Зоология позвоночных - В. М. Константинов 2011
Хордовые
Наземные, или четвероногие, позвоночные - Позвоночные с зародышевыми оболочками
Класс Птицы
Общая характеристика
Птицы — прогрессивная специализированная ветвь рептилий, приспособившихся к полету. Прогрессивные черты организации птиц, принципиально отличающие их от рептилий, заключаются: 1) в более высоком уровне развития нервной системы, а в связи с этим и в более разнообразном и совершенном приспособительном поведении; 2) высокой и постоянной температуре тела, связанной со значительно возросшей интенсивностью обмена веществ и с более совершенной терморегуляцией; 3) способности к полету, что не ведет в подавляющем большинстве случаев к потере возможности передвигаться по твердому субстрату или лазать; 4) более совершенном размножении: откладывании, насиживании яиц, обогреве, охране и выкармливании птенцов.
Указанные особенности позволили птицам, несмотря на относительную молодость класса, распространиться по всему земному шару и заселить разнообразные места обитания. В связи с широким распространением и разнообразием жизненной обстановки класс птиц характеризуется большим, чем рептилии, многообразием и обилием видов. Он включает в себя около 10 тыс. ныне живущих видов, объединенных в 30 — 35 отрядов.
Морфологически птицы характеризуются тем, что тело их покрыто перьями, передние конечности превращены в крылья, кости пневматические, череп с одним затылочным мыщелком, сердце четырехкамерное с одной правой дугой аорты, зубы у современных птиц отсутствуют и функционально замещены роговым клювом.
Морфофизиологический обзор
Кожные покровы и их производные. Кожа птиц тонкая, не имеет каких-либо костных образований и почти лишена желез. Исключение составляет лишь копчиковая железа, расположенная над корнем хвоста, секрет которой служит для смазывания перьев и для придания перьевому покрову водонепроницаемости. Копчиковая железа сильно развита у водоплавающих птиц, а у некоторых наземных видов, например страусов и дроф, обитающих в засушливом климате, она отсутствует.
Наряду с отсутствием в коже костных образований обильны и разнообразны роговые производные эпидермиса. Так, верхняя и нижняя челюсти покрыты роговыми чехлами, образующими клюв. На концах пальцев имеются когти, а на нижней части ног (на пальцах, цевке, а у некоторых на голени) — роговые щитки. Тело покрыто перьями, которые у подавляющего большинства видов располагаются не повсеместно, а лишь на некоторых участках — птерилиях. На других участках — аптериях — перьев вовсе или почти нет. Такое расположение перьев, свойственное летающим птицам, имеет приспособительное значение, так как облегчает сокращение мышц, подвижность кожи и перемещение перьев на туловище, связанное с движением крыльев. Подобное же значение имеют аптерии и при движениях задних конечностей и шеи (рис. 126).
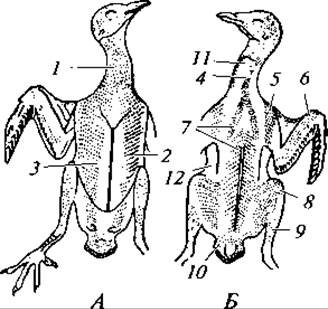
Рис. 126. Птерилии и аптерии голубя:
вид с брюшной (А) и спинной (Б) сторон; 1 — шейная птерилия; 2 — брюшная птерилия; 3 — брюшная аптерия; 4 — шейная птерилия; 5 — плечевая птерилия; 6 — крыловая птерилия; 7 — спинная птерилия; 8 — бедренная птерилия; 9 — голенная птерилия; 10 — хвостовая птерилия; 11 — шейная аптерия; 12 — боковая аптерия
Общее число контурных перьев больше у крупных птиц, чем у мелких. Так, у рубиногорлого колибри их около 100, у мелких воробьиных 1,5 — 2,5 тыс., у чаек 5 — 6 тыс., у уток 10 — 12 тыс., у лебедей 25 тыс.
Перья птиц различны по строению и функции. Снаружи тело покрыто контурными перьями, состоящими, из полого стержня, к которому более или менее симметрично прикреплены две боковые пластинки — опахала. Нижняя часть стержня погружена в кожу и носит название очина; большую верхнюю часть стержня, к которой прикреплены опахала, называют стволом (рис. 127). Опахало состоит из многочисленных длинных бородок первого порядка, на которых сидят бородки второго порядка. Последние снабжены очень мелкими крючочками, сцепляющими бородки второго порядка между собой. В итоге опахало представляет собой упругую эластичную пластинку (рис. 128).
Контурные перья являются основой оперения. Они предохраняют тело птиц от потери тепла и механических воздействий, образуют гребную лопасть крыла и рулевую плоскость хвоста. В зависимости от расположения на теле контурные перья разделяются на группы. Так, длинные перья, расположенные по заднему краю передних конечностей и формирующие лопасти крыльев, называют маховыми, длинные перья хвоста — рулевыми, перья, покрывающие крылья сверху и снизу, — кроющими крыла, верхнюю часть хвоста — надхвостьем и т.д. Под контурными перьями находятся мелкие пуховые перья. Стержень их тонок, бородок второго порядка нет, поэтому опахала не образуют сомкнутых пластинок. Часто стержень пухового пера настолько укорочен, что бородки отходят от вершины одним пучком. Такое перо называют собственно пухом. Пуховые перья и пух особенно сильно развиты у водоплавающих и видов, обитающих в холодных странах. Основная их роль в уменьшении теплоотдачи.
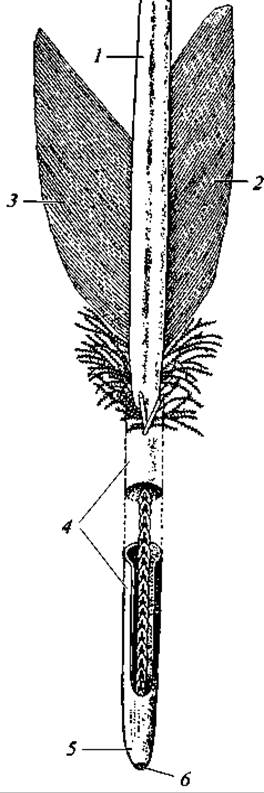
Рис. 127. Контурное перо (очин вскрыт, чтобы показать дужку пера):
1 — стержень; 2 — наружное опахало; 3 — внутреннее опахало; 4 — очин; 5 — отверстие очина; 6 — дужка пера
Среди пуха находятся чувствующие нитевидные перья, в углах рта у многих птиц имеются щетинки. У насекомоядных видов, ловящих добычу в воздухе, они образуют при раскрытом клюве как бы воронку, увеличивающую возможность ловли насекомых.
Закладка и начальные стадии развития перьев сходны с чешуями рептилий. Зачаток пера, как и зачаток роговой чешуи, представляет собой бугорок соединительнотканного слоя кожи, покрытый снаружи эпидермисом. По мере роста бугорок отклоняется назад, а основание его опускается в глубь кожи, образуя влагалище будущего пера и его сосочек, богатый кровью, через который происходит питание растущего пера. Эктодермальная часть зачатка, разрастаясь, дифференцируется на продольное утолщение — будущий стержень и два продольных киля этого утолщения, которые в последующем распадаются на бородки опахал. Вначале зачаток пера покрыт снаружи тонким роговым чехликом, который разрушается в последующем по мере формирования пера. После этого опахала освобождаются, и левая и правая их половинки раздвигаются в стороны (рис. 129). Перья регулярно сменяются. У многих птиц в году бывает не одна, а две или три линьки. В последнем случае меняется обычно не все оперение, а только определенные его участки. Многократная линька связана с наличием сезонного полиморфизма и брачным нарядом. Характер линьки у птиц различен. Хищные и насекомоядные, ловящие добычу в воздухе, линяют постепенно и не теряют способности к полету. Куриные, обитатели лесов, кустарниковых и травянистых зарослей линяют быстро. В это время они с трудом могут подняться в воздух и держатся в укромных местах, затаиваясь при приближении опасности в чаще кустов или травы. Очень своеобразно линяют утки, гуси, лебеди, чистики, поганки, гагары и большинство пастушковых. Маховые перья у них выпадают почти одновременно, и птицы на долгое время утрачивают возможность летать. Гуси, некоторые утки, лебеди в это время собираются в глухих, труднодоступных местах по берегам рек, озер и морей, концентрируясь здесь в огромном количестве, иногда помногу тысяч особей.
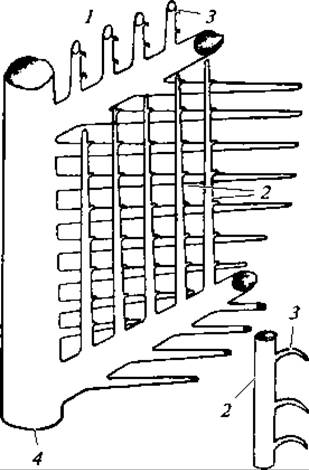
Рис. 128. Схема строения опахала:
1 — бородка; 2 — бородочки; 3 — крючочки; 4 — ствол
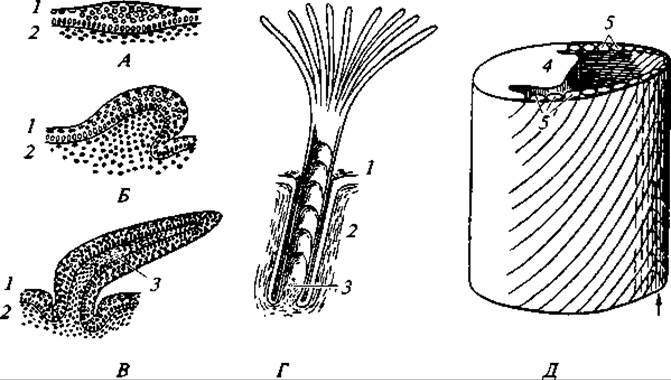
Рис. 129. Схема развития пера:
А, Б и В — продольные разрезы через зачатки пера разного возраста; Г — эмбриональное перо в разрезе; Д — стереограмма развивающегося контурного пера; 1 — эпидермис; 2 — кутис; 3 — сосочек пера; 4 — зачаток стержня; 5 — зачатки бородок. После сбрасывания наружной кожицы бородки опахала освобождаются и раздвигаются в обе стороны по линии, указанной стрелкой
При линьке происходит не только смена оперения, но меняется (у ряда видов) и его структура. Так, в летнем оперении чижа насчитывается примерно 1 500 перьев, а в зимнем — 2 100 — 2 400, у одного из видов синиц летом 1100 перьев, зимой — 1700. У белой куропатки длина контурных перьев на спине зимой равна в среднем 5,4 см, летом — 3,8 см; пуховая часть их соответственно равна 1,8 и 1,4 см; побочный ствол — 3,7 — 2,5 см (А. В. Михеев, 1960).
Мускулатура. Мускулатура птиц обладает рядом особенностей, связанных с основными чертами их жизнедеятельности. Во-первых, мышечная система более дифференцирована, чем у рептилий, что обусловлено сложными движениями при полете, хождении, лазании, добыче пищи. Во-вторых, наиболее крупные мышцы, приводящие в движение крылья, располагаются на туловище, а к самим конечностям идут сухожилия. Наконец, в-третьих, в связи с огромной работой, выполняемой крыльями, основная масса мускулатуры располагается не на спинной стороне тела, а на груди, где лежат мышцы, приводящие в движение крылья.
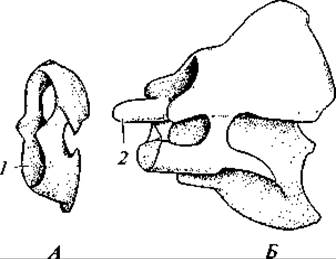
Рис. 130. Атлант (А) и эпистрофей (Б) гуся:
1 — сочленовная ямка для мыщелка черепа; 2 — зубовидный отросток
Особого внимания заслуживают мышцы конечностей. Огромные грудные мышцы, прикрепляющиеся к килю грудины и достигающие 20 % общей массы тела птицы, служат для опускания крыла. Лежащие под ними подключичные мышцы имеют несколько меньшие размеры и служат для поднятия крыльев. Очень сложную мускулатуру имеют задние конечности (до 35 мышц). Большой интерес представляет имеющаяся у некоторых видов обходящая мышца ноги. Она начинается на тазе, тянется вдоль бедра, далее в виде тяжа перекидывается через колено и затем соединяется со сгибателем пальцев. Птица, севшая на ветку, сгибает колени и этим движением натягивает обходящую мышцу, а следовательно, и сгибатель пальцев, в результате чего пальцы сжимаются, плотно обхватывая ветку. Чем ниже садится птица, тем сильнее натягивается обходящая мышца и тем сильнее птица обхватывает пальцами ветку. Спящая птица автоматически удерживается на ветке.
У других видов (например, у воробьиных) механизм автоматического сгибания пальцев иной. Он обусловлен действием мышцы, называемой глубоким сгибателем пальцев. Ее сухожилия, идущие к концам пальцев, имеют сильно шероховатую нижнюю поверхность и двигаются во влагалищах, внутренняя поверхность которых обладает поперечными ребрами. Когда птица садится на ветку и обхватывает ее пальцами, шероховатая поверхность сухожилий под тяжестью птицы прижимается к влагалищу и закрепляется на его ребрышках. В итоге пальцы фиксируются в согнутом положении и плотно обхватывают ветку без участия мышц.
Скелет. Скелет птиц имеет ряд специфических особенностей, связанных с приспособлениями к полету и хождению по суше только на задних конечностях. Особенности строения скелета заключаются, во-первых, в своеобразных видоизменениях конечностей и их поясов, во-вторых, в легкости и прочности всего скелета. Легкость его обеспечивается пневматичностью большинства костей, прочность — минерализацией и срастанием многих костей еще на ранних стадиях онтогенеза, поэтому у взрослой птицы швы между отдельными костями часто вовсе не заметны.
Позвоночник. Позвоночник включает шейный, грудной, поясничный, крестцовый и хвостовой отделы. Шейный отдел имеет значительную длину и высокий уровень подвижности. Подвижность обусловливается своеобразной формой позвонков, которые имеют седлообразные сочленовные поверхности. Такие позвонки называют гетероцельными.
Число шейных позвонков у разных видов птиц неодинаково и варьирует от 11 до 25. Два первых шейных позвонка имеют типичное для амниот строение, т. е. представлены атлантом и эпистрофеем (рис. 130). Шейный отдел участвует в сложных движениях головы. Угол поворота головы достигает обычно 180°, а у сов даже 270°.
Грудные позвонки (от 3 до 10) сращены между собой и с крестцом. Они несут ребра, подвижно сочлененные с грудиной. Ребро состоит из двух отделов, спинного и грудного, подвижно соединенных друг с другом и образующих угол, направленный вершиной назад. В связи с таким строением ребер грудина при сокращении соответствующих мышц может то отодвигаться, то приближаться к позвоночнику. Это изменяет объем грудной клетки и играет важную роль в механизме дыхания.
Своеобразное строение имеет грудина. Она представляет собой широкую, слегка выгнутую наружу костную пластинку, к краям которой причленяются грудные ребра. У современных летающих птиц грудина несет костный киль. К килю симметрично слева и справа прикреплены мышцы, приводящие в движение крылья. Только страусовые птицы (немногие виды, вторично утерявшие способность летать) не имеют киля, их грудина лишь слегка выпуклая. У пингвинов киль развит сильно, так как передние конечности у них выполняют большую работу при плавании.
Поясничные позвонки срастаются между собой, с подвздошными костями и с крестцовыми позвонками. С крестцовыми позвонками срастается часть хвостовых позвонков. В итоге образуется характерный для птиц сложный крестец, состоящий из большого числа позвонков (от 10 до 22). Однако истинных крестцовых позвонков у птиц, как и у рептилий, только два. Сложный крестец имеет важное приспособительное значение в связи с опорой тела птиц только на задние конечности при движении по земле. Свободных хвостовых позвонков у птиц 6 —9; хвостовой отдел заканчивается вертикальной костной пластинкой — копчиковой костью, или пигостилем, представляющим несколько сросшихся позвонков. Копчиковая кость служит опорой для прикрепления рулевых перьев.
Череп. Череп птиц по общей схеме строения близок к черепу рептилий. Затылочный отдел составлен обычными четырьмя затылочными костями (основной, двумя боковыми и верхней). Затылочный мыщелок, как и у рептилий, один. Слуховая капсула формируется, как и у рептилий, тремя ушными костями, которые у взрослых птиц сливаются в одну ушную кость.
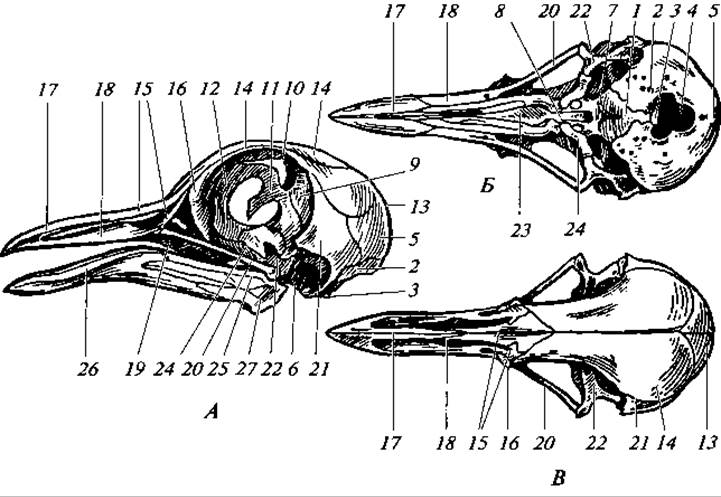
Рис. 131. Череп молодого голубя:
вид сбоку (А); снизу (Б); сверху (В); кости: 1 — основная затылочная; 2 — боковая затылочная; 3 — затылочный мыщелок; 4 — большое затылочное отверстие; 5 — верхнезатылочная; 6 — ушная; 7 — основная клиновидная; 8 — переднеклиновидная; 9 — крылоклиновидная; 10 — глазоклиновидная; 11 — межглазничная перегородка; 12 — средняя обонятельная; 13 — теменная; 14 — лобная; 15 — носовая; 16 — слезная; 17 — межчелюстная; 18 — верхнечелюстная; 19 — скуловая; 20 — квадратно-скуловая; 21 — чешуйчатая; 22 — квадратная; 23 — сошник; 24 — крыловидная; 25 — сочленовная; 26 — зубная; 27 — угловая
Дно черепа образовано основной клиновидной и переднеклиновидной костями, а также нёбными и крыловидными костями. Крышу черепа формируют парные носовые, лобные, теменные и чешуйчатые кости. Верхняя челюсть представлена межчелюстными и верхнечелюстными костями. К последним сзади причленяются палочковидные скуловые и квадратно-скуловые кости, которые, в свою очередь, соединяются с квадратными костями. В итоге образуется характерная для птиц нижняя височная дуга, отграничивающая глазницу и височную яму. Нижняя челюсть состоит из гомологичной меккелеву хрящу сочленовной кости и из кожного происхождения зубной, пластинчатой, угловой и венечной (надугловой) костей (рис. 131).
Подъязычный аппарат костный, в виде удлиненной пластинки и очень длинных рожков, гомологичных первой паре жаберных дуг, лежит в области гортани.
Слуховая косточка, как и у рептилий, одна (стремечко).
Наряду со сходством общей схемы строения череп птиц существенно отличается от черепа рептилий рядом особенностей приспособительного характера. Обращает внимание относительно крупный объем мозговой коробки и огромные глазничные впадины, что связано с хорошим развитием головного мозга (главным образом его полушарий) и очень крупными глазами, которые у птиц являются важнейшим органом чувств. Челюсти сильно вытянуты, и кости их срастаются между собой. Возникает весьма совершенный прочный хватательный аппарат. Кости мозговой коробки тонкие и срастаются очень рано; мозговая коробка легкая и прочная. Добавим к этому, что ряд костей черепа пневматичен.
Передние конечности и их пояс. В связи с приспособлениями к полету передние конечности и их пояс имеют ряд особенностей. Пояс состоит из лопатки, коракоида и ключицы, которые своими проксимальными концами формируют площадку для причленения плечевой кости. Лопатки длинные, саблевидно изогнутые, лежат на ребрах, по которым они могут свободно скользить. Крупный коракоид, развитый очень сильно, одним концом упирается в грудину, другой конец дает опору для причленения плеча. Левая и правая ключицы срастаются между собой, образуя характерную для птиц вилочку, придающую поясу особую упругость (рис. 132).
Скелет самого крыла состоит из всех типичных для пятипалой конечности отделов. При этом плечо и предплечье существенно не изменены, а кисть, наоборот, имеет ряд своеобразных особенностей. Запястье сильно редуцировано. Его проксимальные части срастаются в две косточки, а дистальные — с пястью. Пясть состоит из двух удлиненных костей, сращенных как в проксимальном, так и в дистальном отделе и образующих в итоге сложную пястнозапястную кость, пряжку. Основной сустав — межзапястный, или интеркарпальный. Из пальцев сохраняются только три — второй, третий и четвертый, при этом только третий палец имеет две фаланги, а второй и четвертый — по одной.
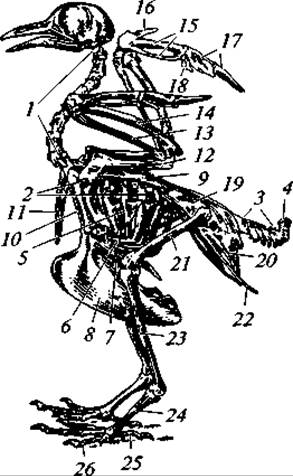
Рис. 132. Скелет голубя:
1 — шейные позвонки; 2 — грудные позвонки; 3 — хвостовые позвонки; 4 — копчиковая кость; 5 — спинная часть ребра с крючковидным отростком; 6 — брюшная часть ребра; 7— грудина; 8 — киль грудины; 9 — лопатка; 10 — коракоид; 11 — ключица; 12 — плечо; 13 — лучевая кость; 14 — локтевая кость; 15 — пястно-запястная кость; 16 — II палец; 17 — III палец; 18 — IV палец крыла; 19 — подвздошная кость; 20 — седалищная кость; 21 — бедро; 22 — лобковая кость; 23 — голень; 24 — цевка; 25 — I палец; 26 — IV палец ноги
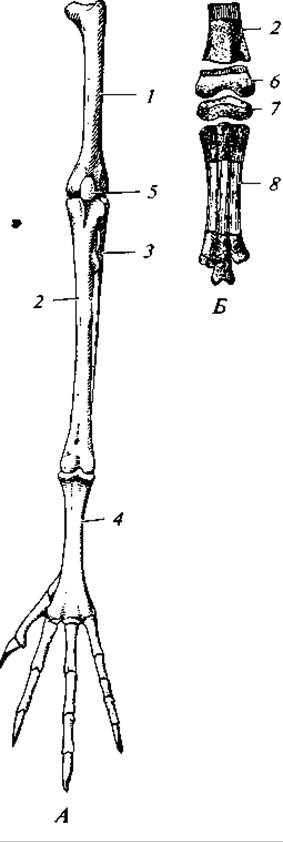
Рис. 133. Скелет задней конечности голубя (А) и часть задней конечности его птенца (Б):
1 — бедро; 2 — большая берцовая кость; 3 — редуцированная малая берцовая кость; 4 — цевка; 5 — чашечка; 6 — сросшиеся проксимальные элементы предплюсны, прирастающие позднее к большой берцовой кости; 7 — сросшиеся дистальные элементы предплюсны, прирастающие позднее к основаниям плюсневых костей; 8 — срастающиеся кости плюсны
В связи с наличием особых мускулов, сухожилий и кожистых перепонок кости крыла соединены между собой так, что все крыло не может быть вытянуто по прямой и его отделы остаются под некоторым углом друг к другу. Отделы конечности могут двигаться только в одном направлений — в плоскости крыла, складывая и расправляя его.
Задние конечности и их пояс. Задние конечности и их пояс также имеют ряд особенностей, связанных с тем, что при хождении вся тяжесть тела переносится на задние конечности. Прочность таза определяется срастанием больших подвздошных костей по всей их длине со сложным крестцом. С подвздошными костями срастаются также весьма крупные седалищные кости. Лобковые кости, наоборот, малы; в виде тонких палочек они присоединены к наружному краю седалищных. Все три кости таза принимают участие в образовании вертлужной впадины. На брюшной стороне левая и правая половины таза широко расставлены, что связано с откладыванием крупных яиц, покрытых твердой скорлупой.
Бедро типичного строения. Голень состоит из двух типичных костей, однако полностью развивается только большая берцовая кость, малая берцовая кость рудиментарна и прирастает к первой. К дистальной ее части прирастает проксимальный ряд косточек предплюсны. Срастание здесь столь полное, что у взрослой птицы швы утрачиваются полностью. Лежащий вслед за голенью отдел конечности носит название цевки. У взрослой птицы он состоит из одной длинной кости. Однако эмбрионально эта кость возникает в результате срастания костей плюсны и нижнего (дистального) ряда костей предплюсны. В итоге голеностопное сочленение у птиц (как и у пресмыкающихся) расположено между двумя рядами костей предплюсны, а не между предплюсной и голенью, как у амфибии. Это сочленение правильнее именовать межпредплюсневым, или интертарзальным (рис. 133).
Пальцев у птиц чаще всего бывает четыре, реже три и только в одном случае (африканский страус) — два.
Органы пищеварения. Все современные птицы не имеют зубов, и функцию их как орудия захвата и удерживания пищевых объектов выполняют роговые чехлы, одевающие верхнюю и нижнюю челюсти. Форма клюва сильно варьирует и находится в прямой зависимости от характера пищи и способов ее добывания. Так, у хищных птиц большая часть клюва покрыта толстым роговым чехлом; клюв у них крючкообразно изогнут. У зерноядных птиц клюв конический, приспособленный для расщепления твердых оболочек семян. У гусиных клюв уплощенный, с роговыми пластинками, выполняющими роль цедильного аппарата. У пеликанов между ветвями нижней челюсти находится большой кожистый мешок, служащий для зачерпывания рыбы.
К дну ротовой полости прикреплен язык, форма которого также разнообразна. Так, у хищных птиц он короткий и твердый, у гусиных — мясистый и уплощенный, у дятлов, вертишеек — очень длинный, тонкий. У питающихся цветочным нектаром птиц (колибри, цветочницы и др.) язык очень подвижен и сворачивается в трубочку, через которую они сосут нектар.
Слюнные железы развиты у птиц различно, а у некоторых (например, у козодоев) они почти отсутствуют. Слюна смачивает пищу, облегчая ее заглатывание. Особенно велика продукция слюнных желез у стрижей-саланганов, которые строят из затвердевающей на воздухе слюны свои гнезда, известные под неправильным названием «ласточкиных». У некоторых птиц в слюне присутствует фермент амилаза, поэтому переваривание углеводов начинается уже во рту.
Длинный пищевод у некоторых птиц (хищных, куриных, голубей) образует расширение — зоб. Он служит для временного пребывания в нем проглоченной пищи. У голубей стенки зоба в период выкармливания птенцов выделяют жирное творожистое вещество — так называемое «молочко» (содержит более 10 % белка и 12— 15 % жира), которым птицы кормят своих птенцов. Выделениями стенок пищевода кормят птенцов также трубконосые, фламинго и некоторые другие.
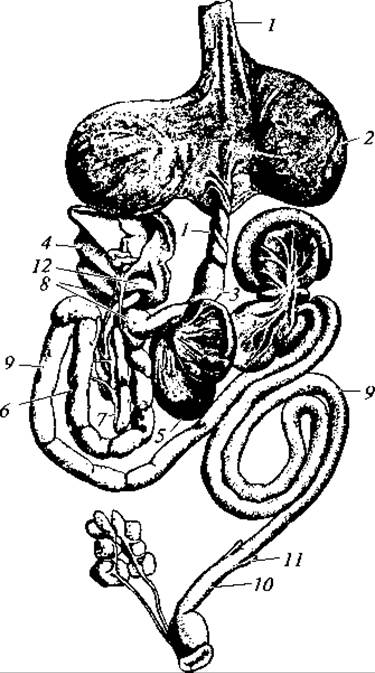
Рис. 134. Пищеварительная система голубя:
1 — пищевод; 2 — зоб; 3 — железистый желудок; 4 — печень; 5 — мускулистый желудок; 6 — двенадцатиперстная кишка; 7 — поджелудочная железа; 8 — желчные протоки; 9 — тонкая кишка; 10 — толстая кишка; 11 — слепые выросты кишки; 12 — селезенка
Пищевод ведет в тонкостенный железистый желудок, где пища подвергается воздействию секрета пищеварительных желез. Вслед за железистым желудком располагается толстостенный мускульный желудок, который изнутри выстлан плотной рогоподобной кутикулой. Здесь пища перетирается сокращением сильных мускульных стенок желудка (до 30 сокращений в 1 с) и камешками, которые находятся в полости желудка и играют роль жерновов. Развитость мускульного желудка связана с характером пищи. Меньше он развит у насекомоядных и хищных птиц, хорошо развит у зерноядных, наибольшего развития достигает у куриных, питающихся грубыми вегетативными частями растений (хвоей, концами веток, почками). У зерноядных птиц в мускульном желудке создается давление до 20 — 30 кг/см2. Скорость переваривания велика. Так, у домового воробья зерна перевариваются за 3 — 4 ч, жуки — за 1 ч, гусеницы — за 15 мин.
Тонкий отдел кишечника относительно длинный. В петле двенадцатиперстной кишки лежит поджелудочная железа. Хорошо отграниченный задний отдел кишечника сравнительно короток и недифференцирован на толстую и прямую кишку. На границе между толстым и тонким отделами кишечника у большинства птиц имеются два небольших слепых выроста. Короткая толстая кишка открывается в клоаку, на спинной стороне которой у многих птиц имеется слепой вырост — фабрициева сумка, играющая, видимо, роль железы внутренней секреции и участвующая в иммунологической защите организма. С возрастом она уменьшается в размерах.
Относительная длина кишечника различна и зависит от характера пищи. Так, у африканского страуса кишечник в 20 раз превышает длину тела, у баклана и коршуна — в 11 — 12 раз, у дневных хищников — в 7 — 8 раз, у большинства насекомоядных — в 4 — 7 раз.
Печень большая, двухлопастная. Желчный пузырь имеется у большинства видов (у голубя его нет). Желчный проток открывается в двенадцатиперстную кишку (рис. 134).
Таким образом, общие признаки пищеварительной системы птиц следующие: 1) отсутствие зубов, функционально замещенных отчасти роговым клювом (захват и удерживание пищи), отчасти мускульным отделом желудка (механическое перетирание пищи); 2) относительно небольшая длина кишечника. Так, у травоядных млекопитающих кишечник в 20 раз превышает длину тела, а у растительноядных птиц (у куриных) он длиннее тела всего в 8 раз; 3) дифференцировка желудка на два отдела, что связано с отсутствием способности измельчать пищу в ротовой полости и с необходимостью весьма интенсивного химического воздействия на пищу из-за укороченности кишечника; 4) отсутствие дифференцировки задней кишки на толстый и прямой отделы, видимо, надо рассматривать как приспособление к облегчению тела, так как функция прямой кишки в основе сводится к временному скапливанию фекальных масс и всасыванию из них воды; 5) наличие фабрициевой сумки — своеобразной железы внутренней секреции.
Органы дыхания. Органы дыхания крайне своеобразны и больше чем какая-либо другая система внутренних органов приспособлены к воздушному образу жизни.
Гортанная щель ведет в трахею, верхняя часть которой образует гортань, поддерживаемую непарным перстневидным хрящом и парными черпаловидными хрящами. Эта гортань у птиц известна под названием верхней и не играет роли голосового аппарата. Функцию такового выполняет так называемая нижняя гортань, свойственная только птицам. Она располагается в месте разделения трахеи на два бронха и представляет расширение, поддерживаемое хращевыми кольцами (рис. 135). Внутрь полости нижней гортани от ее стенок вдаются наружные голосовые перепонки, а снизу, от места ветвления трахеи, вдаются внутренние голосовые перепонки. Голосовые перепонки в связи с сокращением специальных певчих мышц могут менять положение и форму, чем и обусловливается разнообразие издаваемых ими звуков.
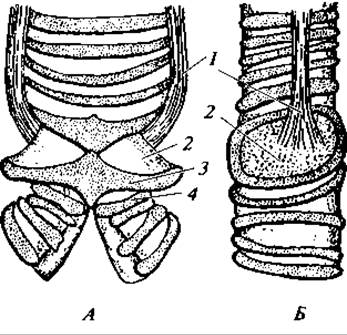
Рис. 135. Нижняя, или певчая, гортань голубя:
вид спереди (А) и сбоку (Б); 1 — бронхотрахейные мышцы; 2 — наружная голосовая перепонка; 3 — последнее кольцо трахеи; 4 — первое кольцо бронха
Верхние дыхательные пути имеют важное значение для терморегуляции. Установлено, что при повышении температуры внешней среды дыхание птиц резко учащается и становится поверхностным. Одновременно происходит очень сильное расширение кровеносных сосудов в полости рта и в глотке. Поэтому и возникает усиленная отдача тепла из организма птицы.
Легкие птиц представляют собой не полые мешки, как у амфибий и отчасти у рептилий, а плотные губчатые тела, прикрепленные к спинной стенке грудной клетки. Бронхи, войдя в легкие, многократно дихотомически ветвятся, и их главные разветвления пронизывают легкие насквозь и впадают в воздушные мешки (см. ниже). Ветки бронхов соединяются между собой тонкими каналами — третичными бронхами или парабронхами. Стенки парабронхов имеют небольшие углубления, оплетенные кровеносными капиллярами. Именно здесь происходит насыщение крови кислородом.
Часть разветвлений бронхов, как сказано, выходит за пределы собственно легких и расширяется в огромные тонкостенные воздушные мешки, объем которых примерно в 10 раз превосходит объем легких.
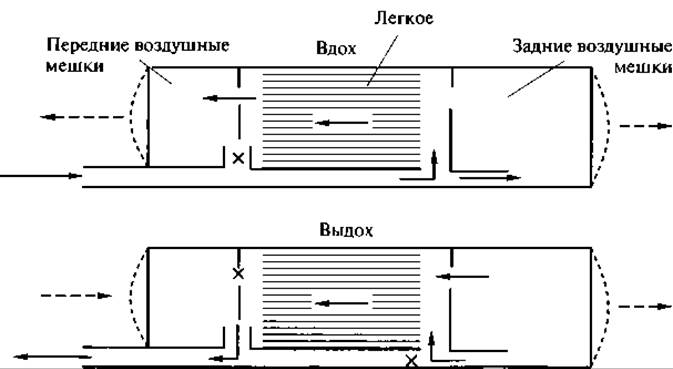
Рис. 136. Схема движения воздуха в дыхательной системе птиц (по К. Шмидт-Ниельсону, 1976).
Сплошными стрелками показано направление воздушного потока, пунктирными — расширение или сжатие воздушных мешков, значком х — места перекрытия потока воздуха при данной фазе дыхания.
Воздушные мешки располагаются между различными внутренними органами, а их ответвления проходят между мышцами под кожу и заходят в пневматичные кости. У птиц несколько воздушных мешков: два шейных, один межключичный, две-три пары грудных и одна пара очень крупных брюшных.
Значение воздушных мешков велико и разнообразно. Основная их роль заключается в том, что они обеспечивают механизм дыхания. Они растягиваются, и воздух по центральным бронхам с силой засасывается в задние воздушные мешки. При первом вдохе в задние воздушные мешки попадает богатый кислородом воздух. При первом выдохе из задних воздушных мешков в легкие поступает воздух с большим содержанием кислорода. На этой фазе дыхания происходит основное окисление крови.
При втором вдохе воздух из легких перемещается в передние воздушные мешки. При втором выдохе воздух из передних воздушных мешков выталкивается наружу. Таким образом, движение воздуха в дыхательной системе птиц происходит всегда в одном направлении (рис. 136): из задних воздушных мешков через легкие в передние воздушные мешки и наружу.
Кроме обеспечения акта дыхания (продвижения воздуха через легкие) воздушные мешки имеют и другие функции. Так, при усиленной работе мускулатуры во время полета они предохраняют его от перегревания, так как относительно холодный воздух «обтекает» практически все внутренние органы, а частично и мускулатуру. Воздушные мешки уменьшают во время полета трение между органами. Наконец, они уменьшают плотность тела, увеличивают внутрибрюшное давление и способствуют дефекации.
Частота дыхания у разных видов неодинаковая. У голубя в состоянии покоя число дыханий в одну минуту в среднем равно 26, при ходьбе — 77, в полете — 400. При этом легочная вентиляция в 2,5 раза превосходит потребность в метаболическом газообмене и служит для сброса избыточного тепла с легочным испарением. Следует учесть, что теплопродукция в полете в 8 раз больше, чем в состоянии покоя. Частота дыхания у мелких птиц, как правило, больше, чем у крупных: среднее число дыхательных движений в минуту у утки 30 — 43, у мелкой воробьиной птицы — 90 — 100.
Мелкие птицы обладают более интенсивным обменом веществ и потребляют значительно больше кислорода, чем крупные. Так, колибри с массой тела от 3 до 7 г употребляет от 4 до 10 мл кислорода за 1 ч на 1 г массы тела; кукша с массой тела 71 г потребляет 1,75 мл, голубь при массе тела 150 г — 0,98, страус эму при массе 38 кг — 0,023 мл. Указанные примеры подтверждают правило общей обратной зависимости размеров тела и интенсивности метаболизма у гомойотермных животных. Укажем для сравнения, что у филогенетически ниже стоящих рептилий этот показатель равен всего 0,1-0,3 (Л. Проссер, 1977).
Органы кровообращения. Основной особенностью кровообращения у птиц является полное разделение артериальной и венозной крови, что обусловливается строением их сердца и отводящей системы артериальных дуг (рис. 137).
Сердце четырехкамерное, состоящее из двух предсердий и двух желудочков. Относительные размеры сердца у птиц сравнительно с другими позвоночными велики, что связано с высокой интенсивностью обмена веществ, особенно во время полета. Так, сердечный индекс (масса сердца/масса тела, %) утки равен 0,6, а у сходного с ней по массе кролика — всего 0,2. Масса сердца у мелких птиц относительно большая, чем у крупных, что связано с более интенсивным обменом веществ (в свою очередь, это обусловлено тем, что у мелких животных менее выгодное соотношение между объемом тела и его поверхностью, отдающей тепло). Например, у снегиря при массе тела в 23 г относительная масса сердца составляет 1,3 %, у чечетки массой 13 г — 1,6 %, а у синицы-московки массой 8 г — 1,8 %.
Существует также зависимость между относительной массой сердца и энергичностью движений. У хорошего летуна сокола-чеглока масса сердца составляет в среднем 1,7 % от массы тела, у менее хорошего летуна пустельги — 1,2 %, а у плохого летуна сороки — всего около 0,9 %.
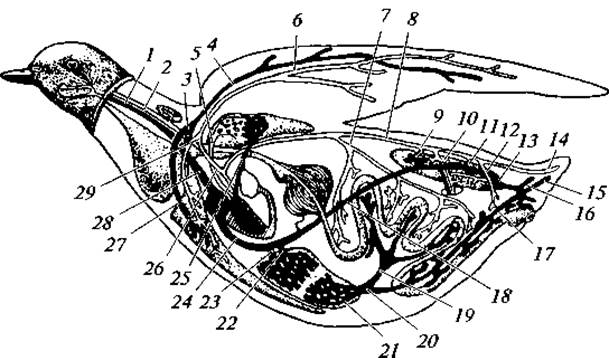
Рис. 137. Кровообращение голубя:
1 — яремная вена; 2 — сонная артерия; 3 — подключичные вены; 4 — плечевая вена; 5 — легочные вены; 6 — плечевая артерия; 7 — внутренностная артерия; 8 — спинная аорта; 9 — почечные артерии; 10 — седалищная артерия; 11 — почечная артерия; 12 — подвздошная артерия; 13 — внутренняя подвздошная вена; 14 — средняя копчиковая артерия; 15 — копчиковая вена; 16 — хвостовая вена; 17 — копчиково-брыжеечная вена; 18 — нижняя полая вена; 19 — мезентериальная вена; 20 — воротная вена печени; 21 — воротная система печени; 22 — правая печеночная вена; 23 — левая печеночная вена; 24 — нижняя полая вена; 25 — легочная артерия; 26 — трахейно-бронхиальная артерия; 27 — грудная вена; 28 — верхняя полая вена; 29 — подключичная артерия
Сердце у птиц работает более энергично, чем у низших наземных позвоночных. Так, у травяной лягушки число сокращений сердца в минуту 40 — 50, у снегиря — в среднем 730. Хорошо видна зависимость частоты сокращений сердца от размеров тела. У голубя массой 250 г число сокращений сердца в минуту в среднем 248, у зеленушки массой 22 г — 697, у щегла массой 13 г — 754, у московки массой 8 г — 1037. Число сердечных сокращений зависит и от состояния птицы. Так, у голубя в покое оно равно в среднем 165 ударам в одну минуту, а в полете — 550. При нырянии у птиц возникает брадикардия, т.е. снижение частоты сокращений сердечной мышцы. У оляпки через 5 с после погружения в воду сердечный ритм уменьшается от первоначального до 73 %, через 10 с — до 48 % и через 15 с — до 42 %.
Прогрессивной чертой птиц является и общее увеличение объема крови. У костистых рыб масса крови составляет примерно 3 % массы тела, у бесхвостых амфибий — 6 %, у птиц — 9 %. Кислородная емкость крови у птиц возрастает примерно в 2 раза в сравнении с рептилиями. Все эти черты организации служат важной предпосылкой повышения общего уровня обмена веществ птиц.
От левого желудочка отходит единственная правая дуга аорты, которая вскоре отдает парные безымянные артерии, в свою очередь, делящиеся на парные сонные и подключичные артерии. Так снабжаются кровью голова, пояс передних конечностей и сами конечности. Дуга аорты огибает сердце с правой стороны и в виде спинной аорты тянется под позвоночником к хвостовому отделу тела. От спинной аорты к желудку и кишечнику отходят непарные внутренностная и брыжеечная артерии. Парные бедренные и седалищные артерии снабжают кровью мышцы брюшной стенки, органы таза и задние конечности с их поясом.
От правого желудочка отходит общая легочная артерия, несущая венозную кровь. Она распадается на две короткие ветви — левую и правую легочные артерии.
Венозная кровь от головы собирается в парные яремные вены. Сливаясь с парными подключичными венами, они образуют левую и правую передние полые вены, изливающие кровь в правое предсердие. Венозная система туловищной области похожа на таковую у рептилий. Основные отличия в том, что брюшная вена, образующая у рептилий вместе с некоторыми другими, более мелкими, венами воротную систему печени, у птиц функционально заменена копчиковобрыжеечной веной. Кроме того, частично редуцирована воротная система почек. Задняя полая вена впадает в правое предсердие.
В связи с полным разделением большого и малого кругов кровообращения все органы снабжаются чистой артериальной кровью. Это обстоятельство, а также быстрая циркуляция крови и энергично идущий газообмен обусловливают высокую температуру тела птицы, в среднем равную 42°С. У крупных птиц она обычно 38 — 40°С, а у мелких достигает 45°С.
Подтверждением высокого уровня метаболизма у птиц служит и уровень артериального давления. Так, у голубя он равен 135/105, а у чешуйчатых рептилий — 80/60 — 14/10 мм рт. ст. (Л. Проссер, 1977).
Важное и принципиально новое физиологическое свойство птиц заключается в многообразии у них механизмов терморегуляции. Это касается как химической, так и физической терморегуляции. Первая состоит в изменении интенсивности обмена, т. е. величины теплопродукции в зависимости от температуры внешней среды, количества и качества потребляемой пищи. Так, падение внешней температуры с 33 до 10°С вызывает у воробья повышение потребления кислорода в 3 раза. Физическая терморегуляция заключается в изменении величины теплоотдачи. Существенное значение у птиц имеет так называемая тепловая одышка (или полипное): учащение дыхания приводит к увеличению отдачи тепла с выдыхаемым воздухом и испарению влаги из органов дыхания и дыхательных путей. Этим способом, например, мелкие птицы могут рассеять около половины накопленного в теле тепла. У крупных птиц рассеивание тепла таким образом может даже превосходить теплопродукцию. Благодаря этому, например, страусы и даже голуби могут выдерживать почти без перегревания температуру среды в 51°С.
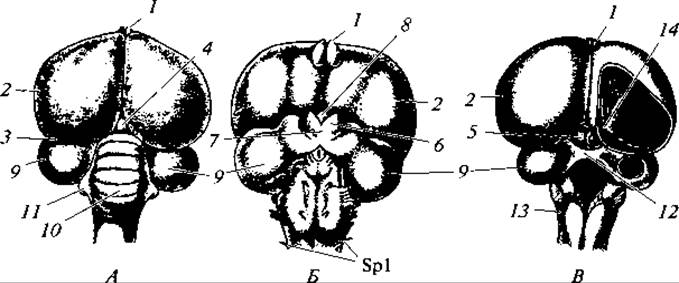
Рис. 138. Головной мозг голубя:
вид со спинной (А) и с брюшной сторон (Б), со вскрытыми желудочками и удаленным мозжечком (В); 1 — обонятельные доли переднего мозга; 2 — большие полушария переднего мозга; 3 — промежуточный мозг; 4 — надмозговая железа (эпифиз); 5 — зрительные бугры среднего мозга; 6 — зрительные тракты; 7 — хиазма; 8 — воронка с гипофизом; 9 — зрительные доли среднего мозга; 10 — мозжечок; 11 — боковые выступы мозжечка; 12 — зрительная комиссура; 13 — продолговатый мозг; 14 — полосатые тела; Spl — первая пара спинномозговых нервов
Нервная система. Центральная нервная система птиц сложнее, чем у рептилий. Это связано с более высоким уровнем жизненных отправлений и с более сложными взаимоотношениями птиц с условиями жизни. Нервно-рефлекторная деятельность и приспособительное поведение у птиц весьма разнообразны и сложны. Морфологически это обусловлено относительно крупными размерами головного мозга. У рептилий масса головного мозга примерно равна массе спинного, у птиц головной мозг всегда больше: у куриных в 1,5, у голубей в 2,5 раза. Его укрупнение обусловлено развитием полушарий переднего мозга (рис. 138). Так, отношение массы переднего мозга к массе всех других отделов головного мозга у куриных равно 1:1, у хищных — 2:1, у воробьиных и попугаев — даже 3:1. Однако, как и у рептилий, большая часть переднего мозга образована полосатыми телами, разрастанием архипаллиума в дне и стенках боковых желудочков. Наоборот, крыша полушарий в сравнении, к примеру, с млекопитающими развита относительно слабо и представлена тонким слоем нервных клеток. Обонятельные доли малы, что связано с недоразвитием органов обоняния. В промежуточном мозге на его верхней поверхности, прикрытой большими полушариями, находится эпифиз, а в дне, позади хиазмы зрительных нервов, — крупный гипофиз. Мозжечок очень большой, состоящий преимущественно из средней доли — червячка, испещренного характерными поперечными бороздами. Развитие мозжечка связано со сложными, требующими координации, движениями во время полета. Благодаря развитию мозжечка зрительные доли среднего мозга сильно отодвинуты в бока. Головных нервов одиннадцать пар.
Спинной мозг, как и у рептилий, имеет расширения в области плечевого и поясничного сплетений нервов.
Органы чувств. Орган слуха включает три отдела, как и у рептилий, есть внутреннее и среднее ухо. Канал улитки развит сильнее и отделен от мешочка перехватом. Евстахиевы трубы открываются в глотку. В среднем ухе — слуховая косточка одна. У птиц выражено наружное ухо. Барабанная перепонка лежит несколько вглубь от уровня кожных покровов, на дне небольшого воронкообразного углубления, представляющего собой наружный слуховой проход. У некоторых ночных птиц, например у сов, полость этого углубления большая и оторочена кожистой складкой, усиливающей тонкость слуха, а расположенные вокруг перья обеспечивают лучшее улавливание и направление звука. Птицы слышат очень хорошо, в диапазоне 30 Гц — 30 кГц, и орган слуха наряду с глазами является их важнейшим органом чувств. Зачастую об опасности птицы узнают в первую очередь именно при помощи слуха. Совы во время охоты ориентируются в основном при помощи слуха, определяя источник звука с расстояния 20 — 25 м с точностью до 1°, и почти без промаха ловят пробегающую мышь.
Установлено, что у некоторых птиц, например у яванского салангана (Callocalia fuciphaga), есть способность к эхолокации. Он издает звуковые импульсы с частотой 1,5 — 4,5 кГц с повторением 9 — 10 раз в секунду. Эта птица может летать в полной темноте в пещерах, успешно избегая столкновения с другими птицами и окружающими предметами. Полагают, что эхолокацией пользуются пингвины при охоте в воде, кроншнепы в тумане.
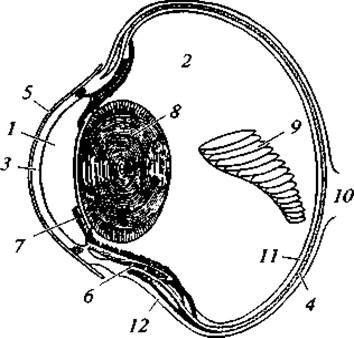
Рис. 139. Поперечный разрез глаза хищной птицы:
1 — передняя камера; 2 — задняя камера, заполненная стекловидным телом; 3 — роговица; 4 — сосудистая оболочка; 5 — соединительнотканная оболочка; 6— ресничная мышца; 7 — радужина; 8 — хрусталик; 9 — гребень; 10 — зрительный нерв; 11 — сетчатка; 12 — склера
Органы зрения у птиц развиты очень хорошо (рис. 139). В отличие от амфибий, рептилий и млекопитающих среди птиц нет видов с недоразвитыми глазами. Глазные яблоки относительно крупные, особенно у тех видов, которые деятельны в сумерках и ночью, либо у высматривающих добычу издалека. Отношение массы глазных яблок к общей массе тела у разных по биологии видов таково: гуси (кормятся травой) — 1/570; сороки (кормятся медленно передвигающейся добычей, насекомыми, которых разыскивают вблизи от себя) — 1/70; соколы (кормятся птицами, которых ловят чаще в полете, или насекомыми, высматриваемыми издалека) — 1/40; совы (кормятся ночью главным образом мелкими грызунами) — 1/30. У разных видов птиц на 1 мм2 сетчатки находится от 50 до 300 тыс. фоторецепторов — палочек и колбочек, а в области острого зрения — 500 тыс. — 1,5 млн. Разное сочетание палочек и колбочек у разных видов позволяет различать либо многие детали объекта, либо его контуры при малой освещенности. Наиболее чувствительное место на сетчатке глаза человека — желтое пятно — имеет 200 тыс. фоторецепторов. По этому показателю дневные хищные птицы по остроте зрения превосходят человека в 8 раз (рис. 140). Сокол-сапсан, например, реагирует на двигающуюся птицу, находящуюся на расстоянии до 1 100 м; сокол-дербник — до 800 м.

Рис. 140. Восприятие сетчаткой глаз изображения кролика человеком
(А) и ястребом (Б)
У большинства птиц зрение почти монокулярное. Поле зрения каждого глаза составляет примерно 150°, а поле бинокулярного зрения равно всего 30 — 50°. У сов зрение в большей мере бинокулярное, что обеспечивает определение точного расстояния до жертвы.
Аккомодация в глазе птицы достигается двумя способами: 1) изменением формы хрусталика под действием ресничной мышцы, 2) изменением расстояния между хрусталиком и сетчаткой, возникающим при действии кольцевых мышц, окружающих склеру и меняющих форму глазного яблока. Кроме верхнего и нижнего век имеется еще третье веко, или мигательная перепонка, прикрепленная основанием к внутреннему (переднему) краю глаза и способная, подобно шторе, затягивать весь глаз.
Обоняние развито слабо. Видимо, только некоторые птицы способны хорошо воспринимать запахи. Таковы новозеландские киви. Сравнительно с другими видами обоняние лучше развито у трубконосых, куликов, уток и грифов. Морфологически обонятельные органы птиц отличаются от таковых у рептилий большей дифференцировкой обонятельных раковин.
Органы выделения. Органы выделения устроены весьма сходно с органами выделения рептилий. У эмбрионов птиц закладываются туловищные почки, которые впоследствии замещаются, как и у прочих амниот, тазовыми почками.
Относительный размер почек у птиц больший, чем у рептилий и даже млекопитающих. Так, у скворца он составляет 1,2% массы тела, у крачек — 1,6, у крохаля — 2,6%. Укрупнение почек стоит в прямой связи с очень интенсивным общим обменом веществ у птиц. Об этом же свидетельствует и большое количество нефронов, исчисляемое десятками тысяч. В строении нефронов, как и у рептилий, характерна редукция сосудистого клубка; мочеотделение происходит в большей мере за счет секреторной деятельности извитых канальцев. Основным продуктом белкового обмена, как у рептилий, является мочевая кислота. Такой тип белкового метаболизма, несомненно, обусловлен, во-первых, наземным развитием в яйце эмбриона, при котором он практически полностью лишен возможности воспринимать воду из внешней среды, а во-вторых, необходимостью резервации продуктов метаболизма в системе замкнутого яйца. Естественно, что в такой ситуации не мог проявиться мочевинный тип обмена, при котором необходим очень большой расход воды при мочеотделении. Кроме того, мочевая кислота обладает наименьшей токсичностью, что позволяет этому продукту метаболизма сохраняться в системе яйца в течение всего периода развития. Таким образом, птицы дают еще один пример, когда в системе «закрытого» яйца возникает не мочевинный, как у рыб, амфибий и млекопитающих, а мочевокислотный тип обмена.
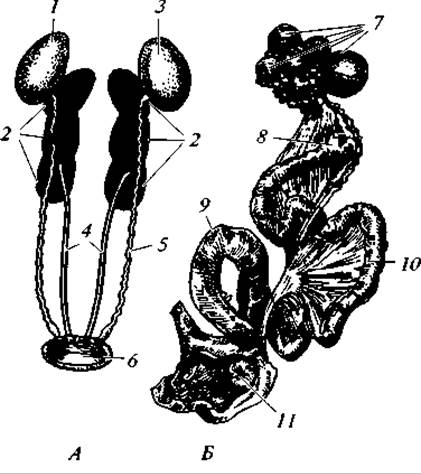
Рис. 141. Половые органы самца (А) и самки (Б) птиц:
1,3 — семенники; 2 — почечные доли; 4 — мочеточники; 5 — семяпровод; 6 — клоака; 7 — граафовы пузырьки; 8 — воронка яйцевода; 9 — матка; 10 — часть яйцевода, выделяющая белок; 11 — отверстие кишки
Моча проходит через мочевыводящие пути очень быстро, что связано, видимо, с плохой растворимостью мочевой кислоты и возможностью закупорки мочевыми солями проводящих путей. С этим же связано и отсутствие у птиц мочевого пузыря. Иногда приводимое в учебниках объяснение, связывающее отсутствие мочевого пузыря с облегчением массы тела, не имеет оснований. Потеря воды при мочеиспускании у птиц невелика, так как в клоаке происходит обратное всасывание воды из мочи в организм. Это обстоятельство наряду с отсутствием сколько-нибудь значительного кожного испарения обусловливает ничтожную потребность птиц в поглощении воды. Многие виды не пьют воду вовсе. Таково большинство хищных птиц, некоторые воробьиные.
Органы размножения. Семенники представляют собой пару бобовидных тел, расположенных над верхней долей почек и подвешенных на брыжейке (рис. 141). Величина семенников меняется по сезонам года. У зяблика, например, к периоду размножения объем семенников увеличивается в 1 125 раз, у скворца — в 1 500 раз. К внутренним краям семенников примыкают слабовыраженные придатки, гомологичные передним отделам туловищных почек эмбриона. От придатков отходят семяпроводы, которые тянутся параллельно мочеточникам и впадают в клоаку. У некоторых видов семяпроводы перед впадением в клоаку образуют расширения — семенные пузыри, служащие в качестве резервуара для семени.
Копулятивные органы есть только у немногих видов. Соответствующие выпячивания клоаки есть у гусиных, тинаму и страусовых. У цапель, аистов, дроф они рудиментарны. У большинства птиц совокупительных органов нет, и внутреннее оплодотворение достигается путем сближения отверстий клоак самца и самки, при котором сперма попадает в половые пути самки.
Репродуктивная система самок, за очень немногими исключениями, устроена резко асимметрично и состоит из левого яичника в левого яйцевода (см. рис. 141). Правый яичник развивается только иногда у дневных хищников, сов, гагар, попугаев, пастушков, куриных. Особенно часто он присутствует у попугаев. Однако функционирует правый яичник весьма редко. Любопытно, что в большинстве случаев яйцеклетки, созревшие в правом яичнике, выводятся через левый яйцевод. Причина редукции правой половины половых органов у самок птиц заключается, видимо, в откладывании относительно крупных яиц с жесткой скорлупой и длительном их перемещении. Яйцо движется по яйцеводу 1 — 2 суток.
Яичник птицы представляет собой зернистое тело неправильной формы, лежащее впереди от левой почки. Размеры яичника сильно варьируют в зависимости от зрелости и размеров формирующихся в нем яйцеклеток. Яйцевод имеет вид длинной трубки, один конец которой открывается воронкой в полость тела, другой (нижний) — в клоаку. Созревшая яйцеклетка, богатая желтком, попадает из полости тела в воронку яйцевода, где оплодотворяется и транспортируется по нему далее. Собственно яйцевод состоит из нескольких отделов. Начальный отдел почти по всей длине (кроме самой верхней части) очень богат железами, выделяющими белок, который покрывает проходящее яйцо толстым слоем. У курицы в этом отделе яйцо находится 3 — 6 ч. В следующем, более тонком, отделе яйцо покрывается двумя пергаментообразными подскорлуповыми оболочками. Далее следует так называемая матка — отдел, богатый железами, которые образуют известковую скорлупу яйца, окрашивающие ее пигменты и тонкую надскорлуповую оболочку. В матке яйцо находится 12 — 20 ч. Последний отдел яйцевода (влагалище) короток, обладает значительной мускулатурой; из него яйцо выходит в клоаку и затем наружу.
Весь период прохождения яйца по яйцеводу составляет у курицы около суток, у голубя — 41 час.
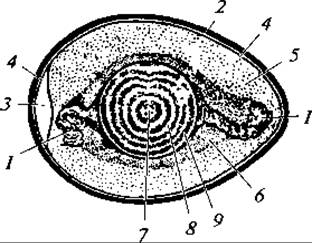
Рис. 142. Строение яйца птиц:
1 — халаза; 2 — скорлупа; 3 — воздушная камера; 4 — наружная подскорлуповая оболочка; 5 — жидкий белок; 6 — плотный белок; 7 — зародышевый диск; 8 — светлый желток; 9 — темный желток
Яйцо. Из приведенного описания видно, что собственно яйцеклеткой является лишь то, что в быту называют желтком, который покрыт первичной яйцевой оболочкой. После оплодотворения яйцеклетки образуется зигота, которая при движении яйца по яйцеводу начинает дробиться, формируя многоклеточный зародышевый диск. При откладывании яйца дробление клеток в зародышевом диске приостанавливается и возобновляется с началом насиживания. Остальные, дополняющие яйцо образования (например, белок) представляют собой оболочки, продуцируемые уже не яичником, а яйцеводом (рис. 142).
Желток состоит из мельчайших пузырьков — темного желтка, прослоенного яйцевой плазмой, или светлым желтком. Слои светлого желтка и темного желтка располагаются концентрически. В желтке накоплен запас питательных веществ и воды, которые расходуются развивающимся эмбрионом при его формировании. Состав желтка у разных видов варьирует, обращает внимание обилие в нем жиров и воды. При развитии зародыша яйца птиц не поглощают воду из наружной среды, подобно яйцам большинства рептилий, и весь запас воды, необходимый для развития, находится в самом яйце.
Жир в желтке при окислении дает воду. Куриный желток содержит воды 50 %, жиров — 23, протеинов — 16, липоидов — 11; в желтке гусиного яйца: воды — 44, жиров — 36, протеинов — 17, прочего — 3 %.
Белок представляет собой вторичную яйцевую оболочку полужидкой консистенции. Основное значение белка состоит в том, что он предохраняет собственно яйцо (т. е. желток) от механических повреждений, резких толчков и, кроме того, служит важным источником получения воды при развитии эмбриона. Состав куриного белка: вода — 87 %, протеины — 12, прочее — 1 %. Снаружи белок одет двумя тонкими оболочками, которые несколько расходятся на тупом конце яйца, образуя воздушную камеру, обеспечивающую возможность изменения объема яйца при перемене температуры. От скорлуповой оболочки у полюсов яйца в толще белка к желтку идут уплотненные слои белка, имеющие вид закрученных шнуров — халазы. Своими внутренними концами они прикрепляются к ободочке желтка. В итоге последний не плавает в белке свободно, а как бы растянут на хал азах, которые предохраняют желток от толчков. Кроме того, в связи с большой массой питательной части желтка его вегетативный полюс при любом положении яйца оказывается внизу, а зародышевый диск, наоборот, сверху. Халазы при этом скручиваются. Такое приспособление имеет большое значение для инкубации яиц, так как зародышевый диск всегда находится в наилучших условиях для обогревания.
Скорлупа состоит в основном из бикарбоната кальция (89 — 97 %) и включает незначительное количество других солей и органических веществ. Основное значение скорлупы состоит в защите яйца от механических повреждений, она не препятствует газообмену, но предотвращает проникновение бактерий. Как правило, яйца у птиц, гнездящихся открыто и имеющих крупные размеры, одеты более толстой скорлупой. Во время развития эмбриона скорлупа утончается, так как частично расходуется на построение скелета. Так, у курицы масса известковых элементов скорлупы за время насиживания понижается почти на 8 %, в то же время содержание извести в развивающемся эмбрионе на 20-е сутки насиживания увеличивается на 500 %.
Газообмен зародыша в яйце обеспечивается пористостью скорлупы. Число пор в скорлупе куриного яйца более 7 тыс., при этом они сосредоточены главным образом у тупого конца яйца. По мере насиживания и усиления газообмена просвет пор увеличивается.
Самой наружной оболочкой яйца является кутикулярный слой, формирующийся за счет белковых выделений, поступающих уже после прохождения яйца через матку. Функции кутикулы — предохранение яйца и зародыша от проникновения бактерий.