Зоология позвоночных - В. М. Константинов 2011
Хордовые
Наземные, или четвероногие, позвоночные - Позвоночные с зародышевыми оболочками
Класс Пресмыкающиеся, или рептилии - Подкласс Архозавры - Отряд Крокодилы
Среди современных рептилий крокодилы — наиболее высокоорганизованная группа, приспособленная к полуводному образу жизни.
Вытянутые туловище и голова сплюснуты сверху вниз. Хвост сжат с боков и служит основным орудием при плавании. Передние лапы с пятью свободными пальцами, задние лапы несут по четыре пальца, которые соединены перепонкой. Тело покрыто роговыми щитками, под которыми (чаще на спине) развиваются костные пластинки. В отличие от большинства рептилий крокодилы имеют кожные железы (правда, весьма немногочисленные), расположенные на хребте, под нижней челюстью и в области клоаки. Значение желез пока не вполне ясно.
Зубы расположены только на межчелюстных, верхнечелюстных и зубных костях и сидят в альвеолах, как у млекопитающих. В связи с разрастанием нёбных отростков межчелюстных и верхнечелюстных костей, а также нёбных костей развивается костный помост, известный под названием вторичного костного нёба, разделяющий полость рта на верхний отдел — носоглоточный ход и нижний отдел — собственно ротовую полость (рис. 111).
С такой перегородкой внутренние отверстия ноздрей открываются не непосредственно в ротовую полость, а в носоглоточный ход, который, в свою очередь, сообщается с глоткой; поэтому пища, находящаяся во рту, не мешает дыханию. Это, несомненно, прогрессивный признак, свойственный также млекопитающим.
Отделы позвоночника хорошо выражены. Шейный отдел содержит обычно девять позвонков, грудной — 12 — 13, поясничный — 2 — 4, крестцовый — 2 — 3, хвостовой — 30 — 40. Позвонки процельные. Плечевой пояс не имеет ключиц и состоит только из лопаток и коракоидов.
В связи с водным образом жизни есть своеобразные особенности в органах пищеварения и дыхания. Так, от заднего края нёба свешивается мускулистая складка, известная под названием нёбной завесы, которая своим нижним краем может примыкать к тыльной части языка. При таком положении нёбной завесы ротовая полость оказывается изолированной от глотки и крокодилы могут дышать, когда рот в воде раскрыт для принятия добычи и наружу выставлен только конец морды с ноздрями. У наружного края ноздрей располагаются клапаны, при погружении в воду всей головы они могут замыкать ноздри.
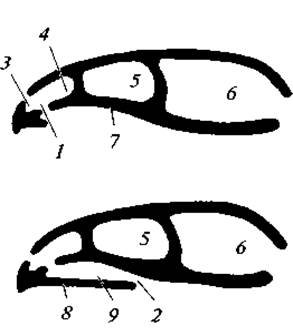
Рис. 111. Схема образования вторичного твердого костного нёба у пресмыкающихся: 1 — первичные хоаны; 2 — вторичные хоаны; 3 — дыхательная и 4 — чувствующая части обонятельной полости; 5 — орбита; 6 — черепная полость; 7 — первичное нёбо; 8 — вторичное твердое нёбо; 9 — носоглоточный проход
Легкие большие и имеют сложноячеистое строение, что связано с нахождением крокодилов под водой иногда в течение долгого времени.
В кровеносной системе характерно разделение желудочка на две изолированные половины. Следовательно, сердце крокодилов четырехкамерное (как у птиц и млекопитающих). Однако желудочки между собой сообщаются и наряду с правой дугой аорты, выходящей из левого (артериального) желудочка, имеется и левая дуга аорты, выходящая из правого (венозного) желудочка. Эти особенности способствуют смешению крови в спинной аорте (рис. 112).
В настоящее время эта немногочисленная группа насчитывает 25 видов, относящихся к трем семействам, распространенным в тропических и субтропических областях обоих полушарий.
Семейство Гавиалы (Gavialidae) включает один вид — Гавиал (Gavialis gangeticus). Он характеризуется чрезвычайно длинным рылом, которое на конце несколько вздуто (особенно у самцов). Длина тела до 6 м. Обитает на юге Индии. Наиболее многочисленно семейство Настоящие крокодилы (Crocodylidae) — 14 видов. Они имеют умеренно вытянутую морду. Распространены в Африке, Южной Азии, Северной Австралии и тропической Америке. Типичный вид — нильский крокодил (Crocodylus niloticus), имеющий длину тела до 8, а по некоторым данным, и до 10 м. Семейство Аллигаторы (Alligatoridae) включает 8 видов, в том числе китайский аллигатор (Alligator sinensis) — очень мелкий вид, длиной всего до 2 м (рис. 113). Распространен в Китае.
Крокодилы живут в воде, чаще в медленно текущих реках, озерах, многоводных болотах. Реже они встречаются в лагунах моря. На сушу выходят редко: для размножения, отдыха или при переселении из пересыхающего водоема в другой, более полноводный. Бывают случаи, когда крокодилы, не найдя в период засухи подходящего водоема, впадают в спячку, зарываясь в ил.
Размножаются откладыванием яиц. Яйца (несколько десятков) величиной с гусиные, покрыты крепкой известковой скорлупой.
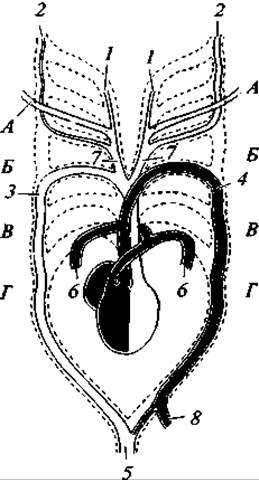
Рис. 112. Схема строения сердца и артериальных дуг у крокодила (с брюшной стороны):
А —Г— жаберные дуги (рыб); 1 — внутренняя сонная артерия; 2 — наружная сонная артерия (правая почти атрофирована); 3 — правая системная дуга аорты; 4 — левая системная дуга; 5 — спинная аорта; 6 — легочная артерия; 7 — безымянная артерия; 8 — желудочнокишечная артерия

Рис. 113. Крокодилы:
1 — гавиал; 2 — нильский крокодил; 3 — китайский аллигатор
Крокодилы откладывают их в вырытые на берегу ямки, зарывают их. У некоторых видов самка остается у гнезда и охраняет его от врагов. Период инкубации 1,5 — 2 месяца. Развитие эмбриона происходит при высокой температуре окружающей среды.
Питаются крокодилы разнообразными животными — от моллюсков и раков до птиц и сравнительно крупных млекопитающих. Известны случаи нападения на людей. Добычу чаще ловят в воде, медленно подкрадываясь к ней. Пасть при этом у крокодилов раскрыта, тело погружено в воду. Ловят птиц и зверей, находящихся на берегу. При охоте ориентируются главным образом с помощью слуха и зрения. Обоняние и осязание развиты слабо. В воде крокодилы очень проворны, плавают с большой ловкостью и быстротой. На суше они неуклюжи и медлительны.
Крокодилы служат предметом промысла, местами численность их заметно сократилась. Люди используют кожу и мясо этих животных. Во многих местах их промысел запрещен, создаются специальные фермы для искусственного разведения крокодилов. Почти все виды настоящих крокодилов включены в Красную книгу МСОП.
Происхождение и эволюция рептилий
Наземные позвоночные возникли в девоне. Это были панцирноголовые амфибии, или стегоцефалы. Они обитали близ водоемов и были тесно связаны с ними, так как размножались только в воде. Освоение отдаленных от водоемов пространств требовало существенной перестройки организации: приспособлений к защите тела от иссушения, совершенствования дыхания кислородом воздуха, хождения по твердому субстрату, способности размножаться вне воды и, конечно, усложнения форм поведения. Это основные предпосылки для возникновения качественных отличий, возникших у новой группы животных. Все эти черты оформились у рептилий.
К этому надо добавить, что к концу карбона произошли большие изменения природной обстановки, которые привели к возникновению на планете более разнообразного климата, развитию всевозможной растительности, ее распространению на отдаленных от водоемов территориях, а в этой связи и к широкому расселению трахейнодышащих членистоногих, возможных кормовых объектов рептилий, которые распространились и на водораздельные пространства суши.
Эволюция рептилий шла очень быстро и бурно. Еще задолго до конца пермского периода палеозоя они вытеснили большинство стегоцефалов. Получив возможность существовать на суше, рептилии в новой среде столкнулись с новыми и крайне разнообразными условиями. Многоплановость этого разнообразия и отсутствие на суше существенной конкуренции со стороны других животных послужили основными причинами расцвета рептилий в последующее время. Рептилии мезозоя — это прежде всего наземные животные. Многие из них вторично в той или иной мере приспособились к жизни в воде. Некоторые освоили воздушную среду. Приспособленность рептилий была поразительной. С полным основанием мезозой считают веком рептилий.
Ранние рептилии. Древнейшие рептилии известны из верхних пермских отложений Северной Америки, Западной Европы, России и Китая. Их называют котилозаврами. По ряду признаков они еще очень близки к стегоцефалам. Их череп представлял сплошную костную коробку с отверстиями только для глаз, ноздрей и теменного органа, шейный отдел позвоночника был слабо оформлен, крестец имел только один позвонок; в плечевом поясе сохранялся Клейтрум — кожная кость, свойственная рыбам; конечности были Коротки и широко расставлены.
Многочисленные остатки примитивных рептилий были найдены В. П. Амалицким в пермских отложениях Восточной Европы, на Северной Двине. Среди них — трехметровые растительноядные парейазавры (Pareiasaurus). Возможно, что котилозавры были потомками Каменноугольных стегоцефалов — эмболомеров.
Котилозавры достигли расцвета в средней перми. Но лишь немногие сохранились до конца перми, а в триасе эта группа исчезла, Уступив место более высокоорганизованным и специализированным группам рептилий, развивавшимся из различных отрядов котилозавров (рис. 114).

Рис. 114. Котилозавры (1, 2, 3) и псевдозухия (4):
1 — парейазавр (верхняя пермь), скелет; 2 — парейазавр, реставрация внешнею облика животного; 3 — сеймурия; 4 — псевдозухия
Дальнейшая эволюция рептилий сопровождалась изменчивостью их в связи с воздействием разнообразных условий жизни, с которыми они сталкивались при расселении. Большинство рептилий приобрели большую подвижность; скелет их стал легче и прочнее. Рептилии использовали более разнообразную пищу, чем амфибии. Изменялась техника ее добывания. В этой связи существенным изменениям подверглось строение конечностей, осевого скелета и черепа. У большинства конечности стали длиннее, таз приобрел большую устойчивость за счет прикрепления к двум и более крестцовым позвонкам. В плечевом поясе исчезла кость клейтрум. Сплошной панцирь черепа подвергся частичной редукции. В связи с большей дифференцировкой мускулатуры челюстного аппарата в височной области черепа возникли ямы и разделяющие их костные мосты — дуги, которые служили для прикрепления сложной системы мускулов.
Далее рассматриваются главнейшие группы рептилий, обзор которых должен показать исключительное разнообразие этих животных, их приспособительную специализацию и вероятное соотношение с ныне живущими группами.
В становлении облика древних рептилий и в оценке их последующей судьбы существенна характеристика их черепа. Примитивность стегоцефалов («цельночерепных») и ранних рептилий выражалась в структуре черепа: отсутствием в нем каких-либо впадин, кроме глазных и обонятельных. Эта особенность отразилась в названии Ąnapsida. Височная область у рептилий этой группы была закрыта кoстями. Вероятными потомками этого направления стали черепахи (ныне Testudines, или Chelonia), у них позади глазниц сохраняется сплошной костный покров. Предком черепах принято считать пермского эунотозавра (Eunotosaurus) — небольшое ящерообразное животное с короткими и очень широкими ребрами, образующими подобие брюшного щита (рис. 115). Спинного щита у него не было. Имелись зубы. Наиболее ранние черепахи известны из нижнего триаса мезозоя. Их ископаемые останки обнаружены на территории Германии. Череп, зубы, структура панциря древних черепах чрезвычайно близки современным.
Мезозойские черепахи первоначально были сухопутными и, видимо, роющими животными. Некоторые группы впоследствии перешли к водному образу жизни, и в связи с этим многие даже частично утратили костный и роговой панцирь.
С триаса и до наших дней черепахи сохранили основные черты своей организации. Они пережили все испытания, которые погубили большинство рептилий, и в настоящее время так же процветают, как и в мезозое.
Нынешние скрытошейные и бокошейные в большей мере хранят первичный облик сухопутных черепах триаса. Морские и мягкокожистые появились в позднем мезозое.
Все прочие рептилии, как древние, так и современные, приобрели в структуре черепа одну или две височные впадины. Одну, нижнюю, височную впадину имели Синапсидные. Одна верхняя височная впадина отмечена в двух группах: Парапсидные и Эвриапсидные. И наконец, две впадины имели Диапсидные.
Эволюционная судьба этих групп различна. Первыми от предкового ствола отошли Синапсиды (Synapsida) — рептилии с нижними височными впадинами, ограниченными скуловой, чешуйчатой и заглазничной костями. Уже в позднем карбоне эта группа из первых амниот стала самой многочисленной. В палеонтологической летописи они представлены двумя последовательно существовавшими отрядами: Пеликозавры (iPelicosauria) и Терапсиды (Therapsida). Их еще (менуют Звероподобные (Theromorpha). Период своего расцвета звероподобные пережили задолго до того, как появились первые динозавры.
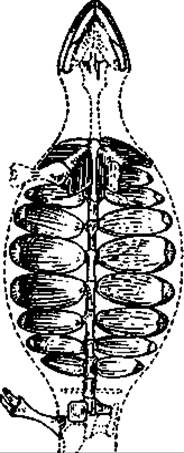
Рис. 115. Реконструкция скелета Eunotosaurus afriсanus — вероятного предка черепах (из пермских I отложений)
Их непосредственными родичами были котилозавры. В частности пеликозавры (Pelicosauria) были еще очень близки к котилозаврам Их останки нашли в Северной Америке и в Европе. Внешне они походили на ящериц и имели небольшие размеры (1—2 м), двояковогнутые позвонки и хорошо развитые брюшные ребра. Однако зубы их сидели в альвеолах. У некоторых намечалась, хотя и в небольшой степени, дифференцировка зубов.
В средней перми пеликозавров сменили более высокоорганизованные Зверозубые (Theriodontia). Зубы у них были явственно дифференцированы, появилось вторичное костное нёбо. Единый затылочный мыщелок разделился на два. Нижняя челюсть в основном была представлена зубной костью. Положение конечностей также изменилось. Локоть переместился назад, а колено — вперед, и в итоге конечности начали занимать положение под туловищем, а не по бокам его, как у других рептилий. В скелете появилось много черт, общих с таковым у млекопитающих.
Многочисленные пермские зверозубые рептилии были весьма разнообразны по облику и образу жизни. Многие были хищниками. Возможно, такой была найденная экспедицией В. П. Амалицкого в отложениях пермского периода на Северной Двине иностранцевия (Inostrancevia alexandrovi, рис. 116). Другие питались растительной или смешанной пищей. Эти малоспециализированные виды стоят ближе всего к млекопитающим. Среди них надо указать на циногнатуса (Cynognathus), имевшего много прогрессивных черт организации.
Зверозубые были многочисленны еще в раннем триасе, но при появлении хищных динозавров они исчезли. Любопытные материалы, приведенные в табл. 7, свидетельствуют о резком сокращении разнообразия звероподобных на протяжении триаса. Звероподобные представляют большой интерес как группа, давшая начало млекопитающим.
Следующей группой, отделившейся от анапсидных котилозавров, были Диапсидные (Diapsida). Их череп имеет две височные впадины, расположенные выше и ниже заглазничной кости. Диапсидные в конце палеозоя (пермь) дали чрезвычайно широкую адаптивную радиацию систематическим группам и видам, которых обнаруживают и среди вымерших форм, и среди нынешних рептилий. Среди диапсидных наметились две основные группы (инфраклассы): инфракласс Лепидозавроморфы (Lepidosauromorpha) и инфракласс Архозавроморфы (Archosauromorpha).
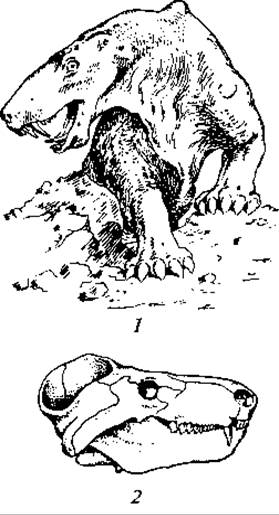
Рис. 116. Зверозубые:
1 — иностранцевия, верхняя пермь (реставрация животного); 2 — череп циногнатуса
Таблица 7
Соотношение родов звероподобных и завропсидных (ящероподобных рептилий) в конце палеозоя — начале мезозоя
(П. Робинсон, 1977)
Период |
Звероподобные |
Завропсидные |
Верхний триас |
17 |
8 |
Средний триас |
23 |
29 |
Нижний триас |
36 |
20 |
Верхняя пермь |
170 |
15 |
Палеонтологи не располагают точными сведениями, для того чтобы сказать, кто из них древнее и моложе по времени появления, но эволюционная судьба их различна.
Кто такие лепидозавроморфы? Этот древний инфракласс объединяет ныне живущих гаттерию, ящериц, змей, хамелеонов и их вымерших предков.
Гаттерия, или сфенодон (Sphenodon punctatus), живущая сейчас на небольших островах у побережья Новой Зеландии, — потомок достаточно распространенных в середине мезозоя первоящеров, или клинозубых (надотряд Prosauria, или Lepidontidae). Для них характерно множество клиновидных зубов, сидящих на костях челюстей и на нёбе, как у амфибий, и амфицельные позвонки.
Ящерицы, змеи и хамелеоны составляют ныне широкое многообразие отряда Чешуйчатые (Squamata). Ящерицы — одни из древнейших продвинутых групп рептилий, их останки известны из верхней Перми.
Ученые обнаруживают много черт сходства ящериц и сфенодонов. Их конечности широко расставлены и тело движется, волнообразно Изгибая позвоночный столб. Любопытно, что среди общих черт их Морфологического сходства — присутствие интертарзального сустава. Змеи появляются только в мелу. Хамелеоны — это специализированная группа более поздней эпохи — кайнозоя (палеоцен, миоцен). Архозавров считают самыми удивительными из всех рептилий, когда-либо живших на Земле. Среди них — крокодилы, птерозавры, динозавры. Крокодилы — единственные архозавры, дожившие до наших дней.

Рис. 117. Птерозавры
Крокодилы (Crocodylia) появляются в конце триаса. Юрские крокодилы существенно отличны от современных отсутствием настоящего костного нёба. Их внутренние ноздри открывались между нёбными костями. Позвонки были еще амфицельными. Крокодилы современного типа с полностью развитым вторичным костным нёбом и процельными позвонками произошли от древних архозавров — псевдозухий. Известны они с мела (около 200 млн лет назад). Большинство обитало в пресных водоемах, но среди юрских форм известны и настоящие морские виды.
Крылатые ящеры, или птерозавры (Pterosauria), представляют собой один из замечательных примеров специализации мезозойских рептилий. Это были летающие животные весьма своеобразного строения. Их крылья представляли складки кожи, натянутые между боками тела и очень длинным четвертым пальцем передних конечностей. Широкая грудина имела хорошо развитый киль, как у птиц; кости черепа срастались рано; многие кости были пневматичными. Вытянутые в клюв челюсти несли зубы. Длина хвоста и форма крыльев варьировали. У одних (рамфоринхи) были длинные узкие крылья и длинный хвост, летали они, видимо, скользящим полетом, часто планируя. У других {птеродактили) хвост был очень короткий, а крылья широкие; полет их чаще был гребной (рис. 117). Судя пo тому, что остатки птерозавров обнаружены в отложениях соленых водоемов, это были обитатели побережий. Они питались рыбой и по доведению, видимо, были близки к чайкам и крачкам. Размеры варьировали от нескольких сантиметров до метра и более.
Самые крупные среди летающих позвоночных принадлежат к позднемеловым крылатым ящерам. Это птеронодоны. Предполагаемый размах их крыльев 7 — 12 м, масса тела около 65 кг. Они обнаружены на всех континентах, кроме Антарктиды.
Палеонтологи предполагают постепенное угасание в эволюции этой группы, что совпало по времени с появлением птиц.
Динозавры (Dinosauria) известны в палеонтологической летописи с середины триаса. Это наиболее многочисленная и разнообразная группа рептилий, когда-либо живших на суше. Среди динозавров были мелкие животные с длиной тела менее метра, и гиганты длиной почти до 30 м. Одни из них ходили только на задних лапах, другие — на всех четырех. Очень разнообразен был и общий облик, но у всех голова относительно тела была мала, а спинной мозг в крестцовой области образовывал местное расширение, объем которого превосходил объем головного мозга (рис. 118).
В самом начале формирования динозавры разделились на две ветви, развитие которых шло параллельно. Характерной особенностью их было строение тазового пояса, в связи с чем эти группы именуются ящеротазовыми и птицетазовыми.
Ящеротазовые (Saurischia) первоначально были сравнительно мелкими хищными животными, передвигавшимися скачками только на задних ногах, в то время как передние служили для схватывания пищи. Для опоры служил и длинный хвост. В последующем появились крупные растительноядные формы, ходившие на всех четырех ногах. К ним относились самые крупные позвоночные, когда-либо жившие на суше: бронтозавр имел длину тела около 20 м, диплодок — до 26 м. Большинство гигантских ящеротазовых, видимо, были полуводными животными и питались сочной водной растительностью.
Птицетазовые (Ornithischia) получили свое название в связи с удлиненным тазом, похожим на таз птиц. Первоначально они передвигались на одних удлиненных задних ногах, но более поздние виды имели обе соразмерно развитые пары конечностей и ходили на четырех ногах. По характеру питания птицетазовые были исключительно растительноядными животными. Среди них — игуанодон, ходивший на задних ногах и достигавший в высоту 9 м. Трицератопс Внешне был очень похож на носорога, обладал обычно небольшим Рогом на конце морды и двумя длинными рогами над глазами. Длина его доходила до 8 м. Стегозавр отличался несоразмерно маленькой головой и двумя рядами высоких костных пластинок, располагавшихся на спине. Длина его тела была около 5 м.

Рис. 118. Динозавры:
1 — игуанодон; 2 — бронтозавр; 3 — диплодок; 4 — трицератопс; 5 — стегозавр; 6 — цератозавр
Динозавры были распространены почти по всему земному шару и обитали в крайне разнообразных условиях. Они населяли пустынилеса, болота. Некоторые вели полуводный образ жизни. Несомненно, что в мезозое эта группа рептилий была на суше господствующей. Наибольшего расцвета динозавры достигли в меловое время, а к концу этого периода вымерли.
Наконец, необходимо вспомнить еще об одной группе рептилий, в черепе которых имелась только одна верхняя височная впадина. Это было характерно для парапсидных и эвриапсидных. Высказано предположение, что они произошли от диапсидных путем утраты нижней впадины. В палеонтологической летописи они были представлены двумя группами: Ихтиозавры (Ichthyosauria) и Плезиозавры (Plesiosauria). В течение всего мезозоя, с раннего триаса до мела, они господствовали в морских биоценрзах. Как подмечено Р. Кэрролом (1993), пресмыкающиеся становились вторичноводными всякий раз, когда жизнь в воде оказывалась выгоднее с точки зрения наличия источников пищи и малого числа хищников.
Ихтиозавры (Ichthyosauria) занимали в мезозое то же место, которое ныне занимают китообразные. Они плавали, волнообразно изгибая тело, особенно хвостовую его часть, плавники у них служили для управления. Их конвергентное сходство с дельфинами поразительно: веретенообразное тело, вытянутое рыло и большой двухлопастный плавник (рис. 119). Парные конечности у них превратились власты, при этом задние конечности и таз были недоразвиты. Фаланги пальцев были удлинены, а число пальцев у некоторых доходило до 8. Кожа была голой. Размеры тела варьировали от 1 до 14 м. Ихтиозавры жили только в воде и питались рыбой, отчасти беспозвоночными. Установлено, что они были живородящими. Ихтиозавры появились в триасе, в конце мела они вымерли.
Плезиозавры (Plesiosauria) имели иные, чем ихтиозавры, приспособительные особенности в связи с жизнью в море: широкое и плоское тело со сравнительно слаборазвитым хвостом.
Орудием плавания служили мощные ласты. В отличие от ихтиозавров у них была хорошо развита шея, несущая небольшую голову. Их облик напоминал Ластоногих. Размеры тела от 50 см до 15 м. Отличен был и образ жизни. Во всяком случае, некоторые виды населяли прибрежные воды. Питались Рыбой и моллюсками. Появившись в начале триаса, плезиозавры, как и ихтиозавры, вымерли в конце мелового периода.

Рис. 119. Ихтиозавр и плезиозавр
Из приведенного краткого обзора филогении рептилий видно, что подавляющее большинство крупных систематических групп (отрядов) вымерли до начала кайнозойской эры и современные рептилии представляют собой лишь жалкие остатки богатейшей мезозойской фауны рептилий. Причина этого грандиозного явления понятна лишь в самых общих чертах. Большинство мезозойских рептилий представляли собой чрезвычайно специализированных животных. Успех их существования зависел от наличия своеобразных жизненных условий. Надо думать, что односторонняя глубокая специализация была одной из предпосылок их исчезновения.
Установлено, что хотя вымирание отдельных групп рептилий происходило в течение всего мезозоя, наиболее резко это проявилось в конце мелового периода. В это время за сравнительно короткий срок вымерло большинство мезозойских пресмыкающихся. Если справедливо называть мезозой веком рептилий, то не менее оправданно название конца этой эры веком великого вымирания. Следует принять во внимание, что в меловое время произошли значительные изменения климата и ландшафтов. Это совпало с существенными перераспределениями суши и моря и движениями земной коры, приведшими к громадным горообразовательным явлениям, известным в геологии под названием альпийской стадии горообразования. Считают, что в это время вблизи Земли проходило крупное космическое тело. Нарушения сложившихся условий жизни в этой связи были значительными. Однако они заключаются не только в изменении физического состояния Земли и других условий неживой природы.
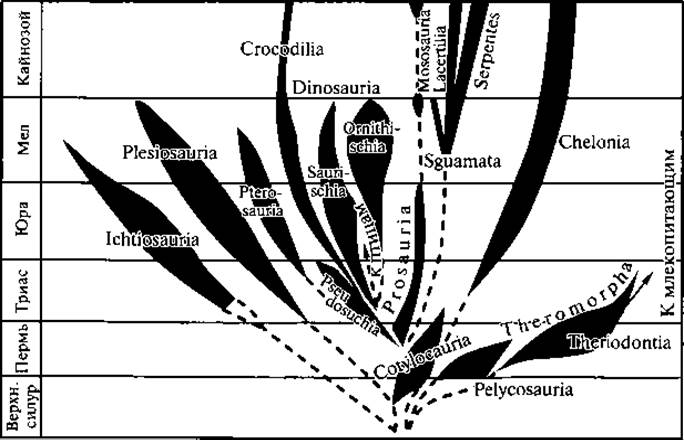
Рис. 120. Филогенетическое древо пресмыкающихся
В середине мелового периода произошла смена мезозойской флоры хвойных, саговниковых и других растений представителями флоры нового типа, а именно покрытосеменными. Не исключены и генетические изменения в природе самих рептилий. Естественно, что все это не могло не отразиться на успехе существования всех животных и специализированных в первую очередь.
Наконец, надо учесть, что к концу мезозоя все большее развитие получали несравненно более высокоорганизованные птицы и млекопитающие, сыгравшие важную роль в борьбе за существование между группами наземных животных.
На рис. 120 дана общая схема филогении рептилий.
Экология пресмыкающихся
Условия существования и общее распространение. По сравнению с условиями жизни земноводных условия жизни современных пресмыкающихся более разнообразны. Это связано с более высоким уровнем организации пресмыкающихся, обусловившим широкое их распространение в различных местообитаниях. Одной из наиболее важных в этом смысле особенностей является их способность переносить сухость наземной среды как во взрослом состоянии, так и в период эмбрионального развития. У взрослых животных это связано с изменением структуры кожи, в частности с ороговением эпидермиса. Эмбриональные приспособления будут описаны далее, в разделе о размножении.
Рептилии приспособлены к обитанию и в условиях теплого влажного климата, и в обстановке сухих жарких пустынь. Общее географическое распространение пресмыкающихся значительно расширилось в сравнении с амфибиями. Они заселяют все климатические зоны земного шара, за исключением арктической и антарктической. Однако в связи с пойкилотермностью рептилии успешно существуют только при относительно высокой температуре среды. Так, например, змеи уже при 10 °С становятся малоактивными, при 6 —8 °С они перестают двигаться, а при температуре 2, 3 °С впадают в оцепенение. Охлаждение тела до -4 - 6 °С вызывает гибель.
В результате рептилии наиболее многочисленны в тропическом поясе, а по направлению к полюсам обилие видов и особей уменьшается. Так, на островах Индонезии живет 150—200 видов, а в Среднем Китае всего лишь около 30. В Средней Азии обитает около 50 видов, на Кавказе — 68, в средней полосе Западной Европы — 12, наконец, до Северного полярного круга в западной (наиболее теплой) части Европы доходят только два вида (гадюка и живородящая ящерица). В горных странах по мере поднятия вверх над уровнем моря обилие рептилий также заметно сокращается. Верхний предел распространения рептилий в Центральной Азии лежит на высоте около 5 000 м над уровнем моря. В Кордильерах Южной Америки на высоте 4 900 м над уровнем моря встречается только один вид ящериц (Liolaemus multiformes).

Рис. 121. Степная агама
Вместе с тем чрезмерно высокая температура окружающей среды для рептилий губительна. Ящурка, находящаяся на песке, температура которого 55 °С, через 1,5 — 4 мин при невозможности куда-либо укрыться погибает. Спасаясь от перегревания, в наиболее жаркую часть дня ящерицы (агамы) прячутся в норы или залезают на ветки кустарников (рис. 121). Было прослежено, что в песчаных пустынях Средней Азии летом в середине дня температура воздуха на ветке на высоте 2 м над почвой была в тени на 10 °С ниже температуры почвы, а при прямом солнечном освещении этот разрыв достигал 28 °С (табл. 8).
Есть основания предполагать наличие у рептилий некоторых механизмов физической терморегуляции, но на очень низком уровне их проявления. Так, у живородящей ящерицы при подъеме температуры внешней среды от 16 до 42 °С частота дыхания увеличивалась от 31 до 114 раз в 1 минуту. Это явление можно трактовать как тепловую одышку, способствующую удалению из тела с дыханием избыточного тепла. Кроме того, установлено, что у американских сцинков и игуан весьма существенное значение для терморегуляции имеет потеря воды через кожу. Эти потери, исчисленные в процентах от общей водопотери организма, равняются 66 — 76% (Л. Проссер, 1977).
Таблица 8
Температура 2 июня на разной высоте от поверхности песка, °С, в пустынном районе Туркмении
Высота |
Температура |
||
в 10 ч |
в 12 ч |
в 14 ч |
|
На поверхности песка |
45 |
57 |
63 |
2 см, т.е. на уровне тела песчаной круглоголовки |
39 |
43 |
46 |
4 см, т.е. на уровне тела ушастой круглоголовки |
36 |
40 |
43 |
Высокая температура оказывает и косвенное отрицательное влияние на жизнедеятельность рептилий. Так, выгорание растительности служит причиной летней спячки степных черепах. В тропиках высокая температура и пересыхание водоемов вызывают спячку у крокодилов, некоторых черепах и змей.
Наличие в воде или почве солей не мешает в отличие от земноводных существованию пресмыкающихся. Многие виды ящериц и змей живут на сильно засоленных почвах, где обитание амфибий невозможно. Змеи, черепахи (в том числе даже лишенные рогового панциря) долгое время без вреда для организма проводят в соленой воде морей и озер.
Общее экологическое разнообразие пресмыкающихся исключительно велико. Среди них есть виды наземные, подземные, живущие на деревьях, полуводные, водные. Современные рептилии отсутствуют только в воздухе. Такое многообразие жизненных форм понятно, если учесть, что живущие ныне рептилии — остатки многих, далеко разошедшихся ветвей, представители которых издавна были приспособлены к обитанию в разных условиях.
Наибольшее число видов ведет наземный образ жизни, используя все разнообразие наземных условий. Они встречаются в песчаных, глинистых и каменистых пустынях, в зарослях густой травы, в лесу, на болотах и т.д. Вместе с тем большинство предпочитает открытые участки, где много солнца и растительность не мешает передвижению.
Характер передвижения рептилий различен (рис. 122), и только некоторые из них могут быть названы «пресмыкающимися» в полном смысле этого слова, т. е. животными, волочащими свое тело по земле. Крокодилы, вараны и многие ящерицы бегают, высоко приподняв тело на ногах. Некоторые сухопутные черепахи также не волочат брюхо по земле, а поднимают его. Есть виды, способные пробегать большое расстояние на одних только задних ногах. Таковы некоторые агамы, игуаны и особенно характерная в этом отношении австралийская плащеносная ящерица (Chlamydosaurus). Особенно много таких рептилий было среди вымерших мезозойских групп.
Способность пустынных рептилий бегать, высоко приподнимаясь на ногах, несомненно, имеет приспособительное значение, так как при таком положении тело не испытывает губительного воздействия раскаленного солнцем грунта. Это видно из данных табл. 8, составленной применительно к условиям Туркменистана.
Рептилии, живущие в песчаных пустынях, обладают рядом приспособлений для передвижения по сыпучему грунту: у агам, гекконов, ящериц, игуан на краях пальцев есть оторочки из роговых зубчиков. У одного из видов гекконов (Palmatogecko rangi), живущего в песчаных пустынях Южной Африки, между пальцами имеются кожистые перепонки.
Обширная и разнообразная по составу группа рептилий приспособлена к передвижению в густой траве, среди кустов и растительных остатков. С этим связано приобретение «змеевидной» формы тела, частичной или полной редукции конечностей. Наиболее совершенно эта черта организации развита у змей, большинство видов которых полностью утеряли не только свободные конечности, но и их пояса. Рудименты тазового пояса и задних конечностей имеются у удавов (Bod), слепунов (Typhlopidae), у вальковатых змей (Anilidae, старое название Ilisiidae). Но и среди ящериц есть полностью безногие. Такова, например, обычная в лесах России веретеница.
Очень многие пресмыкающиеся хотя бы часть времени проводят под землей, в которую закапываются сами, залезают в трещины в почве, в норы других животных. Приспособления к рытью различны. Сухопутные черепахи роют норы обеими парами ног (в норах они спасаются от дневной жары). Так же поступают и некоторые ящерицы. Но большинство, закапываясь, раздвигают грунт с помощью головы. Межчелюстной щиток многих роющих змей выдается вперед и служит как бы заступом.
К примеру, легко зарывается в песок и свободно ползает под его покровом песчаный удавчик.
Своеобразно закапываются в песок круглоголовки. По бокам их уплощенного тела располагаются кожистые оторочки, снабженные чешуйками. Спасаясь от врагов, животное плотно прижимается к земле и, быстро перемещая тело вправо и влево, разгребает песок, который засыпает его тело сверху. Круглоголовка утопает в песке, опускаясь в него не под углом (как это делают змеи и другие ящерицы), а отвесно вниз. Подобные приспособления есть и у некоторых змей (рогатая гадюка, эфа).
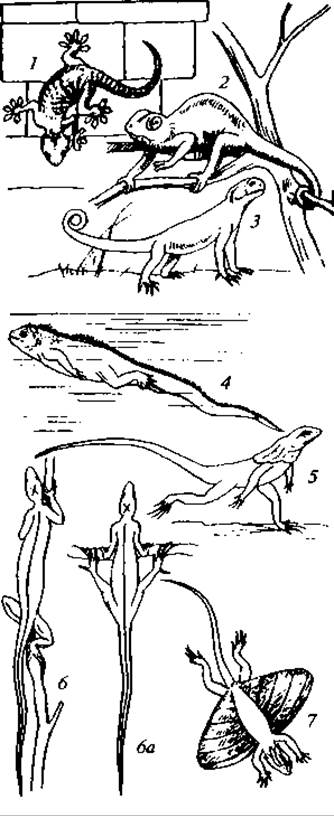
Рис. 122. Различные формы движения у ящериц:
1 — степной геккон; 2 — лопастеносный хамелеон; 3 — ушастая круглоголовка; 4 — морская ящерица; 5 — плащеносная ящерица; 6, 6а — игуана; 7 — летучий дракон
Среди рептилий есть много видов, хорошо лазающих по отвесным плитам камней, кустам и деревьям. Эта особенность свойственна разным систематическим группам и связана с такими приспособлениями. У некоторых ящериц сильно развиты пальцы с длинными изогнутыми когтями, длинный необламывающийся хвост, который может закручиваться вокруг ветвей и служит дополнительной точкой опоры. Интересно приспособление к удержанию на ветвях, свойственное хамелеонам: пальцы их ног срастаются так, что животные обхватывают ими ветки, как клещами. Среди ящериц хорошо лазают по кустам агамы. Есть лазающие виды и среди змей.
Немногим пресмыкающимся свойственна способность к планирующему полету, развившаяся в связи с древесным образом жизни. У живущего в лесах Зондских островов лопатохвостого геккона (Ptychozoon) по бокам головы, тела, хвоста и между пальцами имеются кожистые перепонки, служащие парашютом. Еще более оригинальное приспособление имеет летающий дракон (Draco draco) — обитатель лесов Малайского архипелага. Парашютом служит широкая складка кожи, находящаяся по бокам тела и растягивающаяся при прыжке 5 — 6 парами ребер. Длина прыжка достигает 20 м. Эта ящерица нередко на лету ловит насекомых.
Значительное число рептилий ведет в той или иной мере водный образ жизни. Крокодилы плавают при помощи сжатого с боков хвоста; лишь отчасти плаванию им помогают лапы. Живущая на Галапагосских островах морская игуана (Amblyrhynchus) плавает только при помощи хвоста, а лапы держит плотно прижатыми к туловищу. Изгибая хвостовую часть тела (также сжатую с боков), плавают морские змеи. Иное приспособление имеют водные черепахи: орудием плавания у них служат ластообразные конечности.
Наряду с основными органами передвижения (плавания) у водных рептилий имеются и другие приспособления к жизни в воде. Напомним о клапанах, закрывающих ноздри. У настоящих водных черепах в той или иной мере редуцируется костный и роговой панцирь. Изменены органы дыхания. Легкие имеют сложное строение и обеспечивают лучший газообмен при больших интервалах между вдохами. К примеру, игуанам свойственно замедление кровотока и даже остановка сердца при нырянии. Кроме того, у черепах и водных змей в глотке появляются вздутые участки, богатые кровеносными сосудами и являющиеся своего рода внутренними «жабрами».
Питание. Рептилии питаются самыми различными животными — от мелких наземных и водных беспозвоночных до крупных млекопитающих. Ряд видов едят растительную пищу, но все же большинство добывают животные корма. Так, ящерицы ловят наземных насекомых, червей и моллюсков. Кроме этого, некоторые из них (агамы, игуаны) охотно поедают и растительную пищу. Морские черепахи и морские змеи обычно питаются рыбами. Ужи отлавливают много амфибий. Некоторые морские змеи, например пеламида, кормятся почти исключительно головоногими моллюсками. Крупные ящерицы, например вараны, и многие наземные змеи добывают мелких зверьков, птиц и пресмыкающихся. Крокодилы и крупные змеи едят не только мелких животных, но и крупных зверей, например молодых оленей. Известны случаи нападения их на людей.
Настоящими растительноядными рептилиями являются наземные черепахи (например, среднеазиатская черепаха). Лишь иногда они поедают мелких животных. Большинство рептилий пьют воду, и только виды, обитающие в пустынях, получают ее из съеденной пищи.
Способы отыскивания и добывания пищи различны. Ящерицы и некоторые змеи, разыскивая еду, много бродяжничают. Добычу они ловят как на поверхности, так и под землей. Например, сетчатая ящурка (Eremias grammied) в пустынях Средней Азии выкапывает из песка насекомых, зарывающихся на глубину до 10 см. Сцинки, роющиеся в песке на глубине нескольких сантиметров, легко обнаруживают пищу, находящуюся на поверхности, и как бы выныривают из песка, чтобы схватить ее. Некоторые змеи (например, удавы), крокодилы и водные черепахи подкарауливают добычу и бросаются на нее. Караулят, сидя на ветвях, свою добычу и хамелеоны.
Почти все рептилии пищу глотают целиком, не разделяя ее на части и не подвергая во рту измельчению. Черепахи и крокодилы отделяют куски пищи. У крокодилов имеется вторичное костное нёбо, позволяющее задерживать пищевой комок во рту без помехи для дыхания. Большинство рептилий пожирают свою добычу, предварительно не умерщвляя ее. Ядовитые змеи и удавы глотают уже убитую жертву. Обычно змеи, заглатывая жертву, как бы наползают на ее тело.
Размножение. Размножение рептилий существенно отличается от размножения амфибий и имеет много особенностей, приспособленных к наземному существованию. Оплодотворение всегда внутреннее, и самцы имеют копулятивные органы. Яйца развиваются вне воды. Развитие проходит без превращения, т.е. в отличие от амфибий личиночной стадии не бывает, и вылупившаяся из яйца молодь живет в тех же условиях, что и взрослые.
Основные приспособления к размножению на суше заключаются в особенностях строения и развития яйца. Рептилии имеют сравнительно с амфибиями крупные яйца. Укрупнение яиц идет за счет обогащения их желтком, а у некоторых (черепах и крокодилов) и за счет появления «белка». Большой запас питательных веществ в яйце обусловливает возможность прямого (без превращения) развития эмбрионов. В связи с откладыванием яиц на суше возник ряд приспособлений, защищающих их от иссушения, механического повреждения и обеспечивающих развивающемуся эмбриону возможность газообмена, снабжения его водой и выведения (хотя бы частично) продуктов распада. Приспособления эти выражаются в основном в появлении ряда оболочек, покрывающих яйцо.
Эмбриональному развитию рептилий сопутствуют два типа оболочек: яйцевые и зародышевые. Яйцевые оболочки формируются при прохождении яйца по яйцеводу после того, как произошло его оплодотворение. Яйца всех рептилий имеют прочную волокнистую оболочку, предохраняющую их от высыхания, растекания, механических повреждений и от проникновения бактерий; скорлуповая оболочка возникает в результате пропитывания волокнистой оболочки солями кальция.
У черепах и крокодилов, кроме того, имеется белковая оболочка (подобная белку птичьего яйца), которая является основным местом накопления в яйце воды, обеспечивающей развитие зародыша (рис. 123). У прочих рептилий необходимая для нормального развития эмбриона вода высвобождается в результате окисления жира, весьма обильного в желтке, — так называемая метаболическая вода; часть воды проникает извне через наружные оболочки яйца. Укажем для сравнения, что в то время, как у амфибий доля жира в яйце составляет 8 — 11 %, у рептилий она достигает 40 % и более. Волокнистая, скорлуповая и белковая оболочки продуцируются соответствующими отделами яйцеводов.
В яйце рептилий (как и прочих амниот) по мере развития зародыша за счет превращений в закладывающихся зародышевых листках возникает еще ряд оболочек, имеющих исключительно важное приспособительное значение; таковы амнион, сероза и аллантоис (рис. 124). Их называют зародышевыми оболочками.
На ранних стадиях развития зародыш погружается в желток. В результате разрастания эктодермы возникают складки, известные под названием амниотических. Они растут навстречу друг другу и в итоге замыкают зародыш. Возникающий таким путем амнион представляет собой сплошной замкнутый мешок, заполненный жидкостью, внутри которого находится зародыш. Таким образом, зародыш как бы погружается в маленький «водоем» — относительно постоянную среду. Роль амниона заключается еще и в том, что он предохраняет зародыш от соприкосновения с твердыми яйцевыми оболочками.
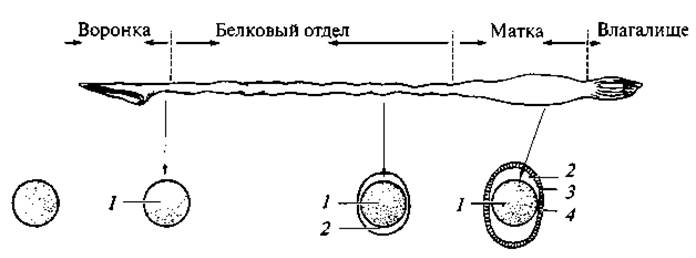
Рис. 123. Схема развития яйцевых оболочек у среднеазиатской черепахи (Testudo horsfieldi) в различных отделах яйцевода:
1 — оплодотворенная яйцеклетка; 2 — белковая оболочка; 3 — волокнистая оболочка; 4 — скорлуповая оболочка
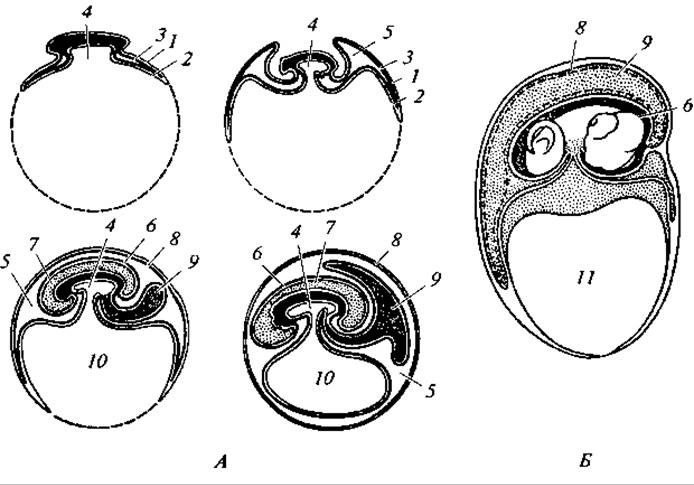
Рис. 124. Последовательные стадии развития зародышевых оболочек у амниот (А) и развивающаяся ящерица (Б):
1 — эктодерма; 2 — энтодерма; 3 — мезодерма; 4 — полость кишечника; 5 — внезародышевая полость тела; 6 — амнион; 7 — амниотическая полость, заполненная амниотической жидкостью, в которую погружен зародыш; 8 — сероза; 9 — аллантоис; 10 — желточный мешок; 11 — желток
При наличии оболочек, отделяющих зародыш от наружной среды, естественно возникает вопрос, как дышит зародыш и как выводятся наружу продукты обмена. Первоначальным приспособлением для дыхания служит густая сеть кровеносных сосудов, оплетающих желток, — желточный круг кровообращения. В дальнейшем он постепенно замещается новым образованием — сетью сосудов, развивающихся в аллантоисе. Аллантоис возникает как мешковидный вырост брюшной стенки задней кишки. По мере развития он приобретает вид большого пузыря, лежащего между желточным мешком и серозой, третьей зародышевой оболочкой. К концу развития аллантоис полностью окружает тело зародыша и желточный мешок.
Аллантоис выполняет одновременно две функции: он является органом дыхания и зародышевым мочевым пузырем. При этом аллантоис не только является пассивным приемником мочи, но и воспринимает из мочи излишнюю воду, которая через сеть кровеносных сосудов вновь вводится в тело зародыша. Кроме того, часть продуктов обмена зародыша выводится наружу в газообразном состоянии, в частности углекислый газ, так как основным энергетическим запасом в яйце рептилий служит жир, окисляющийся до воды и СO2.
Все описанные особенности строения яйца и развития эмбриона рептилий носят явно выраженный приспособительный характер: 1) предохраняют яйца от растекания, механических повреждений, иссушения (волокнистая и известковая оболочки); 2) создают возможность прямого развития (обилие питательного материала — желтка); 3) не препятствуют поступлению воды в яйцо из наружной среды (гигроскопичность яйца и пористость оболочек); 4) обеспечивают запас воды в самом яйце (жир желтка, а у некоторых — богатый водой белок); 5) создают относительно постоянные условия для эмбриона (жидкость амниотической полости); 6) обеспечивают возможность дыхания и выведения газообразных продуктов обмена из яйца, покрытого рядом оболочек (аллантоис). Эти особенности эмбрионального развития имели решающее значение при освоении рептилиями наземной среды обитания.
Большинство пресмыкающихся размножаются путем откладывания оплодотворенных яиц чаще всего в специально сооружаемую в земле ямку в сравнительно влажных и хорошо обогреваемых солнцем местах. Есть виды, откладывающие яйца в кучи растительного мусора, в гнилые пни. Американские крокодилы, к примеру, вырыв в болотистой почве ямку, засыпают яйца гниющими растениями. Морские черепахи выкапывают ямы в песке и затем засыпают отложенную кладку.
Как правило, отложив яйца, самки покидают их. Но некоторые крокодилы остаются вблизи «гнезда» и охраняют яйца от других животных. Самки питонов обвиваются вокруг яиц своим телом, обеспечивая таким образом не только сохранность яиц, но и относительно постоянную температуру. В таком гнезде она на 2 — 4 °С выше температуры окружающей среды. Охраняет яйца и самка варана.
Наряду с откладыванием яиц у некоторых рептилий существует яйцеживорождение. В простейшем случае оплодотворенные яйца остаются в половых путях и проходят там все стадии развития. Детеныши вылупляются из яиц тотчас же после откладывания их во внешнюю среду. Так бывает у гадюки, живородящей ящерицы, веретеницы.
Любопытно, что у ужей срок развития яиц вне тела матери варьирует от 30 до 60 суток: это зависит от того, сколько времени оплодотворенные яйца находились в теле матери. В отложенных яйцах зародыши оказываются сформированными примерно наполовину. Большая разница в длине периода внутриутробного нахождения яиц наблюдалась и у прыткой ящерицы (15 — 20 суток), при этом зародыши оказывались развитыми на 1/3 и более. Таким образом, предпосылкой для развития яйцеживорождения служит возможность более или менее длительной задержки оплодотворенных яиц в теле матери. Кроме указанных видов яйцеживорождение свойственно некоторым удавам, в частности песчаному удавчику, морским змеям, многим ужам, ящерицам.
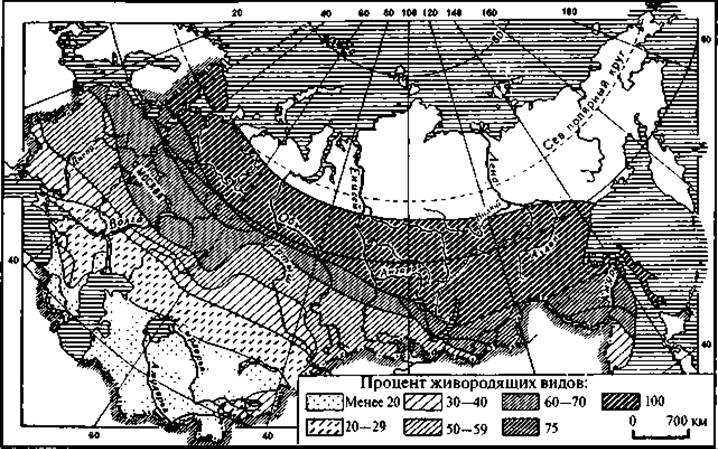
Рис. 125. Распределение живородящих рептилий по территории СНГ
Немногим видам свойственно настоящее живорождение, при котором кровеносные сосуды желточного мешка тесно сближены с сосудами маточного отдела яйцевода, и питание эмбриона идет в значительной степени за счет организма матери. Таковы некоторые сцинки.
Наконец, у некоторых змей и ящериц формируется настоящая аллантоидная плацента, при которой ворсинки хориоаллантоиса зародыша внедряются в слизистую оболочку половых путей самки.
В явлении живорождения (включая и яйцеживорождение) существуют довольно ясно выраженные географические закономерности. Установлено, что наибольшее число живородящих видов характерно для северных широт и высокогорных областей (рис. 125). Учеными обнаружены виды, которые в разных частях ареалов бывают то яйцеродными, то живородящими. Так, тибетская круглоголовка (Phrynocephalus theobałdi) на высоте 2 — 3 тыс. м размножается откладыванием яиц, а на высоте 4 — 5 тыс. м она живородяща. Установлено, что живородящая ящерица (Lacerta vivipard) на юге ареала (во Франции) откладывает яйца.
Очевидно, что основной причиной живорождения у рептилий является холодный климат. При нахождении яиц в теле матери обеспечивается более постоянная и высокая, чем в окружающей среде, температура, необходимая для их успешного развития. В отличие от млекопитающих, обогревающих своих эмбрионов постоянным теплом собственного тела, у рептилий это достигается тем, что самка своим поведением, перемещаясь в наиболее хорошо обогреваемые места, регулирует термические условия развития зародышей (например, гелиотермия). Дополнительной причиной живорождения, проявляющейся у немногих видов, служит водный и отчасти древесный и подземный образ жизни, усложняющий откладывание яиц. Так, живородящи морские змеи и некоторые хамелеоны.
В заключение подчеркнем, что живорождение (во всех его вариантах) свойственно только чешуйчатым с их сравнительно просто организованными яйцами. У крокодилов и черепах яйца более сложного строения: покрыты белковой, а у некоторых и известковой оболочкой. И у этих животных нет даже намеков на живорождение.
Все это позволяет заключить, что среди современных рептилий намечаются два различных пути совершенствования размножения. Один состоит в совершенствовании яйцекладности (усложнение строения яйца, устройство хотя бы примитивных гнезд, начальные элементы охраны будущего потомства); это свойственно крокодилам, черепахам и крупным змеям. Второй путь — живорождение в разных модификациях, вплоть до формирования аллантоидной плаценты, принципиально не отличимой от плаценты млекопитающих. Этот путь демонстрируют чешуйчатые.
Количество яиц, откладываемых рептилиями, сравнительно невелико. Некоторые крокодилы, крупные черепахи и змеи откладывают 50— 100 яиц, но обычно не более 30. Мелкие виды ящериц откладывают всего 1 — 2 яйца. Любопытно, что в этом случае бывает несколько кладок в год. Откладывание яиц в несколько приемов свойственно и некоторым некрупным черепахам. В общем плодовитость у рептилий заметно меньше, чем у амфибий и тем более рыб, что говорит о прямой связи с более совершенным стилем всего процесса размножения рептилий.
Половая зрелость наступает в различные сроки: у крокодилов и большинства черепах в 8 — 10-летнем возрасте, у змей — чаще в возрасте 3 — 5 лет, у ящериц обычно на 2 — 3-м году жизни, но некоторые мелкие виды становятся половозрелыми уже на следующую весну, т.е. в возрасте 9 — 10 месяцев. Крайне интересно, что у некоторых скальных ящериц, распространенных в Восточном Закавказье (например, Lacerta armeniaca), самцы неизвестны вовсе или встречаются крайне редко. Самки размножаются партеногенетически и рождают только самок.
Говоря о взаимоотношении полов у рептилий, отметим, что у большинства видов самки и самцы расходятся после совокупления. Есть случаи объединения в пары на длительное время. Таковы некоторые агамы, ящерицы и немногие черепахи. По некоторым наблюдениям, самки миссисипского аллигатора не только строят гнездо и охраняют его, но и помогают выведшимся молодым выбраться из гнезда. Молодые держатся вместе с матерью по крайней мере в течение одного сезона.
Экономическое значение и охрана пресмыкающихся
Среди пресмыкающихся фауны России нет видов, вредящих хозяйству человека: все они заслуживают самого бережного отношения. Ящерицы и змеи уничтожают насекомых и мелких грызунов — вредителей сельского и лесного хозяйства. Например, прыткая ящерица в средней полосе европейской части России поедает насекомых- вредителей чаще, чем обыкновенная овсянка и пеночка-весничка, и лишь немного реже, чем садовая славка. Опасность ядовитых змей неоправданно преувеличена. Сами змеи делают попытку укусить человека только в том случае, если их потревожить или преследовать. Многолетний опыт наблюдений за змеями, широко практикуемый в южных районах нашей страны, наглядно это подтверждает. Вместе с тем надо учесть исключительно большую ценность змеиного яда, используемого в медицинской промышленности. Это обстоятельство послужило предпосылкой дня организации специальных питомников, в которых содержат отловленных змей и периодически берут от них яд. Такие питомники есть в тропической Азии, на юге Африки, в Южной Америке. Ряд питомников организован в Средней Азии. В них содержат кобр, гюрз, степных гадюк и змей некоторых других видов. К сожалению, змеи в питомниках не размножаются.
Охрана рептилий. В естественных природных биоценозах рептилии нигде не достигают высокой численности. Человек часто неоправданно преследует и добывает рептилий, используя в пищу их мясо и яйца. Кожу крупных ящериц, змей, крокодилов используют для различных поделок. Красивый роговой панцирь морских черепах бисс идет на изготовление изящных женских украшений. Неумеренный промысел гигантских слоновых и крупных морских черепах привел к исчезновению одних видов и поставил на грань исчезновения другие. Гигантские сухопутные черепахи были почти истреблены мореплавателями, китобоями и пиратами в XVII—XIX вв. На Галапагосских островах раньше обитало 15 видов (с многими подвидами) слоновых черепах (Geochelone), сейчас сохранилось 11, шесть из них разводят в неволе для расселения по местам прежнего обитания. Только на двух из 13 Галапагосских островов (раньше черепахи обитали на 11 островах) численность слоновых черепах достаточно высока, на остальных нужны специальные меры для их сохранения.
Использование деликатесного мяса и яиц морских черепах местными жителями и гурманами во многих странах, продолжающийся браконьерский промысел прибывающих в период размножения на побережья поставили на грань гибели многие виды морских черепах. К сожалению, не везде запретительные меры приводят к эффективной охране и восстановлению их численности.
Все виды и подвиды слоновой черепахи внесены в Красную книгу МСОП. На Галапагосских островах создан Национальный парк, а в 1964 г. открыта Биологическая станция им. Ч. Дарвина.
Промысел крокодилов теперь везде ограничен, во многих местах запрещен, созданы специальные питомники и фермы по их разведению.
В Красную книгу МСОП занесена самая ценная в гастрономическом отношении зеленая, или суповая, черепаха (Chelonia mydas), которая могла быть важнейшим источником пищи на берегах тропических морей, если бы ее не уничтожали так быстро. Популяция ее и сейчас значительна, но мест, где она размножается, осталось очень мало. Не вымерла она только благодаря резервату Тортугеро в Коста-Рике, откуда в последние десятилетия ежегодно вывозится 20 тыс. вылупившихся черепашек в разные области Карибского бассейна.
Из настоящих крокодилов в Красную книгу МСОП занесено 15 видов, в том числе кубинский крокодил (Crocodylus rhombifer). Промысел его запрещен. В болотах полуострова Сапата создан крокодиловый питомник.
В Красной книге МСОП находятся несколько видов галапагосских наземных игуан (Conolopus), гигантский комодосский варан (Varanus komodoensis).
В Красную книгу Российской Федерации (2001 г.) занесен 21 вид рептилий. Среди них кавказская гадюка (Vipera kaznakovi), эндемик Западного Кавказа. Этот вид внесен также в Красную книгу МСОП. В Красную книгу Российской Федерации занесены дальневосточная кожистая черепаха (Trionyx sinensis), распространенная в Приморье, средиземноморская черепаха (Testudo graeca), встречающаяся в Краснодарском крае и в Дагестане.
Большинство «краснокнижных видов» в России находится у северных пределов ареалов, где их численность продолжает сокращаться, хотя в Российской Федерации существует закон, охраняющий виды, включенные в Красную книгу РФ.