 5 - сім'яприймачі; 6 - сім'яні мішки; 7 - воло; 8 - м'язистий шлунок; 9 - спинна кровоносна судина; 20 - середня кишка; 11 - поперечні судини; 12 - дисепимент, ІЗ - черевна судина; 14 - метанефритай; 15 - тифлозоль; 16 - черевний нервовий ланцюжок; 17- радіальні м’язи глотки
Видільна система, як звичайно, представлена однією парою метанефридіїв у кожному сегменті, за винятком кількох передніх (ларвальних). Іноді кількість метанефридіїв зменшується або вони зовсім зникають (морські види родів Рагаnais та Tubificoides). Нефридій починається в сегменті з війчастої воронки. Від неї відходить тоненький канал із війками, що проходить через дисепимент у наступний сегмент. Тут нефридій утворює три лопаті, які складаються з петель каналу, оточених екскреторними клітинами целомічного епітелію. Лопаті густо обплетені кровоносними капілярами. Продукти обміну дифундують із крові в порожнину каналу. Сеча збирається в кінцевому розширенні каналу — сечовому міхурці, який відкривається назовні видільним отвором. У деяких олігохет нефридії, об'єднуючись, утворюють парний видільний канал, що впадає в задню кишку.
Крім нефридіїв, у виділенні беруть участь так звані хлорагогенні клітини, про які вже згадувалось вище. Це спеціалізовані видовжені клітини целомічного епітелію, розташовані на поверхні кишки та судин. За новими даними, їхня функція набагато складніша: в них нагромаджуються, крім продуктів обміну, запасні поживні речовини (глікоген та ліпіди).
Кровоносна система в олігохет, як і в інших кільчаків, замкнена. Судини залягають під целомічним епітелієм кишечника, дисепиментів, стінок тіла; капіляри проростають у товщу тканин. Кровоносна система складається з кишкового синуса, що може замінюватися кишковим сплетінням судин, спинної, черевної, кільцевих та поперечних судин, які не утворюють повних кілець. Кільцеві судини, що оточують стравохід, мають товсті м'язові стінки й виконують функцію сердець, що женуть кров із спинної судини до черевної. У різних олігохет їх кількість різна; Lumbricus terrestris має п'ять пар сердець (рис. 23). Крім того, у зв'язку зі шкірним диханням розвивається густа підшкірна сітка дрібних судин та капілярів. Це дуже тонкі судини, які проростають у товщу шкірно-м'язового мішка. Особливо багато їх у кільцевому шарі м'язів; найтонші їх гілочки проникають навіть у товщу епідерміса (рис. 24, а). У багатьох олігохет кров безбарвна; у частини вона має червоний колір завдяки наявності близького до гемоглобіну пігменту гемокруорину, розчиненого в плазмі. Зрідка (у деяких дощових червів) пігмент міститься всередині спеціальних клітин (гемоцитів). У тропічних велетенських дощових червів (рід Megascolides) кров зеленкувата завдяки присутності іншого пігменту — хлорокруорину. У крові є також багато безбарвних клітин різної будови. Стінки кровоносних судин утворені неклітинною опорною пластин кою, зовні вони вкриті хлорагогенними клітинами. Великі судини (спинна, кільцеві) мають у стінках м'язи (рис. 24, б).

Рис. 24. Кровоносні судини Т.bricus terrestris:
а - шкірні капіляри; б - поперечини зріз через спинну судину; 1 - поздовжня мускулатура; 2 - кільцева мускулатура; З - епідерміс; 4 - розгалуження капілярів; 5 - опорна пластинка; 6 - хлоратогенні клітини
Дихальна система, як правило, відсутня: газообмін відбувається через тоненьку вологу шкіру та кровоносні капіляри. Проте деякі водяні форми мають зовнішні зябра. Так, у червів, що переднім кінцем закопуються в мул (роди Derp та Auiophoras), хвостовий відділ має вигляд широкої лопаті, на якій розташовані парні листоподібні зябра. Поверхня зябер укрита війками, що спричиняють рух води; в зябрах багато кровоносних капілярів. У донних видів (наприклад, із родин Tubificidae та Naididae) до шкірного дихання приєднується кишкове.
Нервова система має типову для кільчаків будову і складається з парного надглоткового ганглію, навкологлоткових конектив та черевного нервового ланцюжка (рис. 25). Лише у деяких видів черевні стовбури широко розставлені, а ганглії з’єднані довгими комісура- ми (наприклад, в Аеrоsoma).
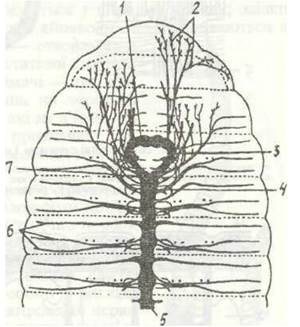
Рис. 25. Нервова система Lumbricus tenrestris:
1 - надглотковий ганглій; 2 - нерви простомія і І сегмента; 3 - навкологлоткова конектива; 4 - сегментарні ганглії;
5 - черевний нервовий ланцюжок; 6 - сегментарні нерви; 7 - підглотковий ганглій
Органи чуття у малощетинкових розвинеш слабо. Очі в основному відсутні, проте дощовики виявляють чутливість до світла. У їхньому шкірному
епітелії розкидані поодинокі клітини або групи чутливих клітин — сенсили. Вони бувають двох типів: чутливі бруньки й світлочутливі сенсили (рис. 26). Чутливі бруньки залягають у шкірному епітелії всього тіла, але особливо багато їх на простомії. Це групи чутливих клітин, що лежать у шарі епідермісу. Зовнішній кінець кожної клітини має коротеньку чутливу паличку, яка проходить через кутикулу й підіймається над її поверхнею, а базальні кінці мають відростки, які разом утворюють нервове волокно. Чутливі бруньки є рецепторами дотику й хімічного чуття. Світлочутливі клітини поодиноко розсіяні в епідермісі або утворюють групи — сенсили на тонких розгалуженнях шкірних нервів, їх також найбільше на простомії. Кожна клітина сенсили зв'язана з чутливим нервовим волокном.
Усі олігохети — гермафродити, їхня статева система розташована в небагатьох сегментах і в представників різних родин —у різних місцях тулуба. Статевий апарат складається з гонад, чоловічих і жіночих статевих проток (целомодукгів), сім'яних та яйцевих мітків, сім'яприймачів. Крім того, до статевої системи належать шкірні залози пояска та статеві щетинки.
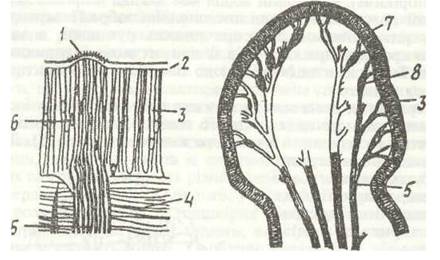
Рис. 26. Чутливі сенсили Lumbricus terrestris:
а - зріз чутливої бруньки; б- скупчення світлочутливих клітин на нервах простомія; 1 -чутливі палички; 2 - кутикула; 3 - епідерміс; 4 - кільцеві м'язи; 5 - нервові волокна; 6 - чутливі клітини; 7 - простимій; 8 - світлочутливі клітини
У дощовика Lumbricus terrestris чоловіча статева система представлена двома сім'яниками, що залягають у X та XI сегментах (рис. 27). Недозріла сперма потрапляє з них до сім'яних мішків — мішкоподібних випинань целомічного епітелію, які прикривають собою сім'яники. Тут сперма дозріває й нагромаджується. Навпроти кожного з сім'яників у целом відкривається миготлива лійка целомодукта, яка продовжується у вивідну протоку. Обидві протоки з кожного боку об'єднуються в один сім'япровід, що відкривається на черевній стороні XV сегмента.
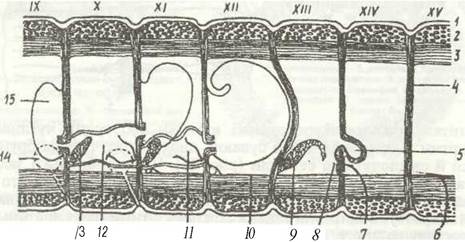
Рис. 27. Схема сагітального перерізу статевих сегментів (IX XV) Lumbricus terrestris:
1 - епідерміс; 2 - кільцеві м'язи; 3 - порожнинні м'язи; 4 - яйцевий мішок; 5 - чоловічий статевий отвір;
6 - жіночий статевий отвір; 8 - лійка яйцепроводу; 9 - яєчних 10 - сімяпровід; 11 - лійка сім'япроводу; 12 - сім'яна капсула; 13 - сім'яник; 14 - сім'яприймач; 15 - сім'яний мішок
Жіноча статева система складається з пари яєчників, що лежать у XIII сегменті. Яйцеклітини потрапляють до целома, де дозрівають і нагромаджуються у яйцевих мішках; звідси вони виходять назовні через яйцеводи, що відкриваються в целом лійкою, а назовні — отворами на XIV сегменті. До жіночої статевої системи належать також сім'яприймачі —дві пари глибоких шкірних упинань на черевній стороні IX—X сегментів, які не з'єднані з порожниною тіла. Вони призначені для зберігання сперми іншої особини при перехресному заплідненні.
При копуляції два черви з'єднуються черевними сторонами, головними кінцями назустріч одне одному так, що поясок одного черва пролягає навпроти отворів сім'яприймачів другого (рис. 28). При цьому пояски виділяють багато слизу, який огортає тіла обох особин у вигляді двох муфт. З отворів сім'япроводів черви виділяють сперму, яка скороченням м'язів переноситься до пояска, де й потрапляє у слизову муфту, а звідти · — до сім'яприймачів партнера, які роблять ковтальні рухи й захоплюють сперму. Після цього черви розходяться. Отже, при копуляції відбувається лише обмін спермою, а не запліднення. Коли у черва дозрівають яйцеклітини, поясок виділяє слизову муфту, в якій також є поживні речовини для зародка. Черв починає зсувати муфту вперед. При цьому до неї спочатку потрапляють яйцеклітини, а потім сперма із сім'яприймачів, і таким чином у муфті відбувається запліднення. Далі черв скидає муфту через головний кінець, її кінці злипаються й утворюється кокон, де й дозрівають яйця.
У деяких видів олігохет відоме самозапліднення. Є види, що здатні до партеногенезу. Дуже рідко трапляються олігохети (наприклад, з родини Eudrilidae), в яких жіночі статеві протоки з'єднуються з сім’яприймачами, через що в них спостерігається внутрішнє самозапліднення.
Розвиток у малощетинкових відбувається без метаморфозу. Яйця розвиваються всередині яйцевого кокона, з якого виходять повністю сформовані черви. У дрібних водяних олігохет, наприклад із родини Naididae, яйця багаті на жовток, який використовує зародок під час ембріонального розвитку. У більших за розміром червів, наприклад Lumbricidae, яйця бідні на жовток, але кокон має поживну білкову рідину. Зародок (рис.29), що розвивається всередині кокона, має рот, глотку й середню кишку, він активно заковтує білок і тому зветься «прихованою личинкою», яка перед виходом із кокона перетворюється на молодого черва. Молоді дощовики виходять назовні, розриваючи оболонку кокона.
Рис. 29. Зародок Lumbricus із черевної сторони:
1 - рот, 2 - глотка; 3 - ектодермальні смужки; 4 - ентодерма; 5 - ектодермальні телобласти;
6 - мезодермальні телобласти; 7- мезодермальні смужки
Крім статевого розмноження, у водяних олігохет спостерігається нестатеве, шляхом поділу тіла. У деяких видів (Lumbriculus, Enchytraeus) тіло розпадається на кілька фрагментів, кожен з яких згодом відновлює втрачені частини. Це явище зветься архітомією. В інших випадках тіло поділяється перетяжкою, проте обидві частини залишаються з’єднаними, передня нарощує задній кінець, а задня — передній. Потім молоді особини розходяться, однак іноді ще до цього кожна з них, у свою чергу, поділяється, утворюються ланцюжки з кількох особин (наприклад, у Aelosoma). Такий спосіб нестатевого розмноження зветься паратомією.
Загальновизнаного поділу класу олігохет на ряди до цього часу немає, тому розглянемо найважливіші родини.
Широко відомі представники родини Naididae, що населяють в основному прісні водойми, але можуть мешкати в прибережній зоні моря або в його опріснених ділянках. Так, види роду Nais у великій кількості трапляються в різних типах річок і стоячих водойм. Це невеликі (10—15 мм) білуваті прозорі тварини з пучечками довгих тоненьких щетинок, на простоміумі є пара очей. Вони повзають по субстрату, водоростях, можуть і плавати. Живляться одноклітинними водоростями, найпростішими, детритом.
До родини трубочників (Tubificidae) належить понад 300 видів морських та прісноводних риючих червів. Більшість трубочників живуть у прісних водоймах. Найбільш поширений рід Tubifex. Це невеличкі черви, довжина тіла яких досягає 2—5 см. В Україні відомо близько 40 видів цього роду, їх багато в замулених ґрунтах, де обмаль кисню. Всі вони мають в крові гемокруорин, який акумулює кисень. Переднім кінцем тіла трубочники занурені в мул, а заднім, в якому є багато шкірних кровоносних капілярів, роблять коливальні рухи у воді, полегшуючи газообмін. Живляться трубочники мулом, засвоюючи з нього органічні рештки. Вони можуть витримувати значне забруднення водойм різними речовинами (нафтопродукти, пестициди, миючі засоби тощо). У багатих на органіку водоймах їх концентрація досягає 100 тис. особин на 1 м2 дна. Серед трубочників є й морські форми. У прибережній смузі тропічних морів мешкають види роду Phalodrilus, для яких характерний своєрідний симбіоз із бактеріями, що селяться під кутикулою червів. Ці бактерії здатні засвоювати сірководень, що утворюється при розкладі решток організмів. Кишечник у Phalodrilus відсутній, і вони живляться за рахунок симбіонтів.
Представники родини ракових п'явок — Branchiobdellidae — мешкають на зябрах і поверхні тіла прісноводних вищих раків. Пристосування до ектопаразитизму різко змінило їхню будову: раніш цих олігохет вважали п'явками, їхнє тіло невелике (не більше 10—12 мм), потовщене, складається з головного відділу, що утворився злиттям простоміуму й чотирьох сегментів, та 11 сегментів тулуба, на кінці якого є присосок. Рот має хітинові верхню та нижню щелепи. Молоді особини живляться детритом, дорослі — кров'ю раків.
Види родини Enchytreidae мешкають у морях, прісних водоймах і Грунтах, їх налічується понад 400 видів. Це білуваті черви завдовжки 2—3 мм, школи до 45 мм, із добре розвиненими щетинками. Деякі види, особливо ґрунтові, розмножуються у величезних кількостях. Вони живляться гумусом. Часто акваріумісти розводять їх у горщиках із квітковою землею як корм для риб, тому їх звуть «горщиковими червами».
Найбільш відомі так звані дощові черви, які відіграють велику роль у ґрунтоутворенні. Вони належать до кількох родин і не становлять єдиної систематичної групи. Най- відоміші з них справжні дощові черви (родина Lumbricidae) мають довжину від 20 мм до 1 м і товщину від 1 до 20 мм. Усі вони сапрофаги (живляться органічними рештками). За типами живлення розрізняють дві групи видів. Одні види, до яких належить широко відомий дощовик (Lumbricus terrestris), живляться рослинним спадом, навіть іноді зеленими частинами, затягуючи їх із поверхні ґрунту в нірки, їх тіло дуже пігментоване, головна лопать добре розвинена; вони здатні виходити на поверхню, мешкати в підстилці й навіть у гнилій деревині (Dendrobaena). Види другої групи живляться ґрунтовим перегноєм; пігментація їх тіла слабо розвинена, простоміум невеликий, шкірно-м’язовий мішок тонший, ніж у попередньої групи; вони мешкають у товщі ґрунту (більшість видів Allolobophora), деякі види мають постійні глибокі (до 1 м) ходи.
Черви здійснюють вертикальні міграції; чим ґрунт сухіший або чим нижча його температура, тим глибше вони проникають у землю. Деякі види можуть утворювати на глибині спеціальні капсули, в яких переживають несприятливі пори року (зима, посушливий сезон тощо). При інтенсивному живленні черви викидають на поверхню горбки пропущеного через кишечник ґрунту — копроліти. За наявністю та кількістю останніх можна встановити чисельність самих червів.
Дощові черви поширені в різних ґрунтах, навіть у пустельних; у деяких ґрунтах їх чисельність величезна, наприклад у чорноземах вона досягає 10 млн на 1 га. В Україні відомо понад 50 видів дощових червів, із них цікаві ендемічні види Карпат (Helodrilus cemosvitovianus, Allolobophora car- pathica) та рівнинної частини нашої країни (Allolobophora leoni та ін.). Ці види потенційно потребують охорони; деякі занесено до Червоної Книги України. Цікаво, що відомий з Альп і Карпат вид Eisenia submontana здатний світитися в темряві.
Зовнішньо подібний до Lumbricidae черв Criodrilus lacuum з родини Criodrilidae завдовжки 12—32 см веде водяний спосіб життя, мешкає в мулі прісноводних та солонуватих водойм, дуже поширений в Україні. Види родини Megascole- cidae мають різні розміри, найбільші досягають довжини 3 м.
Вони живуть у ґрунтах на Півдні та Сході Азії, в Австралії, на островах Індонезії, один вид знайдено в Закавказзі, куди його завезла людина. Копроліти великих видів досягають 20—25 см висоти, мають вигляд башти.
Практичне значення. Трубочники та енхитреїди — добрий корм для риб, тому їх добувають і розводять. Трубочники через високу стійкість до забруднення перспективні для біологічного очищення стічних вод. Деяких велетенських мегасколецидів вживають у їжу аборигени Південно- Східної Азії й Південної Америки.
Деякі види дощових червів розводять у промислових умовах для використання у вигляді харчових додатків для худоби й навіть людини. Звичайний у нас вид Eisenia foetida (комерційна назва «каліфорнійський червоний») живе у гноївці. В результаті селекції виведено його породи, здатні розкладати різні види гною, покидьки. Ці черви з успіхом культивуються в промислових умовах і використовуються, в тому числі й в Україні, для переробки різноманітних біологічних відходів; пропущені через кишечник червів, ці відходи стають цінним комплексним добривом, а самі черви, перероблені на борошно, можуть бути використані як цінні добавки, які значно підвищують ефективність кормів для свійських тварин.
Проте найбільш відома роль дощових червів у ґрунтоутворенні, на що вперше звернув увагу Ч. Дарвін. У праці «Утворення перегною ґрунту при сприянні червів» він показав їхній позитивний вплив на родючість ґрунту. Пізніше цим питанням зацікавилося багато вчених. Черви спушують та перемішують ґрунт, поліпшуючи його аерацію й проникнення в глибинні шари води: загальна довжина ходів червів на 1 м2 може сягати 1—8 км. У пропущеній через кишечник землі збільшується вміст мінеральних речовин, необхідних рослинам, зменшується кислотність ґрунту. За рік черви пропускають через себе шар ґрунту від одного до семи метрш завтовшки; їхня біомаса залежно від типу ґрунту становить від 10—20 до 2000—3000 кг/га.
Негативна роль олігохет незначна. Відомо, що більшість видів дощових червів під час розмноження (у вас —червень- липень) стає отруйною і може спричинити загибель домашніх птахів. Деякі малощетинкові є проміжними хазяями гельмінтів. Так, лроцеркоїди паразита коропових риб Caryophyllaeus (Cestoda) мешкають у целомі Tubifex, у дощових червах личинки нематод, що викликають тяжкі захворювання свиней (Metastrongylus) та курей (Syngamus).