Зоология позвоночных - В. М. Константинов 2011
Хордовые
Наземные, или четвероногие, позвоночные
Класс Земноводные, или амфибии
Общая характеристика
Земноводные — немногочисленная группа наиболее примитивных наземных позвоночных. Группа в целом демонстрирует черты переходности от водного образа жизни к наземному (что отражено в их названии). Подавляющее большинство амфибий обитают, в зависимости от стадий жизненного цикла, то в воде, то на суше. В течение жизни они, как правило, претерпевают метаморфоз, превращаясь из чисто водных личинок во взрослые формы, обитающие большей частью вне воды. В связи с этим в дыхательной системе происходит смена жаберного дыхания на легочное, и соответственно изменяется кровеносная система. В опорно-двигательной системе формируются пятипалые конечности, существенно видоизменяется система органов чувств. Однако степень приспособления к жизни на суше у взрослых форм в общем невелика.
Общее число видов современных амфибий примерно 4 500. Они объединяются в три отряда: Хвостатые (Caudata, или Urodela), Безногие (Apoda) и Бесхвостые (Апига, или Ecaudata).
Строение земноводных
Кожные покровы. Кожа у всех земноводных голая, лишенная чешуй. Многослойный эпидермис богат многоклеточными слизистыми железами (рис. 68), погруженными в кориум. Значение кожных желез многообразно. Они создают на поверхности кожи жидкостную пленку, без которой невозможен газообмен. Эта пленка в некоторой мере предохраняет тело от иссушения. Слизь помогает коагуляции инородных частиц на поверхности тела, обладает бактерицидными свойствами и защищает тело от проникновения через кожу патогенных микробов. Ядовитые кожные железы в значительной степени защищают амфибий от хищников. Часто ядовитые амфибии имеют яркую окраску, предупреждающую хищников об опасности при схватывании добычи. Установлено, что у некоторых бесхвостых амфибий верхний слой эпидермиса ороговевает. Наиболее сильно это развито у жаб, роговой слой у которых на спине составляет примерно 60 % всей поверхности эпидермиса. У большинства амфибий слабое ороговение эпидермиса не препятствует проникновению через кожу воды, в расположенные под ней лимфатические мешки.
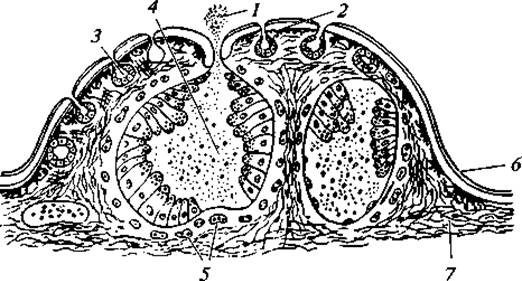
Рис. 68. Разрез через кожу саламандры:
1 — выступающий наружу секрет железы; 2 — пигментный слой; 3 — слизистые кожные железы; 4 — ядовитая кожная железа; 5 — перерезанные кровеносные сосуды; 6 — эпидермис; 7 — волокнистый слой кожи
Скелет. По сравнению со скелетом рыб скелет земноводных имеет ряд изменений. В осевом скелете позвоночник земноводных в связи с полуназемным образом жизни более расчленен. Он включает шейный, туловищный, крестцовый и хвостовой отделы (рис. 69). Шейный отдел представлен одним позвонком, его тело невелико и несет две сочленовные ямки, при помощи которых позвонок сочленяется с черепом. Число туловищных позвонков различно: наименьшее — у бесхвостых (обычно 7), наибольшее — у безногих (более 100). Единственный крестцовый позвонок (у безногих отсутствующий) несет длинные поперечные отростки, к которым причленяются подвздошные кости таза. Хвостовой отдел хорошо выражен у хвостатых, у безногих он очень мал, а у бесхвостых представлен одной косточкой — уростилем во время эмбрионального развития она закладывается в виде ряда отдельных позвонков, тела которых впоследствии сливаются.
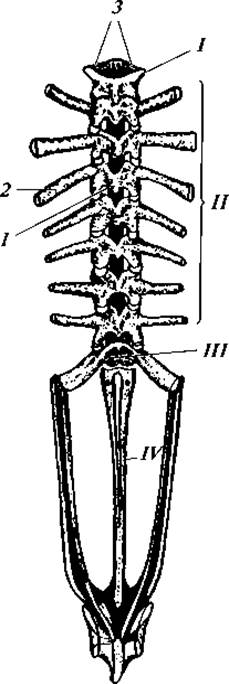
Рис. 69. Позвоночный столб лягушки вместе с тазовым поясом, вид со спинной стороны:
I — шейный отдел (из одного позвонка); II — туловищный отдел; III — крестец; IV — уростиль; I — остистый отросток третьего туловищного позвонка; 2 — поперечный отросток того же позвонка; 3 — сочленовные поверхности на первом шейном позвонке
Форма позвонков у разных представителей класса Земноводные чрезвычайно вариабельна. У низших амфибий (безногие, низшие хвостатые) она бывает амфицельной; в этом случае между позвонками пожизненно сохраняется хорда. У бесхвостых позвонки процельные. вогнутые спереди и выгнутые сзади; у высших хвостатых — опистоцельные, т. е. выгнутые спереди и вогнутые сзади. Существует множество отступлений от этой обшей характеристики, например, у примитивной новозеландской лягушки леопельмы (Leopelma) позвонки амфицельные. Настоящих ребер у бесхвостых амфибий нет, очень короткие есть у безногих; у хвостатых развиваются короткие «верхние» ребра.
Мозговой череп. Мозговая коробка в значительной своей части пожизненно остается хрящевой (рис. 70). Это обусловливается слабым развитием хондральных и накладных окостенений. В первичном мозговом черепе развиваются следующие хондральные кости. В затылочной области только две боковые затылочные кости; места, соответствующие основной и верхней затылочным костям рыб, остаются хрящевыми. В области слуховой капсулы формируется одна небольшая ушная кость, большая же часть капсулы остается хрящевой. В передней части глазницы у бесхвостых развивается одна клинообонятельная кость; у хвостатых эта кость парная. Обонятельная капсула хрящевая.
Покровных костей также немного. Крышу черепа составляют теменные и лобные кости, которые у бесхвостых срастаются в лобнотеменные кости. Впереди от них находятся носовые кости, у безногих они срастаются с предчелюстными костями. По бокам задней части черепа расположены чешуйчатые кости, особенно сильно развитые у безногих. Дно черепа выстилает большой парасфеноид, а впереди от него лежат парные сошниковые кости.
В формировании дна черепа принимают участие и кости висцерального скелета — нёбные и крыловидные. Первые прилегают к сошникам, вторые — к чешуйчатым костям. Они развиваются на нижней поверхности нёбно-квадратного хряща. Функции верхних челюстей выполняют, как у костных рыб, предчелюстные (или межчелюстные) и верхнечелюстные кости.
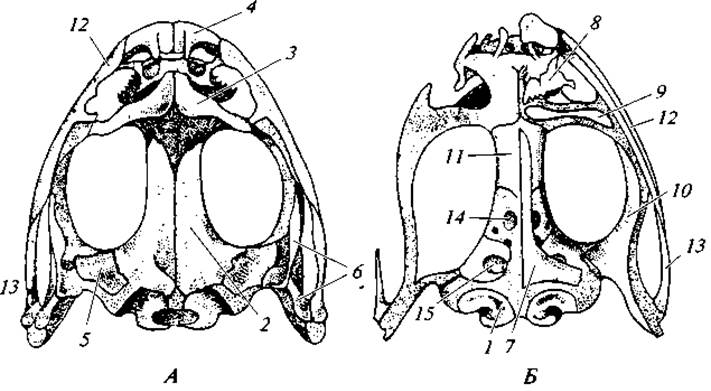
Рис. 70. Череп лягушки:
А — вид сверху; Б — вид снизу (накладные кости с одной стороны удалены). Обозначение костей: 1 — боковая затылочная; 2 — лобно-теменная; 3 — носовая; 4 — межчелюстная; 5 — переднеушная; 6 — чешуйчатая; 7 — парасфеноид (левая его половина); 8 — сошник; 9 — нёбная; 10 — крыловидная; 11 — клинообонятельная; 12 — верхнечелюстная; 13 — квадратночелюстная; 14 — отверстие для выхода зрительного нерва; 15 — отверстие для тройничного нерва
Нижняя челюсть представлена меккелевым хрящом, который прикрыт снаружи зубной и угловой костями.
Череп амфибий аутостилический, т.е. нёбно-квадратный хрящ непосредственно прирастает к мозговому черепу. В связи с аутостилией черепа подъязычная дуга не принимает участия в прикреплении челюстного аппарата к черепу.
Верхний элемент этой дуги — подвесок (гиомандибуляре) — превращен в маленькую кость — стремя, которая проксимальным концом упирается в слуховую капсулу, а наружним (дистальным) — в барабанную перепонку. В связи с формированием полости среднего уха эта кость находится внутри указанной полости и выполняет роль слуховой косточки. Таким образом, гиомандибуляре (подвесок) выходит из системы четвертой (подъязычной) висцеральной дуги (см. рис. 70).
Нижние элементы подъязычной дуги и жаберных дуг видоизменяются в подъязычную пластинку и ее рожки. Эта пластинка расположена между ветвями нижней челюсти. Передние ее рожки, загибаясь вверх и охватывая с боков кишечную трубку, прикреплены к слуховым капсулам. Изменения в висцеральном скелете сопровождаются утратой жаберных крышек.
Таким образом, череп амфибий отличается от черепа большинства костных рыб: 1) слабым развитием хондральных и кожных окостенений; 2) аутостилией; 3) видоизменением подъязычной и жаберных дуг, превращенных частью в слуховой, частью в подъязычный аппарат; 4) редукцией жаберной крышки.
Пояса конечностей. Плечевой пояс имеет вид дуги, обращенной вершиной к брюшной поверхности животного (рис. 71). Каждая половина дуги (левая и правая) состоит из следующих основных элементов. Верхняя (спинная) часть представлена лопаткой с широким надлопаточным хрящом. Нижняя (брюшная) часть включает коракоид и лежащий впереди него прокоракоид. У бесхвостых между предгрудиной и лопаткой расположена тонкая палочковидная ключица. Перечисленные элементы пояса сходятся в точке прикрепления плечевой кости и формируют сочленовную ямку. Впереди от места соединения левого и правого коракоидов находится предгрудина, а сзади — грудина. Обе эти кости заканчиваются хрящами. Плечевой пояс в отличие от костных рыб лежит свободно в толще мускулатуры и не связан с черепом. В связи с отсутствием или неполным развитием ребер грудной клетки у амфибий нет.
Тазовый пояс (рис. 72) образован тремя парными элементами, сходящимися в области вертлужной впадины, которую они и образуют. Длинные подвздошные кости своими проксимальными (передними) концами прикреплены к поперечным отросткам единственного крестцового позвонка. Направленный вперед и вниз лобковый элемент пояса у лягушек остается хрящевым. Сзади него расположена седалищная кость. Такое расположение элементов тазового пояса характерно для всех наземных позвоночных.
Скелет свободных конечностей. Скелет свободных конечностей типичен для наземных позвоночных и существенно отличен от скелета конечностей рыб. В то время как конечности рыб представляют в схеме простые одночленные рычаги, перемещающиеся только относительно корпуса тела и не несущие мускулатуры, конечности наземных позвоночных представляют собой многочленные рычаги с достаточно мощными мышцами. В этом случае не только вся конечность перемещается относительно тела, но и отдельные элементы конечности перемещаются друг относительно друга.
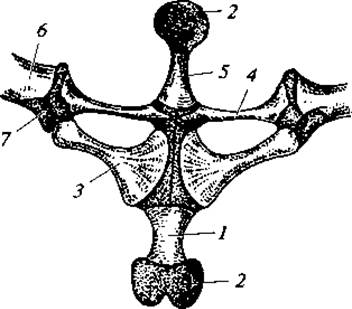
Рис. 71. Плечевой пояс лягушки, вид спереди:
1 — грудина; 2 — хрящевые передняя и задняя части грудины; 3 — коракоид; 4 — ключица лежит на прокоракоиде; 5 — предгрудина; 6 — лопатка; 7 — сочленовная впадина на лопатке для плеча (хрящ покрыт точками)
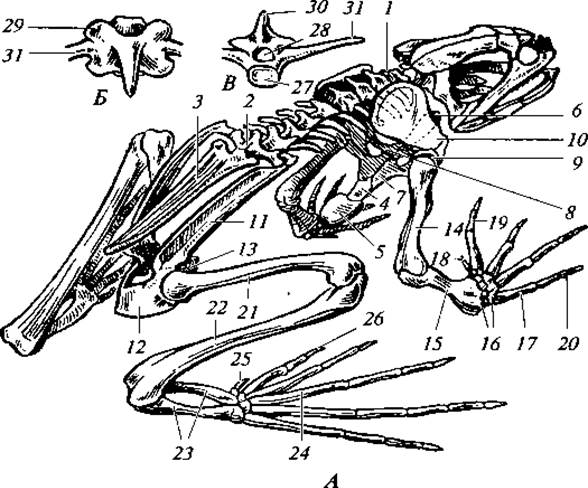
Рис. 72. Скелет лягушки:
А — общий вид; Б — вид позвонка сверху; В — вид позвонка спереди; 1 — шейный позвонок; 2 — крестцовый позвонок; 3 — уростиль; 4 — грудина; 5 — хрящевая задняя часть грудины; 6 — предгрудина; 7 — коракоид; 8 — прокоракоид; 9 — лопатка; 10— надлопаточный хрящ; 11 — подвздошная кость; 12— седалищная кость; 13 — лобковый хрящ; 14 — плечевая кость; 15 — предплечье (лучевая + локтевая кости); 16 — запястье; 17 — пясть; 18 — зачаточный I палец; 19 — II палец; 20 — У палец; 21 — бедро; 22 — голень (большая и малая берцовые кости); 23 — предплюсна; 24 — плюсна; 25 — рудимент добавочного пальца; 26 — I палец; 27 — тело позвонка; 28 — спинно-мозговой канал; 29 — сочленовная площадка; 30 — остистый отросток; 31 — поперечный отросток
В схеме скелет пятипалой конечности состоит из нескольких основных отделов. В частности, у амфибий скелет передней и задней конечностей имеет отделы (см. рис. 72):
I — плечо в передней конечности, бедро — в задней; этот отдел всегда состоит из одной кости, которая проксимальным концом причленена к поясу;
II — предплечье в передней конечности, голень в задней. В типичном случае отдел состоит из двух параллельно расположенных костей: предплечье — из локтевой и лучевой, голень — из большой берцовой и малой берцовой;
III — кисть в передней конечности и стопа в задней; отдел состоит из трех подотделов:
1) запястье — в передней конечности, предплюсна — в задней; этот подотдел представлен в типичном случае 9—10 мелкими косточками, расположенными в три ряда;
2) пясть — в передней конечности, плюсна — в задней; в типичном случае подотдел состоит из 5 удлиненных костей, расположенных в один ряд, как бы веером, от запястья или предплюсны;
3) фаланги четырех-пяти пальцев представляют как бы продолжение пясти или плюсны и включают три—пять рядов косточек в каждом.
Скелет конечностей хвостатых амфибий почти полностью соответствует приведенной схеме. У лягушек произошли следующие изменения: оба элемента предплечья и голени срастаются в одну кость, срастается между собой и большая часть костей запястья (до 6 костей) и предплюсны (до 3 костей), перед первым пальцем задней конечности имеется рудимент добавочного пальца. Эти особенности имеют вторичный характер и связаны, видимо, с приспособлением лягушек к передвижению прыжками.
Мышечная система. Мышечная система земноводных существенно отличается от мышечной системы рыб двумя основными особенностями, связанными с передвижением животных при помощи пятипалых конечностей и в известной мере по твердому субстрату. Во-первых, развивается мощная и сложно организованная мускулатура на свободных конечностях. Во-вторых, в связи со сложными движениями мускулатура тела более дифференцирована, а характерная для рыб ее сегментация у амфибий нарушена. Метамерия мышечной системы выражена более четко у хвостатых и безногих. В свою очередь, у бесхвостых ее можно увидеть только в немногих местах туловища у взрослых форм и в личиночном состоянии.
Органы пищеварения. Ротовая щель ведет в обширную ротоглоточную полость, которая, сужаясь, переходит в пищевод. В ротоглоточную полость открываются хоаны, евстахиевы отверстия (полости среднего уха) и гортанная щель. Сюда же открываются отсутствующие у рыб протоки слюнных желез. Их секрет служит для смачивания пищевого комка и не воздействует на пищу химически. На дне ротоглоточной области расположен настоящий язык, имеющий собственную мускулатуру. Форма языка разнообразна. У некоторых хвостатых он прикреплен неподвижно, у других имеет вид гриба, сидящего на тонкой ножке. У лягушек язык прикреплен одним концом ко дну рта, а свободная его часть в спокойном состоянии обращена внутрь. У всех амфибий язык выделяет клейкое вещество и служит для ловли мелких животных. Немногие амфибии не имеют языка.
На межчелюстных и верхнечелюстных костях, на сошнике, а у некоторых и на нижней челюсти сидят зубы. Они имеют вид мелких однообразных конусов, вершины которых несколько загнуты назад. Некоторые виды, например жабы, зубов на костях челюстей не имеют. При глотании проталкиванию пищевого комка из ротоглоточной области в пищевод помогают глазные яблоки, которые отделены от этой полости только тонкой слизистой оболочкой и с помощью специальных мышц могут несколько втягиваться внутрь ротоглотки.
Короткий пищевод впадает в сравнительно слабо отграниченный желудок. Собственно кишечник относительно более длинный, чем у рыб. В петле переднего (тонкого) отдела лежит поджелудочная железа. Крупная печень имеет желчный пузырь, ее проток впадает в переднюю часть тонкой кишки (в двенадцатиперстную кишку). З желчный проток впадают и протоки поджелудочной железы, которая самостоятельного сообщения с кишечником не имеет (рис. 73). Второй отдел кишечника — толстый — отграничен от тонкого отдела нечетко. Наоборот, третий, прямой, отдел обособлен хорошо. Он заканчивается клоакой.
Органы дыхания. Органы дыхания амфибий разнообразны. Во взрослом состоянии большинство видов дышат легкими и через кожу. Легкие представляют собой парные мешки с тонкими ячеистыми стенками. При относительно малой поверхности легких очень велико значение кожного дыхания. Отношение поверхности легких к поверхности кожи у амфибий равно 2:3 (в то время как у млекопитающих внутренняя поверхность легких в 50—100 раз больше поверхности кожи). У зеленой лягушки через кожу поступает 51 % кислорода. Важна роль кожи в выделении из организма углекислоты: через кожу ее выделяется 86 %, через легкие — 14 %.
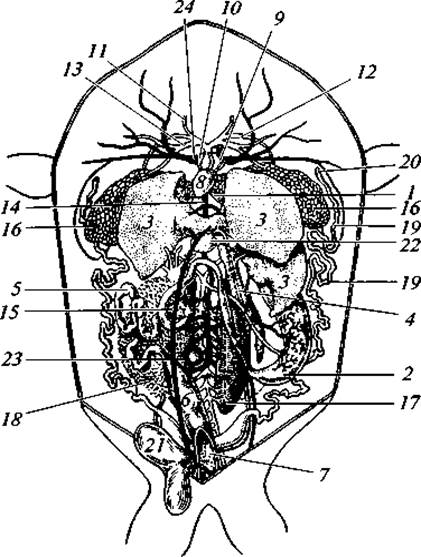
Рис. 73. Общее расположение внутренностей у самки лягушки:
1 — пищевод; 2 — желудок; 3 — лопасти печени; 4 — поджелудочная железа; 5 — тонкая кишка; 6 — толстая (прямая) кишка; 7 — клоака (вскрытая); 8 — желудочек сердца; 9 — левое предсердие; 10 — правое предсердие; 11 — сонная артерия (правая); 12 — левая дуга аорты; 13 — легочно-кожная дуга (правая); 14 — задняя (нижняя) полая вена; 15 — брюшная вена; 16 — легкие; 17 — левая почка (задний ее конец); 18 — правый яичник; 19 — левый яйцевод; 20 — его устье (воронка); 21 — кочевой пузырь; 22 — желчный пузырь; 23 — селезенка; 24 — передняя полая вена (правая)
Таблица 6
Зависимость потребления О2 и выделения СО2 от температуры среды у зеленой лягушки (Rana)
(по Л. Проссеру, 1977)
Температура среды, С |
Потребление О2, мкл/(г ∙ ч) |
Выход СО2, мкл/(г ∙ ч) |
||
кожа |
легкие |
кожа |
легкие |
|
5 |
15 |
10 |
15 |
5 |
15 |
22 |
28 |
30 |
10 |
25 |
40 |
80 |
50 |
35 |
Кожное дыхание имеет большое функциональное значение не только в связи с несовершенством легких, но и как приспособление, обеспечивающее окисление крови при длительном нахождении животного в воде, например во время спячки или затаивания в водоеме при преследовании наземными хищниками. В этих случаях происходит только кожное дыхание, и правое предсердие (куда через полые передние вены вливается окисленная кровь кожной вены - см. выше) получает насыщенную кислородом кровь.
У американских безлегочных саламандр и у дальневосточного тритона легкие полностью атрофированы, и газообмен у них целиком происходит через кожу и слизистую ротовой полости.
Способность кожи и легких поглощать кислород (О2) и выделять углекислый газ (СО2) зависит у амфибий от температуры среды (табл. 6). Как видно, при температуре окружающей среды 5 °С через кожу зеленой лягушки поступает в 1,5 раза больше кислорода, чем через легкие. При температуре 25 °С, наоборот, через легкие поступает кислорода в 2 раза больше, чем через кожу. Иначе обстоит дело с выходом СО2. При повышении температуры от 5 до 25 °С значение кожи в выходе углекислого газа возрастает только в 3,3 раза, а легких — в 7 раз. При любых температурах роль кожи в удалении из организма лягушки СО2 заметно больше, чем легких.
Личинки амфибий дышат при помощи ветвистых наружных жабр, которые у подавляющего большинства видов в последующем исчезают, а у протеев и сирен сохраняются пожизненно. Амфиумы во взрослом состоянии наряду с легкими имеют и внутренние жабры.
В связи с отсутствием грудной клетки механизм легочного дыхания весьма своеобразен. Роль насоса выполняет ротоглоточная полость, дно которой то опускается (воздух при открытых ноздрях засасывается), то поднимается (воздух при закрытых ноздрях проталкивается в легкие). Поэтому череп амфибий при небольшой высоте чрезвычайно широк: эффективность легочного дыхания тем больше, чем больше расстояние между ветвями нижней челюсти. К примеру наибольшей ширины оно достигает у жаб, ороговевшая кожа которых имеет малую дыхательную нагрузку.
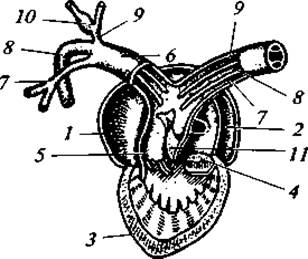
Рис. 74. Схема строения вскрытого сердца лягушки:
1 — правое предсердие; 2 — левое предсердие; 3 — желудочек; 4 — клапан, закрывающий общее отверстие, ведущее из обоих предсердий в желудочек; 5 — артериальный конус; 6 — общий артериальный ствол; 7 — кожно-легочная артерия; дуга аорты; 9 — общая сонная артерия; 10 — сонная «железа»; 11 — спиральный клапан артериального конуса
Кровеносная система. Сердце у всех амфибий трехкамерное, состоит из двух предсердий и одного желудочка (рис. 74). У низших форм (безногие и хвостатые) левое и правое предсердия разделены не полностью. У бесхвостых перегородка между предсердиями полная, но у всех земноводных оба предсердия сообщаются с желудочком одним общим отверстием. Кроме указанных основных отделов сердца имеется венозная пазуха. Она принимает венозную кровь и сообщается с правым предсердием. К сердцу примыкает артериальный конус, в него кровь изливается из желудочка. Артериальный конус имеет спиральную складку, участвующую в распределении крови в выходящие из него три пары сосудов. Сердечный индекс (отношение массы сердца к массе тела в процентах) варьирует у разных видов и зависит от двигательной активности животного. Так, у сравнительно мало перемещающихся травяной и зеленой лягушек он равен 0,35 — 0,55 %, а у полностью наземной (кроме периода размножения) и активной зеленой жабы — 0,99 %.
У высших — бесхвостых — амфибий от артериального конуса берут начало три пары артериальных дуг.
Первая пара (считая от головы к хвосту) несет кровь к голове, это — сонные артерии. Они гомологичны первой паре жаберных артерий рыб. Вторая пара, также отходящая от брюшной стороны артериального конуса, гомологична второй паре жаберных сосудов рыб и носит название системных дуг аорты. От них отходят подключичные артерии, несущие кровь к плечевому поясу и передним конечностям. Правая и левая системные дуги, описав полукруг, соединяются вместе и образуют спинную аорту, расположенную под позвоночником; от нее отходят артерии, идущие к внутренним органам. Последняя, третья, пара, гомологичная четвертой паре жаберных артерий рыб, отходит не от брюшной, а от спинной стороны артериального конуса. Она несет кровь в легкие и представляет собой легочные артерии. От каждой легочной артерии отходит сосуд, несущий венозную кровь в кожу: это кожные артерии.
У хвостатых амфибий, имеющих легкие, схема расположения артериальных сосудов в основном такая же. Но в отличие от бесхвостых у них сохраняется пара артерий, соответствующая третьей паре жаберных сосудов, и, таким образом, общее число парных артериальных стволов у них равно четырем, а не трем, как у бесхвостых. Кроме того, легочные артерии сохраняют связь с системными дугами аорты через посредство так называемых боталовых протоков (см. рис. 77).
У хвостатых амфибий, у которых пожизненно сохраняются жабры, схема кровообращения очень близка к таковой у рыб и личинок высших амфибий. У них от брюшной аорты отходят четыре пары артериальных дуг. Сонные артерии отходят от выносящей части первой дуги, и сохраняются сонные протоки. С появлением легких образуются легочные артерии, берущие начало от четвертой артериальной дуги. Схема кровообращения в этом случае почти тождественна таковой двоякодышащих рыб (см. рис. 44).
Венозная система низших амфибий похожа на венозную систему двоякодышащих рыб. Хвостовая вена делится на две воротные вены почек. Из почек кровь поступает в непарную заднюю полую вену и в парные задние кардинальные вены. Последние на уровне сердца сливаются с парными яремными, подключичными и кожными венами и образуют кювьеровы протоки, изливающие кровь в венозную пазуху. От кишечника кровь собирается по подкишечной и брюшной венам, которые, сливаясь, образуют воротную вену печени. Из печени кровь поступает в полую вену по печеночной вене.
У бесхвостых амфибий кардинальные вены не сохраняются, и вся кровь из туловищной области собирается в конечном счете в заднюю полую вену, впадающую в венозную пазуху. Как и в предыдущем случае, имеются брюшная и подкишечная вены, образующие воротную систему кровообращения в печени. В связи с отсутствием кардинальных вен у бесхвостых не образуются и кювьеровы протоки. Яремные вены, сливаясь с подключичными, образуют в этом случае парные передние полые вены, впадающие в венозный синус, или пазуху. В верхние полые вены впадают и кожные вены соответствующей стороны, которые несут не венозную, а артериальную кровь.
Легочные вены несут кровь непосредственно в левое предсердие. Венозная кровь (с довольно значительной примесью окисленной крови, поступающей из кожных вен в передние полые вены) изливается в венозную пазуху (синус), а оттуда в правое предсердие. При сокращении предсердий венозная и артериальная кровь изливается через общее для обоих предсердий отверстие в желудочек. К его правой части примыкает артериальный конус, в который поступает сначала наиболее венозная кровь, направляющаяся далее в открытое отверстие кожно-легочных артерий. Отверстия остальных артериальных дуг в это время прикрыты спиральным клапаном артериального конуса. При дальнейшем сокращении желудочка давление в артериальном конусе возрастает, спиральный клапан сдвигается и открываются отверстия системных дуг аорты, через которые поступает смешанная кровь из центральной части желудочка. Дальнейшее сдвигание спирального клапана освобождает устья сонных артерий, куда и проходит наиболее окисленная кровь, выходящая в артериальный конус в последнюю очередь из левой части желудочка. Однако полного разделения артериальной и венозной крови не происходит: к органам тела поступает смешанная кровь.
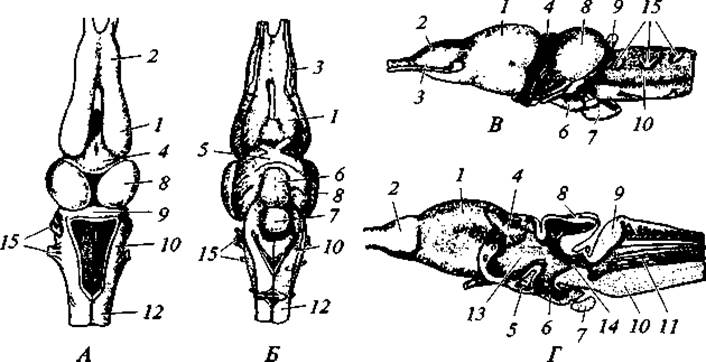
Рис. 75. Головной мозг лягушки:
вид сверху (А), снизу (Б), сбоку (В), в продольном разрезе (Г); 1 — полушария переднего мозга; 2 — обонятельная доля; 3 — обонятельный нерв; 4 — промежуточный мозг; 5 — зрительная хиазма; 6 — воронка; 7 — гипофиз; 8 — средний мозг; 9 — мозжечок; 10 — продолговатый мозг; 11 — четвертый желудочек; 12 — спинной мозг; 13 — третий желудочек; 14 — сильвиев водопровод; 15 — головные нервы
Скорость движения крови (важный показатель интенсивности метаболизма) у амфибий невелика. Об этом косвенно свидетельствует частота пульса. У травяной лягушки массой 50 г она равна 40 — 50 ударам в минуту. Для сравнения: у сходной по размерам птицы этот показатель равен примерно 500. У водных форм наблюдается брадикардия. Уровень артериального давления у амфибий также низок. У хвостатых он равен примерно 22/12 — 30/25, у бесхвостых — 30/20. Для сравнения: у чешуйчатых рептилий этот показатель составляет около 80/60 (Л. Проссер, 1978).
Нервная система. Головной мозг (рис. 75) характеризуется рядом Прогрессивных черт. Это выражается в относительно более крупных, чем у рыб, размерах переднего мозга, в полном разделении его полушарий и в том, что не только дно боковых желудочков, но и их бока и крыша содержат нервные клетки. Таким образом, у земноводных имеется настоящий мозговой свод — архипаллиум, который среди костных рыб свойствен только двоякодышащим. Средний мозг сравнительно небольших размеров. Мозжечок очень мал, а у некоторых хвостатых (у протеев) он практически незаметен. Слабое развитие этой части головного мозга связано с крайне однообразными, не сложными движениями амфибий. Последний отдел — продолговатый мозг — хорошо выражен. От головного мозга отходят десять пар головных нервов (І-Х).
Спинно-мозговые нервы у хвостатых и бесхвостых образуют плечевое и поясничное сплетения. Хорошо развита вегетативная нервная система, представленная нервными стволами, расположенными по бокам позвоночника.
Органы чувств у земноводных более развиты, чем у рыб. Органы обоняния амфибий представляют парные обонятельные капсулы. внутренняя поверхность которых выстлана обонятельным эпителием. Они сообщаются с наружной средой парными наружными ноздрями; от обонятельных капсул отходят внутренние ноздри (хоаны), сообщающиеся с ротоглоточной полостью. У амфибий, как и у всех наземных позвоночных, указанная система служит для восприятия запахов и для дыхания.
Органы боковой линии свойственны личинкам всех амфибий. Во взрослом состоянии они сохраняются только у водных форм хвостатых амфибий и немногих, тоже водных, бесхвостых. В отличие от рыб чувствующие клетки этого органа расположены не в углубленном канале, а поверхностно в коже.
Органы вкуса. Расположены в ротовой полости. Предполагается что лягушка воспринимает только горькое и соленое.
Органы зрения. Глаза амфибий имеют ряд особенностей, связанных с полуназемным образом жизни: 1) подвижные веки защищают глаза от высыхания и загрязнения; при этом, кроме верхнего и нижнего век, имеется еще третье веко, или мигательная перепонка. расположенная в переднем углу глаза; 2) есть слезная железа, секрет которой омывает глазное яблоко; 3) выпуклая (а не плоская, как у рыб) роговица и линзовидный (а не круглый, как у рыб) хрусталик. обе последние особенности определяют более дальнозоркое зрение амфибий (интересно, что в воде роговица у амфибий становится плоской); 4) аккомодация зрения достигается, как у акул, смещением хрусталика под действием ресничного мускула. Видимо, некоторые амфибии обладают цветовым зрением.
Орган слуха устроен значительно более сложно, чем у рыб, и приспособлен к восприятию звуковых раздражений в воздушной среде. Наиболее полно это выражено у бесхвостых амфибий. Кроме внутреннего уха, представленного, как и у рыб, перепончатым лабиринтом, у земноводных имеется еще среднее ухо. Последнее представляет собой полость, один конец которой открывается в ротоглотку, а другой подходит к самой поверхности головы и затянут тонкой перепонкой, которая называется барабанной. Полость эта образует изгиб, вершина которого находится у перепончатого лабиринта Верхняя часть полости от барабанной перепонки до перепончатого лабиринта носит название барабанной полости. В ней расположена палочковидная косточка — стремя, которая одним концом упирается в овальное окно внутреннего уха, другим — в барабанную перепонку. Нижняя часть полости среднего уха, открывающаяся в ротоглотку, именуется евстахиевой трубой.
Данные сравнительной анатомии и эмбриологии показывают, что полость среднего уха гомологична брызгальцу акуловых рыб, т. е. рудиментарной жаберной щели, лежащей между челюстной и подъязычной дугами, а слуховая косточка гомологична верхнему отделу подъязычной дуги — гиомандибуляре. На этом примере видно, что важное изменение органа может достигаться путем видоизменения и смены функций образований, имевшихся ранее у примитивных форм.
У безногих и хвостатых барабанная перепонка и барабанная полость отсутствуют, но слуховая косточка хорошо развита. Редукция среднего уха у этих групп, видимо, явление вторичное.
Органы боковой линии свойственны личинкам всех амфибий. Во взрослом состоянии они сохраняются только у водных форм хвостатых амфибий и немногих, тоже водных, бесхвостых. В отличие от рыб чувствующие клетки этого органа расположены не в углубленном канале, а поверхностно в коже.
Органы выделения. Органы выделения (рис. 76) устроены по типу их организации у хрящевых рыб. В зародышевом состоянии органами выделения служат пронефросы, у взрослых — мезонефросы с их типичными выводными путями — вольфовыми каналами.
Мочеточники открываются в клоаку.
Сюда же у высших наземных амфибий открывается мочевой пузырь. После его наполнения моча через то же отверстие выводится в клоаку и затем изгоняется наружу.
Число нефронов у низших (хвостатых) амфибий около 500, у высших (бесхвостых) — около 2 000. Такая заметная разница, видимо, определяется тем, что у хвостатых, которые теснее связаны с водоемами, имеет место и внепочечное (через кожу и жабры) выделение продуктов азотистого обмена. У хвостатых в отличие от бесхвостых нефроны (или хотя бы часть их) имеют нефростомы, т.е.
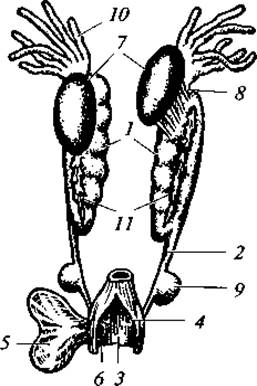
Рис. 76. Мочеполовая система самца лягушки:
1 — почки; 2 — мочеточник; 3 — полость клоаки; 4 — мочеполовое отверстие; 5 — мочевой пузырь; 6 — отверстие мочевого пузыря; 7 — семенники; 8 — семявыносящие каналы; 9 — семенной пузырек; 10 — жировое тело; 11 — надпочечники
Кронки, открывающиеся в полость тела (примитивный признак). Сосудистые клубки в боуменовых капсулах развиты хорошо, и амфибии выделяют много жидкой мочи. Укажем для примера, что у лягушек рода Rana скорость фильтрации плазмы крови в почках составляет около 35 мл/(кг ч).
Основным продуктом белкового обмена у амфибий служит мочевина, которая не очень токсична, но для выведения из организма требует большого количества воды, в которой она растворяется. Физиологически это вполне оправдано, так как поглощение воды организмом у амфибий в подавляющем случае не встречает затруднений.
Связь типа белкового обмена с условиями внешней среды доказывается и следующими примерами. У тритона осенью на суше доля аммиака в общих продуктах азотистого обмена равна 13 %, а при летнем водном существовании она возрастает до 26 %. У головастика доля аммиака равна 75 %, а у лягушонка, утерявшего хвост, с развитыми ногами — всего 16 %.
Органы размножения. У самцов парные семенники не имеют самостоятельных выводных путей. Семявыносящие канальцы проходя і через передний отдел почки и впадают в вольфов канал, который, таким образом, служит не только мочеточником, но и семявыносяшим каналом. Каждый вольфов канал у самцов перед впадением в клоаку образует расширение — семенной пузырек, в котором временно резервируется семя.
Над семенниками лежат жировые тела — образования неправильной формы желтого цвета. Они служат для питания семенников и развивающихся в них сперматозоидов. Величина жировых тел меняется по сезонам. Осенью они велики; весной же, во время интенсивного сперматогенеза, вещество их энергично расходуется и размеры жировых тел резко уменьшаются. Копулятивных органов у подавляющего большинства (кроме безногих) амфибий нет.
У самок развиваются парные яичники, над ними также лежат жировые тела. Созревшие яйца попадают в полость тела, откуда они поступают в воронкообразные расширения парных яйцеводов — мюллеровых каналов. Яйцеводы — длинные, сильно извитые трубки, задний отдел которых открывается в клоаку.
Из изложенного видно, что, как и у хрящевых рыб, у самцов земноводных мочевой и половой проток объединены и представляют собой единый вольфов канал, у самок вольфов канал выполняет функцию только мочеточника, а половые продукты выводятся через самостоятельный половой проток — яйцевод, или мюллеров канат.
Систематика и распространение современных амфибий
Современные амфибии представлены двумя подклассами. К подклассу Тонкопозвонковые (Lepospondyli) относятся два отряда: Хвостатые (Caudata, или Urodela) и Безногие (Apoda); к подклассу Дугопозвонковые (Apsidospondyli) относится отряд Бесхвостые (Ecaudata, или Anura).