Зоология позвоночных - В. М. Константинов 2011
Хордовые
Наземные, или четвероногие, позвоночные - Позвоночные с зародышевыми оболочками
Класс Млекопитающие, или звери
Общая характеристика
Млекопитающие — наиболее высокоорганизованный класс позвоночных животных. Размеры их тела различны: у карликовой белозубки — 3,5 см, синего кита — 33 м, масса тела соответственно 1,5 г и 120 т. Основные прогрессивные черты млекопитающих следующие:
1) высокий уровень развития центральной нервной системы, в первую очередь коры полушарий переднего мозга — центра высшей нервной деятельности. В связи с этим приспособительные реакции млекопитающих на условия среды весьма сложны и совершенны;
2) живорождение и выкармливание детенышей продуктом материнского организма — молоком, что позволяет млекопитающим приносить потомство при крайне разнообразных условиях жизни;
3) высокоразвитая способность к терморегуляции, обусловившая относительно постоянную температуру тела. Это вызвано, с одной стороны, регуляцией теплообразования при окислительных процессах (так называемая химическая терморегуляция), с другой — регулированием сохранения и отдачи тепла путем изменения кожного кровоснабжения и испарения воды при дыхании и потоотделении, развитием шерстного покрова и подкожного жира (так называемая физическая терморегуляция).
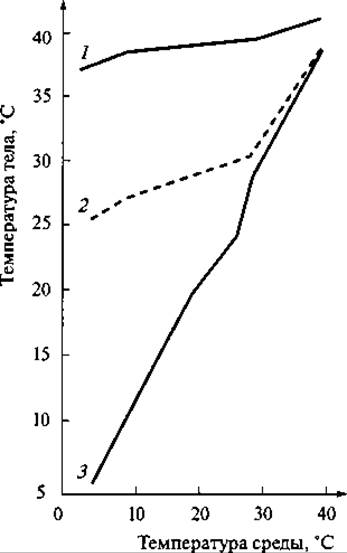
Рис. 164. Кривые зависимости температуры тела различных животных от температуры внешней среды:
1 — кролик; 2 — ехидна; 3 — ящерица
Благодаря описанным механизмам температура тела у многих млекопитающих относительно постоянна, и ее отличие от температуры внешней среды может быть равно примерно 100°С. Так, у песца, обитающего зимой при температуре до -60°С, температура тела равна примерно 39°С. Следует, однако, иметь в виду, что постоянство температуры тела (гомойотермна) не является абсолютным признаком всех млекопитающих.
В полной мере она характерна для плацентарных зверей, имеющих относительно крупные размеры. У низших млекопитающих, у которых менее развит терморегуляционный механизм, и у мелких плацентарных зверей, имеющих невыгодное для сохранения тепла соотношение между объемом тела и его поверхностью, температура тела меняется в значительных пределах в зависимости от температуры внешней среды (рис. 164). Так, у сумчатой крысы температура тела меняется в пределах 37,8 — 29,3°С, у наиболее примитивных насекомоядных (тенреков) — 34 — 13°С, у одного из видов броненосцев — 40 — 27°С, у обыкновенной полевки — 37 — 32°С.
Огромное значение в регулировании отдачи тепла имеет шерстный покров, а у некоторых и подкожный жировой слой.
Эти особенности, равно как и ряд других черт организации, обусловили возможность широкого распространения млекопитающих в самых разнообразных условиях. Географически они обитают почти повсеместно, за исключением Антарктиды. Еще более важно учесть, что млекопитающие населяют самые разнообразные жизненные среды. Помимо многочисленных наземных видов есть виды летающие, полуводные, водные и, наконец, заселяющие толщу почвы. Общее число видов современных млекопитающих составляет примерно 4,5 тыс.
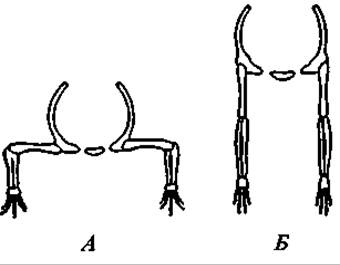
Рис. 165. Положение туловища и конечностей у примитивного наземного позвоночного (А) и у млекопитающего (Б)
Морфологически млекопитающие характеризуются следующими признаками. Тело покрыто шерстью (исключения редки и имеют вторичный характер). Кожа богата железами. Особо должны быть отмечены млечные железы. Череп сочленяется с позвоночником двумя затылочными мыщелками. Нижняя челюсть состоит только из зубной кости. Квадратная и сочленовная кости превращаются в слуховые косточки и располагаются в полости среднего уха. Зубы дифференцированы на резцы, клыки и коренные; они сидят в альвеолах. Локтевой сустав направлен назад, коленный — вперед в отличие от низших наземных позвоночных, у которых оба эти сустава направлены вбок наружу (рис. 165). При полном разделении потоков артериальной и венозной крови сердце четырехкамерное, сохраняется одна левая дуга аорты. Эритроциты безъядерны.
Морфофизиологический обзор
Кожный покров. Кожный покров млекопитающих полифункционален. Он ограничивает и защищает тело с поверхности, участвует в терморегуляции, предохраняя животное и от перегрева, и от охлаждения, участвует в выражении полового диморфизма, через кожу идет процесс дыхания и выделения. В этой связи кожный покров имеет сложное строение (рис. 166).
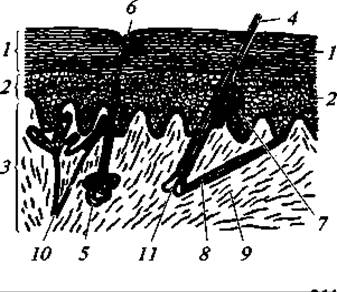
Рис. 166. Строение кожи млекопитающего (сильное увеличение):
1 — наружный слой рогового эпидермиса, клеточки которого спадают время от времени; 2 — глубокий слой эпидермиса с живыми клетками (мальпигиев слой); 3 — собственно кожа (кутис); 4 — волос; 5 — потовая железа; 6 — отверстие ее протока; 7 — сальная железа; 8 — мышцы волоса; 9 — соединительнотканные волокна кожи; 10 — кровеносный сосуд; 11 — сосочек в основании волоса
Как и у других позвоночных, кожа млекопитающих состоит из двух слоев: наружного — эпидермиса и внутреннего — кутиса, или собственно кожи. Эпидермис имеет два слоя: глубокий слой представлен живыми цилиндрическими или кубическими клетками, известен под названием мальпигиева, или росткового, слоя. Ближе к поверхности клетки более плоские. Полости этих клеток постепенно заполняет кератогиалин, что ведет к ее роговому перерождению и отмиранию. Поверхностно расположенные клетки постепенно снашиваются в виде мелкой «перхоти» или целыми лоскутами (так, например, бывает у тюленей). Снашивание рогового слоя эпидермиса восполняется постоянным нарастанием его за счет деления клеток мальпигиева слоя.
Эпидермис дает начало многим производным кожи, основные из которых — волосы, ногти, когти, копыта, рога (кроме оленьих), чешуи, различные железы. Образования эти описаны далее.
Собственно кожа, или кутис, у млекопитающих развита очень хорошо. Она состоит преимущественно из волокнистой соединительной ткани, сплетения волокон которой образуют сложный узор. В верхней части кутиса ветвятся кровеносные сосуды, лежат основания волосяных сумок, потовых желез. Нижняя часть кутиса состоит из очень рыхлой волокнистой ткани, в которой откладывается жир. Слой этот носит название подкожной жировой клетчатки. Наибольшего развития он достигает у водных зверей — китов, тюленей, у которых он в связи с полной (у китов) или частичной (у тюленей) редукцией волосяного покрова и физическими особенностями водной среды выполняет термоизоляционную роль. У некоторых наземных зверей также бывают большие подкожные жировые отложения. Особенно сильно накапливают жир виды, впадающие на зиму в спячку (суслики, сурки, барсуки и др.). Для них жир во время спячки служит основным энергетическим материалом.
Толщина кожи существенно различна у разных видов. Как правило, у видов холодных стран, имеющих пышный волосяной покров, она тоньше. Очень тонкая и непрочная кожа характерна для зайцев, к тому же она бедна кровеносными сосудами. Это имеет приспособительное значение. Хищник, схвативший зайца за кожу, легко вырывает из нее кусок, упуская самого зверька. Образовавшаяся же рана почти не кровоточит и быстро заживает. Своеобразная кожная хвостовая автотомия наблюдается у некоторых мышей, сонь, тушканчиков. Кожный хвостовой футляр у них легко обрывается и соскальзывает с хвостовых позвонков, что дает возможность схваченному за хвост зверьку уйти от врага.
Волосяной покров для млекопитающих столь же характерен, как покров из перьев для птиц или чешуйчатый покров для рептилий. Лишь немногие виды вторично полностью или частично его утратили. Так, вовсе не имеют волос дельфины, у китов есть зачатки волос на челюстях. У ластоногих волосяной покров редуцированный, особенно это заметно у моржей, в наименьшей мере — у ушастых тюленей (например, у котика), которые более чем другие виды ластоногих связаны с сушей.
Строение волоса видно на рис. 166. В нем можно различить выступающий над кожей ствол и сидящий в коже корень. Ствол состоит из сердцевины, коркового слоя и кожицы. Сердцевина представляет собой пористую ткань, между клетками которой находится воздух; именно эта часть волоса сообщает ему малую теплопроводность. Корковый слой, наоборот, очень плотный и придает волосу прочность. Тонкая наружная кожица защищает волос от механических и химических воздействий. Корень волоса в своей верхней части имеет цилиндрическую форму и является прямым продолжением ствола. В нижней части корень расширяется и заканчивается колбообразным вздутием — луковицей волоса, которая, как колпачок, охватывает вырост кутиса — волосяной сосочек. Кровеносные сосуды, входящие в этот сосочек, обеспечивают жизнедеятельность клеток луковицы волоса. Формирование и нарастание волос идет за счет размножения и видоизменения клеток луковицы. Ствол волоса является уже мертвым роговым образованием, неспособным к росту и изменению формы.
Погруженный в кожу корень волоса сидит в волосяной сумке. В воронку волосяной сумки открываются протоки сальных желез, секрет которых смазывает волос и сообщает ему большую прочность и водоупорность. К нижней части волосяной сумки прикрепляются мышечные волокна, сокращения которых вызывают движение сидящего в ней волоса. Это движение и обусловливает ощетинивание зверя.
Обычно волосы сидят в коже не перпендикулярно ее поверхности, а более или менее наклонно, т.е. прилегают к ней. Такой наклон волос выражен не у всех видов одинаково. Наименее заметен он у подземных зверей, например у крота.
Волосяной покров состоит из различных типов волос. Основные из них — это пуховые волосы, или пух, остевые волосы, или ость, чувствующие волосы, или вибриссы. У большинства видов основу шерстного покрова составляет густой низкий пух, или подшерсток. Между пуховыми волосами сидят более длинные, толстые и жесткие остевые волосы. У подземных зверей, например у крота, слепыша, меховой покров почти вовсе лишен остевых волос. Наоборот, у взрослых оленей, кабанов и тюленей редуцирован подшерсток, и покров состоит главным образом из ости. Отметим, что у молодых особей этих зверей подшерсток развит хорошо.
Волосяной покров периодически меняется. Смена волос, или линька, у некоторых видов бывает 2 раза в год: весной и осенью; таковы белка, лисица, песец, крот. Другие виды линяют только раз в год: весной выпадает старый мех, летом развивается новый, который окончательно созревает только к осени. Таковы, например, суслики.
Особую категорию волос представляют вибриссы. Это очень длинные жесткие волосы, выполняющие осязательную функцию.
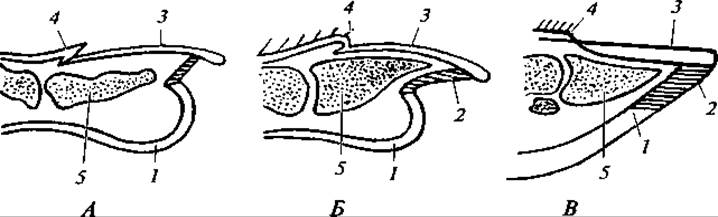
Рис. 167. Продольный разрез через конечные фаланги пальцев обезьяны (А), хищника (Б), копытного (В):
А — ноготь; Б — коготь; В — копыто: 1 — подушечка пальца; 2 — подошвенная пластинка; 3 — ногтевая пластинка; 4 — когтевой валик; 5 — конечная фаланга
Они сидят чаще на голове (так называемые усы), на нижней части шеи, на груди, а у некоторых лазающих древесных форм (например, у белки) и на брюхе. В основании волосяного мешка и в его стенках располагаются нервные рецепторы, воспринимающие соприкосновения стержня вибриссы с посторонними предметами.
Видоизменениями волос являются щетины и иглы.
Прочие роговые производные эпидермиса представлены роговыми чешуями, ногтями, когтями, копытами, полыми рогами, роговым клювом. Чешуя зверей по своему развитию и строению вполне сходна с одноименным образованием рептилий. Наиболее сильно развита чешуя у ящеров и панголинов, у которых она покрывает все тело. У очень многих мышевидных чешуйки имеются на лапах. Наконец, наличие чешуи на хвосте характерно для многих сумчатых, грызунов и насекомоядных.
Концевые фаланги пальцев подавляющего большинства зверей несут роговые придатки в виде ногтей, когтей или копыт. Наличие того или иного из этих образований и строение их находятся в прямой связи с условиями существования и образом жизни зверей (рис. 167). Так, у лазающих зверей пальцы имеют острые загнутые когти; у видов, роющих в земле норы, когти обычно несколько уплощены и расширены. Быстро бегающие крупные млекопитающие имеют копыта, при этом у лесных видов (например, у оленей), часто ходящих по болотам, копыта более широкие и плоские. У степных (антилопы) и особенно у горных видов (козлы, бараны) копыта маленькие, узкие; площадь опоры у них значительно меньше, чем у лесных копытных, ходящих зачастую по размягченному грунту или по снегу. Так, нагрузка на 1 см2 подошвы у центральноазиатского горного козла равна в среднем 850 г, у лося — 500 г, у северного оленя — 140 г.
Роговыми образованиями являются также рога быков, антилоп, козлов и баранов. Они развиваются из эпидермиса и сидят на костных стержнях, представляющих собой самостоятельные кости, сросшиеся с лобными костями. Рога оленей имеют иную природу. Они развиваются из кутиса и состоят из костного вещества.
Кожные железы у млекопитающих в отличие от птиц и рептилий многочисленны и разнообразны по строению и функции. Основные типы желез следующие: потовые, сальные, пахучие, млечные.
Потовые железы трубчатые, глубинные части их имеют вид клубка. Они открываются непосредственно на поверхности кожи. Продуктом выделения этих желез является пот, состоящий в основном из воды, в которой растворены мочевина и соли. Эти продукты не вырабатываются клетками желез, а поступают в них из кровеносных сосудов. Функция потовых желез заключается в охлаждении тела путем испарения выделяемой ими на поверхность кожи воды и в выделении продуктов обмена. Следовательно, эти железы выполняют одновременно терморегуляционную и выделительную функции. Потовые железы есть у большинства млекопитающих, но развиты они не у всех одинаково. Так, их очень мало у собак и кошек; у многих грызунов они имеются только на лапах, в паховой области и на губах. Совершенно отсутствуют потовые железы у китообразных, ящеров и некоторых других.
Сальные железы имеют гроздевидное строение и открываются почти всегда в воронку волосяной сумки. Жирный секрет этих желез смазывает волос и поверхностный слой эпидермиса кожи, предохраняя их от смачивания и снашивания.
Пахучие железы представляют видоизменение потовых или сальных желез, а иногда комбинацию тех и других (например, анальные железы куньих, секрет которых имеет очень резкий запах).
Особенно сильно развиты пахучие железы у американских скунсов, или вонючек (Mephitis), способных выпрыскивать большие порции выделений на значительное расстояние. Мускусные железы имеются у кабарги, выхухоли, бобра, ондатры; значение этих желез не вполне ясно, но, судя по тому, что наибольшее развитие они получают в период гона, деятельность их, видимо, связана с размножением; возможно, они стимулируют половое возбуждение.
Млечные железы — это своеобразное видоизменение простых трубчатых потовых желез. В простейшем случае — у австралийских однопроходных — они сохраняют трубчатое строение и открываются в сумки волос, расположенных группами на небольшом участке брюшной поверхности — так называемом железистом поле. У ехидны железистое поле находится в особой сумке, развивающейся в период размножения и служащей для вынашивания яйца, а потом и детеныша. У утконоса железистое поле располагается непосредственно на брюхе. Сосков у однопроходных нет, и детеныши слизывают молоко с волос, куда оно стекает из волосяных сумок.
У сумчатых и плацентарных млечные железы имеют гроздевидное строение и протоки их открываются на сосках. Расположение желез и сосков бывает различным. У лазающих по деревьям обезьян и висящих во время кормления летучих мышей имеется только одна пара сосков на груди; у бегающих копытных соски располагаются только в паховой области. У насекомоядных и хищных соски тянутся двумя рядами по всей нижней поверхности туловища. Число сосков стоит в прямой связи с плодовитостью вида и в некоторой степени соответствует числу одновременно рождающихся детенышей. Минимальное число сосков (2) характерно для обезьян, овец, коз, слонов и некоторых других; максимальное число сосков (10 — 24) свойственно мышевидным грызунам, насекомоядным, некоторым сумчатым.
Чрезвычайно важную роль у млекопитающих играют кожные покровы и их производные, обеспечивая механизмы физической терморегуляции, регулируя теплоотдачу. Большое значение имеет система кожных кровеносных сосудов: диаметр их просветов регулируется нервно-рефлекторным путем и может меняться в очень больших пределах. При расширении сосудов кожи теплоотдача резко увеличивается, при сужении, наоборот, сильно сокращается.
Охлаждение организма происходит также при испарении с поверхности кожи воды, выделяемой потовыми железами.
Густота и высота волосяного покрова у северных видов существенно меняются по сезонам. Так, у белки на огузке (задней части спины) на 1 см2 летом бывает в среднем 4 200 волос, зимой — 8 100, у зайца-беляка — 8 000 и 14700. Длина волос в миллиметрах на огузке такова: у белки летом пух — 9,4, ость — 17,4, а зимой — 16,8 и 25,9; у зайца-беляка летом пух — 12,3, ость — 26,4, зимой — 21,0 и 33,4. У тропических зверей таких резких изменений в длине шерсти не бывает в связи с малой разницей в температурных условиях зимой и летом.
В развитости потовых желез можно подметить географические и экологические закономерности. Так, среднее число этих желез на 1 см2 у разводимого во влажных тропиках зебу равно 1 700, а у крупного рогатого скота, разводимого в Англии (шорт-хорн), — всего 1 060. Эта же особенность прослеживается при сравнении видов, в разной мере приспособленных к засушливым условиям. В качестве показателя приводим величину испарения, выраженную в миллиграммах за минуту на 100 см2 поверхности кожи. При температуре 37 °С у осла эта величина достигает 17 мг/мин, у верблюда — только 3; при температуре 45°С у осла — 35, у верблюда — 15; наконец, при температуре 50 °С у осла — 45, у верблюда — 25 мг/мин (К. Шмидт-Ниельсон, 1972).
Не меньшее значение, чем участие в терморегуляции, имеет кожа в химической сигнализации млекопитающих.
Секрет кожных желез, как и другие пахнущие выделения (например, полового и пищеварительного трактов, моча, секрет специализированных желез), служит важным средством внутривидового общения — химической сигнализации млекопитающих. Особое значение этого типа сигнализации определяется дальностью ее действия и длительностью сохранения сигнала. У животных, имеющих определенные участки обитания, особи, пары, семьи метят участок запаховыми метками, которые они оставляют на приметных предметах: кочках, камнях, пнях, отдельных деревьях или просто на поверхности земли.
Запаховые метки оставляют родители на детенышах, в гнезде и на следах движения за пределами гнезда или места нахождения детеныша, если гнездо не сооружается. Благодаря химической сигнализации олени, тюлени и такие норники, как лисицы, песцы, соболи, куницы, полевки, мыши, находят своих, а не чужих детенышей. В целом запаховая сигнализация имеет решающее значение в поведении млекопитающих.
Мышечная система. Мышечная система млекопитающих очень дифференцирована и отличается большим числом разнообразно расположенных мускулов. Характерно наличие куполообразной мышцы — диафрагмы, отграничивающей брюшную полость от грудной. В основном ее роль заключается в изменении объема грудной полости, что связано с актом дыхания. Значительное развитие получает подкожная мускулатура, приводящая в движение те или иные участки кожи. У ежей и ящеров она обусловливает возможность свертывания тела в клубок. Поднятие игл у ежей и дикобразов, «ощетинивание» зверей, движение чувствующих волос — вибрисс — также вызываются действием этой мускулатуры. На лице она представлена мимической мускулатурой, особенно развитой у приматов.
Скелет. Скелет млекопитающих по своей структуре сохраняет типичные черты скелета других наземных позвоночных, но более дифференцирован (рис. 168). Осевой скелет включает позвоночник и осевой череп. Характерными чертами в строении позвоночного столба млекопитающих являются плоские сочленовные поверхности позвонков (платицельные позвонки), между которыми располагаются хрящевые диски (мениски), четко выраженная расчлененность позвоночника на отделы (шейный, грудной, поясничный, крестцовый, хвостовой) и постоянное число шейных позвонков. Отклонения от указанных признаков редки и носят вторичный характер.
В шейном отделе млекопитающих всегда семь позвонков, хорошо выражены атлант и эпистрофей. Исключение представляют ламантин, у которого 6 шейных позвонков, и виды ленивцев, имеющие от Шести до 10 шейных позвонков. Однако длина шейного отдела значительно варьирует. В отличие от птиц длина шеи у млекопитающих определяется не числом шейных позвонков, а длиной их тела. Наиболее сильно он развит у копытных, для которых подвижность головы очень важна при добыче пищи. Хорошо развита шея у хищников. Наоборот, у грызунов-норников и особенно у землероев шейный отдел короток и подвижность головы у них малая.
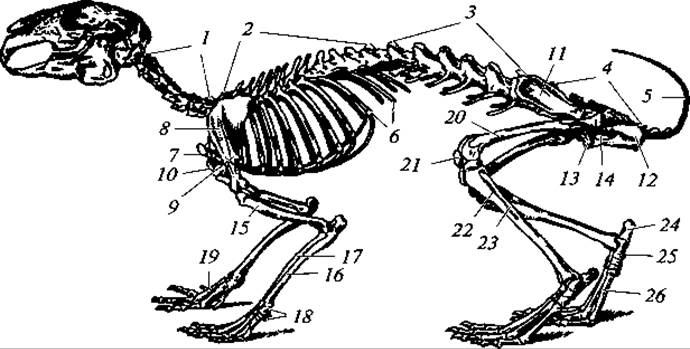
Рис. 168. Скелет кролика:
1 — шейные позвонки; 2 — грудные позвонки; 3 — поясничные позвонки; 4 — крестец; 5 — хвостовые позвонки; 6 — ребра; 7 — рукоятка грудины; 8 — лопатка; 9 — акромиальный отросток лопатки; 10 — коракоидный отросток лопатки; 11 — подвздошный отдел безымянной кости; 12 — седалищный отдел той же кости; 13 — лобковый отдел той же кости; 14 — запирательное отверстие; 15 — плечо; 16 — локтевая кость; 17 — лучевая кость; 18 — запястье; 19 — пясть; 20 — бедро; 21 — коленная чашечка; 22 — большая берцовая кость; 23 — малая берцовая кость; 24 — пяточная кость; 25 — таранная кость; 26 — плюсна
Грудной отдел состоит чаще из 12 — 15 позвонков; у одного из броненосцев и бутылконосого кита (Hyperoodon) их девять, а у ленивцев рода Choloepus — 24. К передним грудным позвонкам (обычно к семи) причленяются ребра, соединенные с грудиной (истинные ребра), образуя грудную клетку. Остальные грудные позвонки несут ребра, не доходящие до грудины (ложные ребра). Грудина представляет собой сегментированную костную пластинку, заканчивающуюся удлиненным хрящом — мечевидным отростком. Расширенный передний сегмент носит название рукоятки грудины. У летучих мышей и у зверей, конечности которых приспособлены для рытья, грудина теряет ясно выраженную сегментацию и несет киль, служащий, как и у птиц, для прикрепления грудных мышц.
В поясничном отделе число позвонков варьирует от двух до девяти. Позвонки эти несут рудиментарные ребра.
Крестцовый отдел чаще состоит из четырех сросшихся позвонков. При этом только первые два позвонка являются истинно крестцовыми, а остальные — приросшими к крестцу хвостовыми позвонками.
У хищных зверей крестцовых позвонков три, а у утконоса, как и у рептилий, — два. Число хвостовых позвонков наиболее изменчиво. Так, у гиббона их три, а у длиннохвостого ящера — 49.
Подвижность позвоночника у разных видов зверей различна. Наибольшую подвижность имеют мелкие зверьки, которые при движении часто выгибают спину дугой. Наоборот, у крупных копытных все отделы позвоночника (кроме шейного и хвостового) перемещаются незначительно, и при беге работают только конечности.
Череп млекопитающих (рис. 169) характеризуется относительно более крупной мозговой коробкой, что связано с большими размерами головного мозга. У молодых зверей мозговая коробка по сравнению с лицевой частью обычно развита относительно сильнее, чем у взрослых. Число отдельных костей в черепе млекопитающих меньше, чем у предыдущих групп позвоночных. Это обусловлено срастанием ряда костей между собой в комплексы, что особенно характерно для мозговой коробки. Так, срастаются основная, боковые и верхняя затылочная кости; сращение ушных костей приводит к образованию единой каменистой кости. Крылоклиновидная кость срастается с основной клиновидной костью, а глазоклиновидная кость срастается с передней клиновидной костью. Швы между костными комплексами зарастают сравнительно поздно, особенно в области мозгового черепа, что создает возможность увеличения объема головного мозга по мере роста животного.
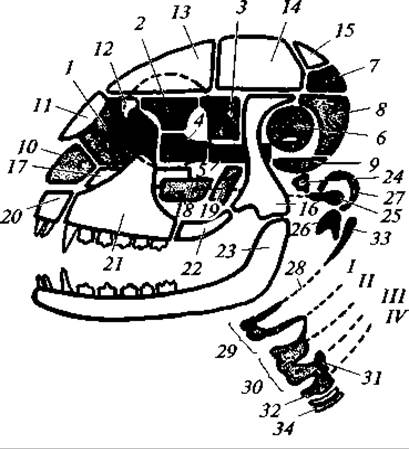
Рис. 169. Схема строения черепа млекопитающих:
1 — решетчатая кость; 2 — глазоклиновидная; 3 — крыловидная; 4 — передняя клиновидная; 5 — основная клиновидная; 6 — каменистая; 7 — верхняя затылочная; 8 — боковая затылочная; 9 — основная затылочная; 10 — хрящ носовой перегородки; 11 — носовая; 12 — слезная; 13 — лобная; 14 — теменная; 15 — межтеменная; 16 — чешуйчатая; 17— сошник; 18 — нёбная; 19 — крыловидная; 20 — межчелюстная; 21 — верхнечелюстная; 22 — скуловая; 23 — зубная; 24 — стремечко; 25 — наковальня; 26 — молоточек; 27 — барабанная кость; 28 — остатки гиоида; 29 — подъязычный аппарат; I — IV — остатки жаберных Дуг; 30 — щитовидный хрящ; 31 — черпаловидный хрящ; 32 — перстневидный хрящ; 33 — шиловидный отросток; 34 — трахея
Затылочная область сформирована единой, как указано, затылочной костью. Она имеет два мыщелка для сочленения с атлантом. Крыша мозгового черепа образована парными теменными, лобными и носовыми костями и непарной межтеменной костью. Бока черепной коробки образованы чешуйчатыми костями, от которых наружу и вперед отходят скуловые отростки. Последние соединяются со скуловой костью, которая, в свою очередь, спереди сочленена со скуловым отростком верхнечелюстной кости. В итоге образуется характерная для млекопитающих скуловая дуга.
Дно мозговой части черепа образовано основной и переднеклиновидной костями, а дно лицевой, висцеральной части — крыловидными, нёбными и верхнечелюстными костями. В дне черепа, в области слуховой капсулы, располагается свойственная только млекопитающим барабанная кость. Слуховые капсулы окостеневают, как уже указано, несколькими центрами, но в конечном счете формируется только одна парная каменистая кость.
Верхние челюсти состоят из парных межчелюстных и верхнечелюстных костей. Характерно развитие вторичного костного нёба, оно образовано нёбными отростками межчелюстных и верхнечелюстных костей и нёбными костями. В связи с образованием вторичного костного нёба хоаны открываются не между верхнечелюстными костями, как у других наземных позвоночных (кроме крокодилов и черепах), а позади нёбных костей. Такое строение нёба предотвращает перерыв дыхания в то время, как пищевой комок задерживается в ротовой полости для его пережевывания.
Нижняя челюсть представлена парными зубными костями, которые причленяются к скуловым отросткам чешуйчатых костей. Сочленовная кость превращается в слуховую косточку — молоточек; квадратная кость — в другую слуховую косточку — наковальню. Вместе со стремечком (гомолог гиомандибуляре) они лежат в полости среднего уха. Наружная стенка этой полости, а также часть наружного слухового прохода окружены упомянутой выше барабанной костью (tympanicum), гомологичной угловой кости — нижней челюсти прочих позвоночных. Таким образом, у млекопитающих наблюдается дальнейшее превращение части висцерального аппарата в слуховой аппарат среднего и наружного уха.
Плечевой пояс млекопитающих сравнительно прост. Основой его становится лопатка, к которой прирастает рудиментарный коракоид. Только у однопроходных коракоид существует в виде самостоятельной кости. Ключица имеется у тех млекопитающих, передние конечности которых совершают разнообразные сложные движения и у которых наличие ключицы обеспечивает более прочное причленение плечевой кости и упрочение всего плечевого пояса, например у обезьян. У видов, которые перемещают передние конечности только или преимущественно в плоскости, параллельной главной оси тела, ключицы рудиментарны или отсутствуют. Таковы копытные, грызуны и прочие.
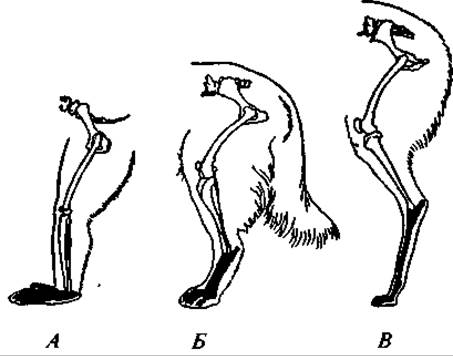
Рис. 170. Задние конечности стопо- и пальцеходящих млекопитающих (элементы стопы зачернены):
А — обезьяна павиан; Б — собака; В — лама
Тазовый пояс состоит из трех типичных для наземных позвоночных парных костей: подвздошных, седалищных к лобковых. У многих видов эти кости срастаются в одну безымянную кость.
Скелет парных конечностей сохраняет все основные черты строения типичной пятипалой конечности. Однако в связи с разнообразием условий существования и характером использования конечностей детали их строения весьма различны (рис. 170). У наземных форм значительно удлинены проксимальные отделы — бедро, голень. У водных зверей, наоборот, эти отделы укорочены, а удлинены дистальные отделы — пясть, плюсна и особенно фаланги пальцев. Превращенные в ласты конечности в этом случае двигаются отдельно от тела — чаще действуют как единое целое с туловищем. Перемещение же отделов конечностей относительно друг друга развито сравнительно слабо. У летучих мышей только первый палец передних конечностей развит нормально, остальные пальцы очень сильно удлинены; между ними расположена кожистая перепонка, образующая основную часть поверхности крыла. У быстро бегающих зверей предплюсна, плюсна, запястье и пясть располагаются более или менее отвесно, например у собаки. У наиболее совершенных бегунов — копытных — сокращается число пальцев. Первый палец атрофируется, и животные ступают либо на одинаково развитые третий и четвертый пальцы, между которыми проходит ось конечности (парнокопытные), либо преимущественное развитие получает один третий палец, через который и проходит ось конечности (непарнокопытные).
Укажем в этой связи максимальную скорость передвижения некоторых млекопитающих (в км/ч): короткохвостая землеройка — 4, красная полевка — 7, лесная мышь — 10, красная белка — 15, дикий кролик — 32 — 40, заяц — 55 — 72, лев — 50, красная лисица — 72, гепард — 105— 112, верблюд — 15 — Гб, африканский слон — 24 — 40, газель Гранта — 40 — 50.
Органы пищеварения. Органы пищеварения характеризуются большой сложностью, которая выражается в общем удлинении пищеварительного тракта, в большей, чем у других позвоночных, его дифференцировке и в лучшей развитости пищеварительных желез.
Пищеварительный тракт начинается предротовой полостью, или преддверием рта. Оно расположено между свойственными только млекопитающим мясистыми губами, щеками и челюстями. У ряда видов хомяков, бурундуков, обезьян преддверие рта, расширяясь, образует большие защечные мешки. Мясистые губы служат у детенышей для сосания молока, а у взрослых для схватывания пищи, а преддверие рта — для временного ее резервирования: хомяки и бурундуки переносят в защечных мешках запасы пищи в свои норы. Мясистых губ нет у однопроходных и китообразных. За челюстями лежит ротовая полость, в которой пища подвергается механическому измельчению и химическому воздействию. У зверей имеются четыре пары слюнных желез. Развитость слюнных желез находится в зависимости от характера питания. У китообразных они практически не развиты; у жвачных, наоборот, они получили исключительно сильное развитие. Так, корова в сутки выделяет около56 л слюны, которая имеет огромное значение для смачивания грубой пищи и для наполнения жидкой средой полостей желудка, где происходит бактериальное расщепление клетчатки пищевой массы.
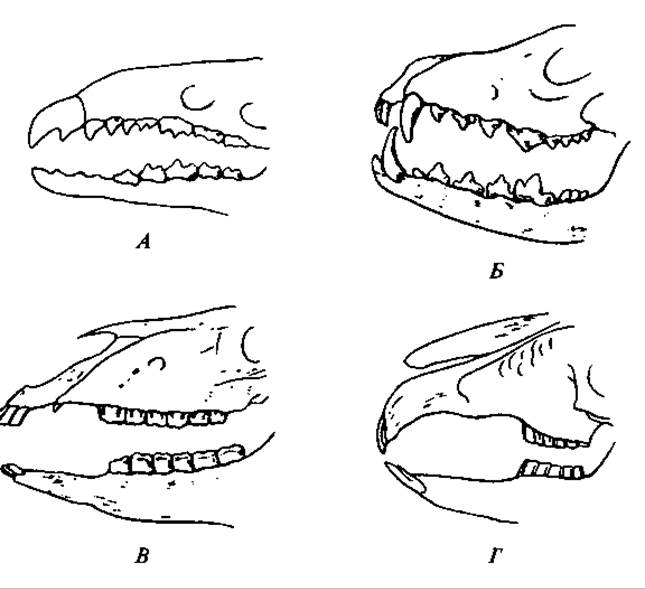
Рис. 171. Схема строения зубного ряда некоторых млекопитающих:
А — насекомоядные (землеройка); Б — хищные (лисица); В — однокопытные (лошадь); Г — зайцеобразные (заяц)
Секрет щечных желез летучих мышей, наносимый на летательные перепонки, сохраняет их эластичность и предохраняет от высыхания. Слюна вампиров, питающихся кровью, обладает антикоагулирующими свойствами, т.е. препятствует свертыванию крови. Слюна некоторых землероек ядовита, секрет их подчелюстной железы вызывает смерть мыши менее чем через 1 мин после инъекции. Токсичность слюнных желез примитивных млекопитающих рассматривается как отражение их филогенетической связи с рептилиями.
Млекопитающие гетеродонтны, т.е. их зубы дифференцированы на резцы, клыки, предкоренные, или ложные коренные, и коренные. Число зубов, их форма и функция существенно различаются у разных групп зверей (рис. 171). Так, например, малоспециализированные насекомоядные обладают большим числом сравнительно слабодифференцированных зубов. Для грызунов и зайцеобразных характерно сильное развитие резцов, отсутствие клыков и плоская жевательная поверхность коренных зубов. Такое строение зубной системы связано с характером питания: резцами они срезают или обгрызают растительность, а коренными зубами перетирают пищу, как жерновами. Хищные характеризуются сильно развитыми клыками; они используются для схватывания, а часто и для умерщвления добычи. Коренные зубы хищных имеют режущие вершины и плоские жевательные выступы. Задний ложнокоренной зуб верхней челюсти и первый истинно коренной нижней челюсти у хищных обычно выделяются своей величиной; они носят название «хищных» зубов.
Общее число зубов и их распределение по группам для видов зверей вполне определенно и постоянно и служит важным систематическим признаком. Для его обозначения пользуются зубными формулами, построенными в виде дробей. Числитель показывает число зубов в верхней челюсти, знаменатель — в нижней. Группы зубов обозначают начальными буквами их латинских названий: резцы — і (incisivi), клыки — с (canini), ложнокоренные — pm (praemolares), коренные — m (molares). Для сокращения пишут число зубов в одной половине челюсти.
Зубная формула волка такова:
![]()
Зубная формула кролика:
![]()
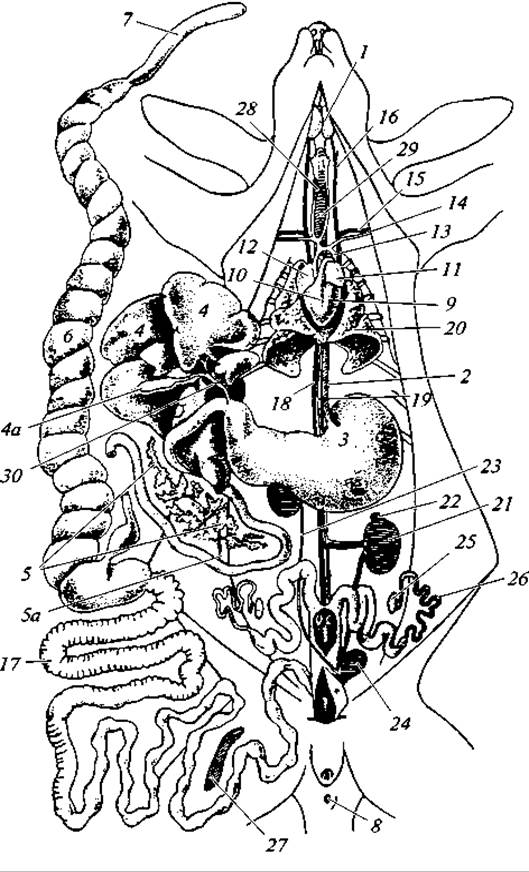
Рис. 172. Общее расположение внутренностей у самки кролика:
1 — подчелюстная слюнная железа; 2 — пищевод; 3 — желудок; 4 — печень (откинута кверху); 4а — желчный пузырь; 5 — поджелудочная железа; 5а — проток поджелудочной железы; 6 — слепая кишка; 7 — ее червеобразный отросток; 8 — заднепроходное отверстие; 9 — левый желудочек сердца; 10 — правый желудочек сердца; 11 — левое предсердие; 12 — правое предсердие; 13 — дуга аорты; 14 — левая подключичная артерия; 15 — левая подключичная вена; 16 — левая яремная вена; 17 — толстый кишечник; 18 — задняя полая вена; 19 — аорта; 20 — левое легкое; 21 — левая почка; 22 — левый надпочечник; 23 — правый мочеточник; 24 — мочевой пузырь; 25 — левый яичник; 26 — левый яйцевод; 27 — селезенка; 28 — щитовидная железа; 29 — трахея; 30 — желчный проток
Зубы сидят в ячейках челюстных костей, т.е. они текодонтные, и у большинства видов зверей меняются один раз в жизни (зубная система дифиодонтная).
Между ветвями нижней челюсти помещается мускулистый язык, служащий отчасти для схватывания пищи (бычьи, муравьеды, ящеры) и для лакания воды, отчасти для переворачивания пищи в полости рта во время ее пережевывания.
Позади ротовой области располагается глотка, в верхнюю часть которой открываются внутренние ноздри и евстахиевы трубы. На нижней поверхности глотки расположена щель, ведущая в гортань. Пищевод хорошо выражен (рис. 172). Мускулатура его чаще гладкая, но у некоторых, например у жвачных, сюда проникает из глоточной области поперечно-полостая мускулатура. Эта особенность обеспечивает произвольное сокращение пищевода при отрыгивании пищи.
Желудок явственно обособлен от других отделов пищеварительного тракта и снабжен многочисленными железами. Объем желудка и его внутреннее строение различны у разных видов, что связано с характером пищи (рис. 173). Наиболее просто устроен желудок у однопроходных, у которых он имеет вид простого мешка. У большинства желудок разделен на большее или меньшее число отделов.
Усложнение желудка связано со специализацией питания, например поглощением огромной массы грубых кормов (жвачные), или недоразвитостью ротового пережевывания пищи (некоторые виды, питающиеся насекомыми). У некоторых южноамериканских муравьедов в выходной части желудка дифференцируется отдел с настолько твердыми складками, что они выполняют функцию зубов, перетирающих пищу.
Сложно устроен желудок у жвачных копытных, например у коровы. Он состоит из четырех отделов: 1) рубца, внутренняя поверхность которого несет твердые вздутия; 2) сетки, стенки которой разделены на ячейки; 3) книжки со стенками, несущими продольные складки; 4) сычуга, или железистого желудка. Попавшие в рубец кормовые массы под влиянием слюны и деятельности бактерий и простейших подвергаются брожению. Из рубца пища благодаря перистальтическим движениям поступает в сетку, откуда она отрыгивается снова в рот. Здесь пища размельчается зубами и обильно смачивается слюной. Получившаяся таким образом полужидкая масса заглатывается и по узкому желобку, соединяющему пищевод с книжкой, поступает в нее и далее в сычуг (см. рис. 173).
Описанное приспособление имеет большое значение, так как пищей жвачных является трудноперевариваемая растительная масса, а в желудке у них живет огромное количество бродильных бактерий и простейших, деятельность которых существенно способствует перевариванию пищи, в том числе грубой клетчатки.
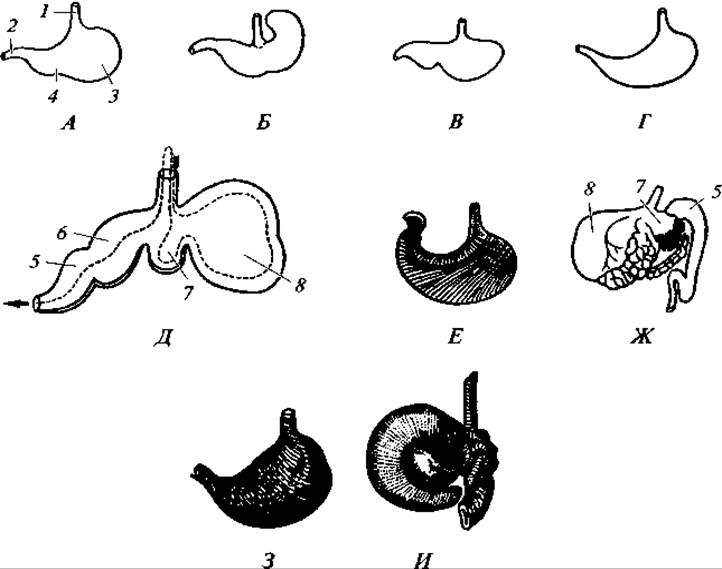
Рис. 173. Различные формы желудков млекопитающих:
А — собаки; Б — крысы; В — мыши; Г — куницы; Д — жвачного (пунктирная стрелка указывает направление движения пищи); Е — человека; Ж — верблюда; 3 — ехидны; И — трехпалого ленивца; 1 — пищевод; 2 — кишка; 3 — кардинальный отдел желудка; 4 — его пилорический отдел; 5 — сычуг; 6 — книжка; 7 — сетка; 8 — рубец
Собственно кишечник подразделяется на тонкий, толстый и прямой отделы. У видов, питающихся грубым растительным кормом (например, у грызунов), на границе тонкого и толстого отделов отходит длинная и широкая слепая кишка, заканчивающаяся у некоторых зверей (например, зайцы, полуобезьяны) червеобразным отростком. Слепая кишка выполняет роль «бродильного чана» и развита тем сильнее, чем больше растительной клетчатки поглощает животное. У мышей, питающихся семенами и отчасти вегетативными частями растений, слепая кишка составляет 7 — 10 % от общей длины всех отделов кишечника, а у полевок, кормящихся в большей мере вегетативными частями растений, — 18 — 27 %. У плотоядных видов слепая кишка развита слабо или отсутствует. В этой же связи варьирует и длина толстого отдела кишечника. У грызунов он составляет 29 — 53 % общей длины кишечного тракта, у насекомоядных и рукокрылых — 26 — 30 %, у хищных — 13 — 22 %. Общая длина кишечника варьирует очень сильно. Как правило, у растительноядных видов кишечник относительно более длинный, чем у всеядных и плотоядных. Так, у некоторых летучих мышей кишечник длиннее тела в 2,5 раза, у насекомоядных — в 2,5 — 4,2, у хищных — в 2,5 (ласка), в 6,3 (собака), у грызунов — в 5,0 (полуденная песчанка), в 11,5 (морская свинка), у лошади — в 12,0, у овцы — в 29 раз.
Таблица 14
Водный баланс североамериканского пустынного грызуна — кенгуровой крысы (по К. Шмидту-Ниельсону, 1982)
Поступление воды, % |
Потери воды, % |
||
Питьевая вода |
0 |
Испарение |
70 |
Пища |
10 |
С мочой |
25 |
Метаболическая вода |
90 |
С калом |
5 |
Характеризуя строение и функционирование пищеварительного тракта, коснемся вкратце проблемы обеспечения организма млекопитающих водой.
Многие виды хищников и копытных регулярно посещают водопой. Другие довольствуются водой, получаемой с сочной пищей. Однако есть и такие, которые никогда не пьют и кормятся очень сухими кормами, например многие пустынные грызуны. В этом случае основным источником водообеспечения служит вода, возникающая при обмене веществ, — так называемая метаболическая вода (табл. 14).
Метаболическая вода — один из обязательных продуктов превращения органических веществ в организме. Однако при метаболизме разных веществ образуется разное количество воды. Первое место занимают жиры. При использовании за сутки 1 кг жира образуется около 1 л воды, 1 кг крахмала — 0,5 л, 1 кг протеинов — 0,4 л (К. Шмидт-Ниельсон, 1982).
Под диафрагмой расположена печень. Желчный проток впадает в первую петлю тонких кишок. В этот же отдел кишечника впадает проток и поджелудочной железы, которая лежит в складке брюшины.
Органы дыхания. Как и у птиц, по существу единственными органами дыхания млекопитающих являются легкие. Роль кожи в газообмене незначительна: только около 1 % кислорода поступает через кожные кровеносные сосуды. Сказанное понятно, если учесть, во-первых, ороговение эпидермиса и, во-вторых, ничтожную общую поверхность кожи сравнительно с суммарной дыхательной поверхностью легких, которая у млекопитающих в 50—100 раз больше, чем поверхность кожи.
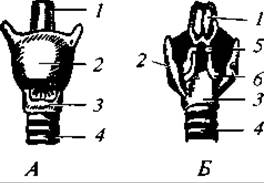
Рис. 174. Гортань кролика: вид спереди (А) и сзади (Б); 1 — надгортанник; 2 — щитовидный хрящ; З — перстневидный хрящ; 4 — трахея; 5 — сантонинов хрящ; 6 — черпаловидный хрящ
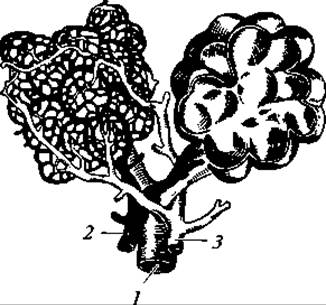
Рис. 175. Схема строения легочных пузырьков млекопитающего (на левом осталась только сеть капиллярных сосудов, на правом — вскрытый пузырек без сосудов):
1 — бронх; 2 — артерия; 3 — вена
Характерно усложнение верхней гортани (рис. 174). В основании ее лежит кольцеобразный перстневидный хрящ, передняя и боковые стенки гортани образованы свойственным только млекопитающим щитовидным хрящом. Над перстневидным хрящом по бокам спинной стороны гортани находятся парные черпаловидные хрящи. К переднему краю щитовидного хряща примыкает тонкий лепесткообразный надгортанник. Между перстневидным и щитовидным хрящами расположены небольшие мешковидные полости — желудочки гортани. Голосовые связки в виде парных складок слизистой оболочки гортани лежат между щитовидными и черпаловидными хрящами. Хорошо развиты трахея и бронхи. В области легких бронхи делятся на большое число мелких веточек. Самые мелкие веточки — бронхиолы — заканчиваются пузырьками — альвеолами, имеющими ячеистое строение (рис. 175). Здесь ветвятся кровеносные сосуды. Число альвеол огромно: у хищных их 300 — 500 млн, у малоподвижных ленивцев — около 6 млн. В связи с возникновением альвеол образуется огромная поверхность для газообмена. Так, например, общая поверхность альвеол у человека равна 90 м2. При расчете на единицу дыхательной поверхности (в см2) приходится у ленивца 6 альвеол, у домашней кошки — 28, у домовой мыши — 54, у летучей мыши — 100.
Обмен воздуха в легких обусловлен изменением объема грудной клетки, возникающим в результате движения ребер и особой, куполообразно вдающейся в грудную полость мышцы — диафрагмы. Число дыхательных движений зависит от величины животного, что связано с различием в интенсивности обмена веществ. Так, у лошади оно достигает 8 — 16 в 1 мин, у человека — 15 — 20, у крысы — 100 — 150, у мыши — около 200. Обратную зависимость между интенсивностью метаболизма и размерами тела наглядно демонстрирует табл. 15 по потреблению кислорода разными по размерам зверями.
Таблица 15
Потребление кислорода млекопитающими разных размеров
(по Л. Проссеру и др., 1977)
Вид |
Масса тела, г |
Потребление кислорода, мл/(г - ч) |
Землеройка |
3,5 |
7-10,6 |
Рыжая полевка |
22 |
2,3 |
Бурундук |
72-145 |
0,8 |
Заяц |
1600 |
0,96 |
Тюлень |
26000 |
0,6 |
Дельфин |
170000 |
0,22-0,34 |
Верблюд |
17000 - 330000 |
0,03-0,04 |
Слон |
3700000 |
0,07-0,11 |
Сумчатая мышь |
8,5 |
1,26 |
Кенгуру |
33000 |
0,2 |
Ехидна |
4000 |
0,22 |
Приведенные в таблице данные свидетельствуют также о более низком уровне обмена у однопроходных и сумчатых по сравнению с плацентарными, что соответствует примитивности их общей организации. Для сравнения укажем, что у чешуйчатых рептилий этот показатель (потребление кислорода, мл/(г ч)) равен всего 0,12 — 0,3, а у амфибий — 0,012 — 0,075 (Л. Проссер и др., 1977).
Соответственно всему сказанному теплопродукция у зверей высокая, хотя и в этом случае видна зависимость от размеров тела. Так, у землеройки она составляет 151,2 Дж/(г∙ ч), у кошки — 12,6, у собаки — 8,4, у слона — 2,1 Дж/(г ∙ ч) (К. Шмидт-Ниельсон, 1982).
Вентиляция легких не только обусловливает газообмен, но и имеет существенное значение для теплорегуляции. Особенно это характерно для видов со слаборазвитыми потовыми железами. У них охлаждение тела при его перегревании в значительной мере достигается повышением испарения воды, пары которой выводятся вместе с выдыхаемым из легких воздухом (так называемое полипное). Например, собака в покое выдыхает 2 л воздуха в минуту; при резком повышении температуры среды количество выдыхаемого воздуха доходит до 50 — 75 л/мин, а количество испаряемой воды может дойти до 200 см3/ч.
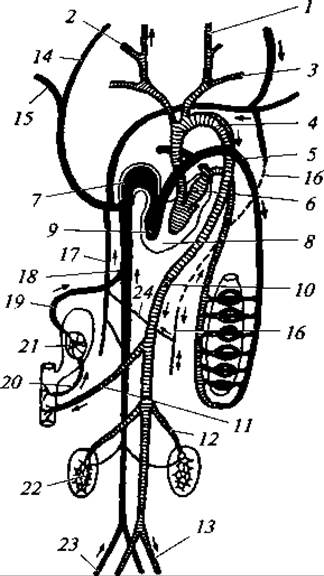
Рис. 176. Схема строения кровеносной системы млекопитающих: 1 — внешняя сонная артерия; 2 — внутренняя сонная артерия; 3 — подключичная артерия; 4 — дуга аорты; 5 — легочная артерия; 6 — левое предсердие; 7 — правое предсердие; 8 — левый желудочек; 9 — правый желудочек; 10 — спинная аорта; 11 — внутренностная артерия; 12 — почечная артерия; 13 — подвздошная артерия; 14 — яремная вена; 15 — подключичная вена; 16 — левая непарная вена; 17 — правая непарная вена; 18 — задняя полая вена; 19 — печеночная вена; 20— воротная вена печени; 21 — печень; 22 — почка; 23 — подвздошная вена; 24 — поперечная вена
Оценивая значение полипное для увеличения теплоотдачи, следует учитывать, что в этом случае увеличивается в первую очередь не легочная вентиляция, а вентиляция дыхательных путей. Так, у быка при общем росте вентиляции с 50 до 300 л/мин альвеолярная вентиляция возрастает с 25 до 75 л/мин, т.е. в 3 раза, а вентиляция мертвого пространства — с 25 до 225 л/мин, т.е. в 9 раз (К. Шмидт-Ниельсон, 1976).
Прослеживаются экологические особенности. Так, пустынные североамериканские грызуны — кенгуровые крысы — теряют с дыханием 50 — 57 мг воды на 1 мл потребленного кислорода, а белые (лабораторные) мыши и крысы — 85 — 94 мг (К. Шмидт-Ниельсон, 1982).
Кровеносная система. Как и у птиц, имеется только одна дуга аорты (но левая), отходящая от толстостенного левого желудочка. Главные артериальные сосуды отходят от аорты различно. Обычно от аорты отходит короткая безымянная артерия, которая делится на правую подключичную артерию, правую и левую сонные артерии, левая же подключичная артерия отходит самостоятельно от дуги аорты. В других случаях левая сонная артерия отходит не от безымянной артерии, а самостоятельно от дуги аорты. Спинная аорта, как и у всех позвоночных, лежит под позвоночным столбом. От нее отходит ряд ветвей к мускулатуре и внутренним органам.
Для венозной системы характерно отсутствие воротного кровообращения в почках. Левая передняя полая вена лишь у немногих видов впадает в сердце самостоятельно: чаще она сливается с правой передней полой веной, которая и изливает всю кровь от переднего отдела тела в правое предсердие. Характерно наличие остатков задних кардинальных вен — так называемых непарных вен. У большинства видов правая непарная вена самостоятельно впадает в переднюю полую вену, а левая непарная вена утрачивает связь с полой веной и впадает в правую непарную вену через поперечную вену (рис. 176).
Относительные размеры сердца различны у видов с разным образом жизни и в конечном счете с различной интенсивностью обмена веществ. Так, сердечный индекс (относительная масса сердца, выраженная в процентах к общей массе тела) у кашалота 0,3, у африканского слона — 0,4, у ленивца — 0,3, у серой полевки — 0,6, у ушана — 1,2 — 1,4, у обыкновенной землеройки — 1,4.
Связь между размерами тела и величиной сердца хорошо подтверждается и сопоставлением сердечного индекса систематически и экологически близких видов животных. Так, у крупного сурка- байбака этот показатель равен 0,47, у большого суслика — 0,61, а у малого суслика — 0,82.
Наряду с этим сердечный индекс зависит и от двигательной активности. Сравним этот показатель у сходных по вличине мышей, перемещающихся много и быстро, и серых полевок, относительно малоподвижных: полевая мышь — 0,89, лесная мышь — 0,85, полевка обыкновенная — 0,54, полевка-экономка — 0,47 (С. С. Шварц, 1968).
К сказанному добавим, что у домашнего кролика относительные размеры сердца в 3 раза меньше, чем у дикого зайца-русака. Примерно та же закономерность выявляется и при сличении этого показателя у комнатной и охотничьей гончей собак. В этой же связи различна и частота сокращений сердца (табл. 16).
Таблица 16
Частота пульса и интенсивность метаболизма у некоторых млекопитающих
Виды животных |
Масса тела, г |
Частота пульса в мин |
Интенсивность метаболизма на кг массы тела за 24 ч, кДж |
Мышь |
25 |
600 |
1680 |
Собака |
6500 |
120 |
277 |
Овца |
50000 |
70-80 |
113 |
Бык |
500000 |
43 |
101 |
Таблица 17
Особенности крови у разных позвоночных (по П. В. Терентьеву, 1961; Л. Проссеру и Ф. Брауну, 1967; Л. Проссеру и др., 1977)
Группы позвоночных |
Количество крови, % к массе тела |
Количество эритроцитов в 1 мм3 крови, млн |
Кислородная емкость крови, % на единицу объема |
Рыбы хрящевые |
— |
0,15 |
6,0 |
Рыбы костные |
1,5-3,0 |
— |
8,8 |
Амфибии хвостатые |
3,0 |
0,12 |
3,0-10,0 |
Амфибии бесхвостые |
4,2-4,9 |
0,48 |
— |
Рептилии |
4,2-7,4 |
0,98 |
7,0-12,0 |
Птицы |
5,7-9,0 |
2,7-3,5 |
10,0-22,0 |
Млекопитающие |
5,5-9,5 |
8,4 |
15,0-24,0 |
В связи с интенсивностью сердечной деятельности кровяное давление у млекопитающих столь же высокое, как и у птиц. У морского слона оно равно 120/90 мм рт. ст., у крысы — 130/90, у собаки — 112/56 мм рт. ст. Для сравнения укажем, что у чешуйчатых рептилий этот показатель лежит в пределах 14/10 — 80/60, а у амфибий 22/12 — 30/25 (Л. Проссер и др., 1978).
Общее количество крови у млекопитающих больше, чем у позвоночных нижестоящих групп. Выгодно отличается кровь млекопитающих и по ряду ее биохимических свойств, отчасти связанных с безъядерностью эритроцитов. Млекопитающие обладают не только относительно большим количеством крови, но, что еще важнее, большой ее кислородной емкостью. В свою очередь, это связано с большим числом эритроцитов и большим количеством гемоглобина. У низших позвоночных (рыбы, амфибии) его содержится в среднем 5 — 10 г, а у млекопитающих — 10 — 15 г на 100 см3 крови. Некоторые обобщенные данные по особенностям крови у разных позвоночных приведены в табл. 17.
Своеобразные приспособления возникают при водном образе жизни, когда возможность атмосферного дыхания периодически прерывается. Это выражается, с одной стороны, в резком увеличении количества связывающего кислород глобина в мышцах (миоглобина) — около 50 % всего глобина организма. Кроме того, у надолго погружающихся в воду зверей отключается периферическое кровообращение, на постоянном уровне остается кровоснабжение мозга и сердца.
С другой стороны, у водных и полуводных млекопитающих при погружении в воду уменьшается число сердцебиений (так называемая брадикардия), что приводит к замедленному току крови и более полному использованию кислорода крови. Например, у тюленя (Phoca vitulinа), находящегося в непогруженном состоянии, число сокращений сердца в 1 мин равно 180. Через 11 с после погружения оно уменьшается до 60, через 27 с — до 35 и далее устанавливается на уровне 30 в течение всего времени нахождения зверя под водой.
Кислородная емкость крови у ныряющих животных наибольшая. Так, у утконоса она равна 24 об. %, у ондатры — 25, у сивуча — 19, у обыкновенного дельфина — 26, у кашалота — 29 об. %.
Кислородная емкость крови меняется и при изменении разреженности атмосферы по отношению к высоте над уровнем моря.
Конечно, повышение кислородной емкости крови зависит и от степени двигательной активности вида. Винторогий козел более подвижен, чем тур, и еще более, чем домашние овцы и козы, поэтому у него большая кислородная емкость крови.
Учитывая все изложенное о явлениях газообмена и кровообращения, есть все основания заключить, что общий уровень метаболизма у млекопитающих заметно более высокий, чем у их филогенетических предшественников, и близок к таковому у птиц.
Кроветворные органы специализированы. Костный мозг продуцирует эритроциты, гранулоциты, тромбоциты. Селезенка и лимфатические железы — лимфоциты.
Нервная система. Нервная система млекопитающих, как и у других позвоночных, включает центральную: головной и спинной мозг и периферическую — отходящие от них нервы. Головной мозг (рис. 177) относительно крупных размеров, что обусловливается увеличением объема полушарий переднего мозга и мозжечка.
Развитие переднего мозга выражается в разрастании его крыши — вторичного мозгового свода (неопаллиума), а не полосатых тел, как у птиц; он состоит из нервных клеток и безмякотных нервных волокон. В связи с развитием коры мозга серое мозговое вещество у млекопитающих расположено поверх белого вещества. В коре мозга расположены центры высшей нервной деятельности. Сложное поведение млекопитающих, сложные реакции их на различные внешние раздражения прямым образом связаны с прогрессивным развитием коры полушарий переднего мозга. Кора обоих полушарий связана комиссурой из белых нервных волокон, так называемым мозолистым телом.
Отношение массы полушарий переднего мозга к массе всего головного мозга различно у млекопитающих разных систематических групп. У ежей оно равно 48 %, у белок — 53, у волков — 70, у дельфинов — 75 %.
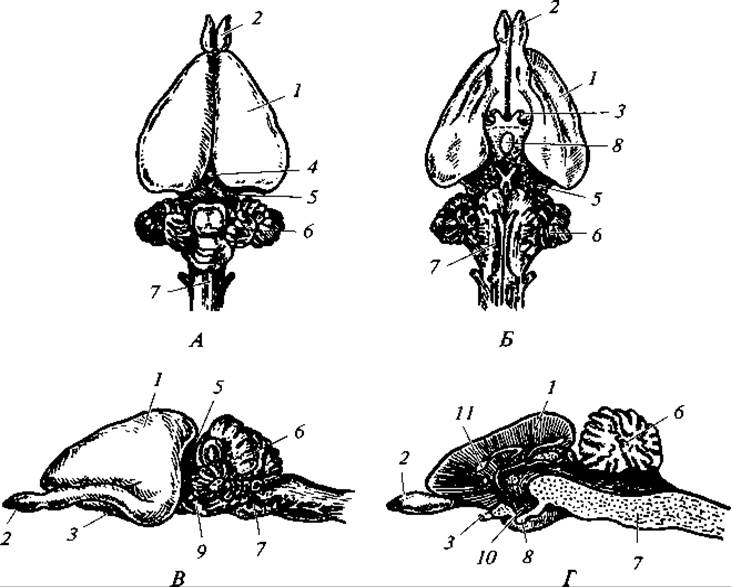
Рис. 177. Мозг кролика:
вид сверху (А); снизу (Б); сбоку (В); Г — продольный разрез; 1 — большие полушария; 2 — обонятельные доли; 3 — зрительный нерв; 4 — эпифиз; 5 — средний мозг; 6 — мозжечок; 7 — продолговатый мозг; 8 — гипофиз; 9 — варолиев мост; 10 — мозговая воронка; 11 — мозолистое тело
Кора переднего мозга у большинства видов достаточно мощная и включает 7 слоев нервных клеток. Она покрыта многочисленными бороздами, увеличивающими ее площадь: сильвиева борозда, отделяющая лобную долю коры от височной доли, роландова борозда, отделяющая сверху лобную долю от теменной. У высших представителей класса число борозд велико. Промежуточный мозг сверху невиден. Эпифиз, гипофиз и гипоталамус невелики, но чрезвычайно важны функционально.
Для среднего мозга характерно подразделение его двумя взаимно перпендикулярными бороздами на четыре бугра. Мозжечок велик и дифференцирован на несколько отделов (см. рис. 177), что связано с очень сложным характером движений у зверей.
Продолговатый мозг важен ядрами центров дыхания, кровообращения, пищеварения и пр.
Органы чувств. Обонятельные органы развиты у млекопитающих очень хорошо, лучше, чем у других наземных позвоночных, и играют в их жизни огромную роль. При помощи этих органов млекопитающие опознают врагов, отыскивают пищу, а также друг друга. Многие виды чувствуют запахи за несколько сот метров и способны обнаруживать пищевые объекты, находящиеся под землей. Только у полностью водных зверей (китов) обоняние понижено. Тюлени же обладают очень острым обонянием.
Прогрессивное развитие описываемых органов выражается в увеличении объема обонятельной капсулы и в ее усложнении вследствие образования системы обонятельных раковин. У некоторых групп зверей (сумчатые, грызуны, копытные) имеется обособленный отдел обонятельной капсулы, открывающийся самостоятельно в нёбноносовой канал (якобсонов орган), служащий для распознавания запаха пищи во рту, который уже был описан в главе о рептилиях.
Орган слуха (рис. 178) в подавляющем большинстве случаев развит очень хорошо. В состав его кроме внутреннего и среднего уха, имеющихся и у нижестоящих классов, входят еще два новых отдела: наружный слуховой проход и ушная раковина. Последняя отсутствует только у водных и подземных зверей (киты, большинство ластоногих, слепыши и некоторые другие). Ушная раковина направленно улавливает звуки и существенно усиливает тонкость слуха. Особенно сильно она развита у ночных зверей (летучие мыши) и у лесных копытных, пустынных собачьих и некоторых других. Внутренний конец слухового прохода затянут барабанной перепонкой, за которой лежит полость среднего уха. В ней у млекопитающих находится не одна, как у амфибий, рептилий и птиц, а три слуховые косточки (рис. 179). Молоточек (гомолог сочленовной кости) упирается в барабанную перепонку, к нему подвижно прикреплена наковальня (гомолог квадратной кости), которая, в свою очередь, соединена со стремечком (гомолог гиомандибуляре), а это последнее упирается в овальное окно перепончатого лабиринта внутреннего уха. Описанная система обеспечивает значительно более совершенную передачу звуковой волны, уловленной ушной раковиной и прошедшей по слуховому проходу к среднему, а затем к внутреннему уху. В строении внутреннего уха зверей обращает внимание сильное развитие улитки и наличие кортиева органа — тончайших волокон, которые в числе нескольких тысяч натянуты в канале улитки. При восприятии звука эти волокна резонируют, чем обеспечивается более тонкий слух млекопитающих.
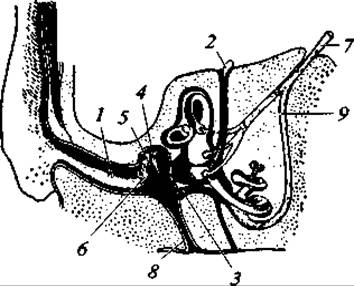
Рис. 178. Схема органа слуха млекопитающего:
1 — наружный слуховой проход; 2 — эндолимфатический канал; 3 — круглое окно; 4 — наковальня; 5 — молоточек; 6 — барабанная перепонка; 7 — слуховой нерв; 8 — евстахиева труба; 9 — нерв улитки
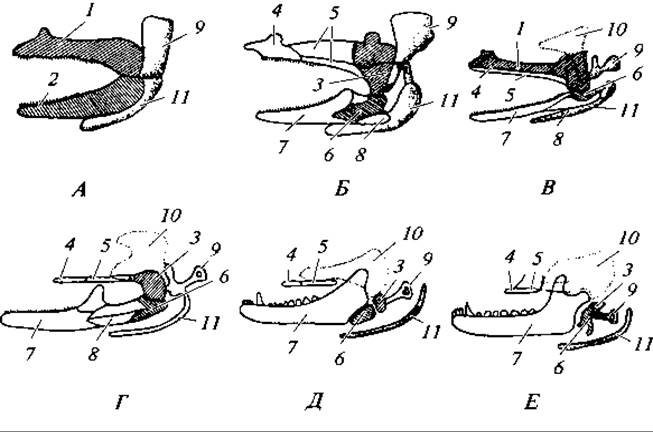
Рис. 179. Преобразование первых двух висцеральных дуг в ряду позвоночных:
А — акула; Б — костная рыба; В — амфибия; Г — рептилия; Д — териодонт; Е — млекопитающее; 1 — нёбно-квадратный хрящ; 2 — меккелев хрящ; 3 — квадратная кость — наковальня млекопитающих; 4 — нёбная кость; 5 — крыловидные кости; 6 — сочленовная кость — молоточек млекопитающих; 7 — зубная кость; 8 — угловая кость; 9 — гиомандибулярный хрящ — слуховая косточка (стремечко) наземных позвоночных; 10 — чешуйчатая кость; 11 — гиоид
У ряда зверей обнаружена способность к эхолокации. Кроме хорошо известных в этом отношении летучих мышей такой способностью обладают китообразные (дельфины), ластоногие (тюлени), землеройки. Землеройки при эхолокации издают импульсы частотой 30 — 60 кГц, продолжительностью 5 — 33 мс. Морской лев при локации издает звуки частотой от 20 до 72 кГц, дельфины — 120 — 200 кГц. Последние способны лоцировать косяки рыб с расстояния до 3 км.
Органы зрения в жизни млекопитающих имеют меньшее значение, чем у птиц. На неподвижные предметы они обычно обращают мало внимания, и к стоящему человеку могут вплотную подойти даже такие осторожные звери, как лисицы, зайцы, лоси. Острота зрения и развитие глаз, естественно, различны и связаны с условиями существования. Особенно большие глаза имеют ночные звери и животные открытых ландшафтов (например, антилопы). У лесных зверей зрение менее острое, а у подземных глаза редуцированы и иногда затянуты кожистой перепонкой (слепыш).
Аккомодация у млекопитающих происходит только путем изменения формы хрусталика под действием ресничной мышцы. У мелких грызунов (полевки, мыши) способности к аккомодации практически нет, что связано преимущественно с ночной активностью и незначительностью обзора.
Цветное зрение у млекопитающих сравнительно с птицами развито слабо. Почти весь спектр способны различать лишь высшие обезьяны Восточного полушария. Европейская рыжая полевка различает только красный и желтый цвета. У опоссума, лесного хоря и ряда других видов цветное зрение вообще не обнаружено.
Характерной особенностью органов осязания млекопитающих является наличие осязательных волос, или вибрисс (рис. 180), описание которых приведено ранее при характеристике покровов.
Выделительная система. Почки у млекопитающих тазовые — метанефрические. Туловищные почки закладываются эмбрионально, но в последующем редуцируются. Почки — компактные органы, бобовидной или дольчатой формы. Поверхность их чаще гладкая, иногда бугорчатая (жвачные, кошки), у некоторых (например, у китообразных) почки разделены перехватами на доли.
На разрезе видно, что почка имеет слои — наружный корковый и внутренний (полосатый) мозговой (рис. 181). В корковом слое расположены извитые канальцы, начинающиеся боуменовыми капсулами, внутри которых находятся клубки кровеносных сосудов (мальпигиевы тельца). В сосудистых клубочках осуществляется фильтрационный процесс, и в почечные канальцы профильтровывается плазма крови — так возникает первичная моча. В собирательных трубочках мозгового слоя происходит реабсорбция из первичной мочи воды, сахара и аминокислот. Число почечных канальцев у млекопитающих очень велико. В обеих почках их насчитывается у мыши 10 тыс., у кролика — 285 тыс. Для сравнения укажем, что в почках лягушки их 2 тыс., а у тритона — всего 400.

Рис. 180. Схема расположения чувствительных волос (вибрисс) на морде кролика. Пунктиром очерчена «осязательная зона»
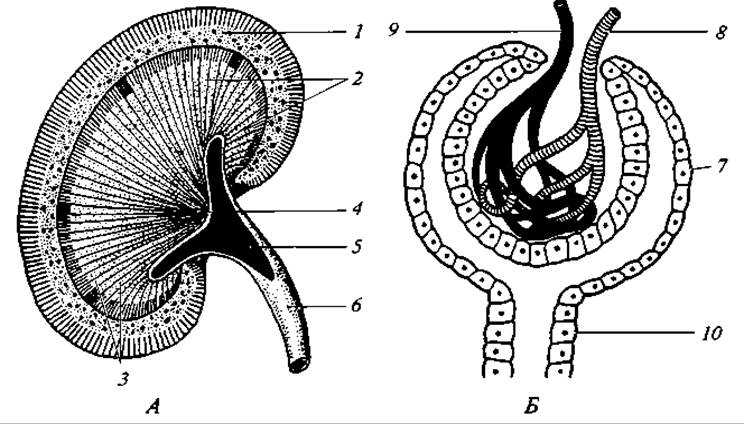
Рис. 181. Почка в продольном разрезе (А) и схема строения мальпигиева тельца (Б):
1 — корковый слой; 2 — мозговое вещество; 3 — пирамиды; 4 — почечные сосочки; 5 — лоханка; 6 — мочеточник; 7 — боуменова капсула; 8— входящая артерия, образующая клубочек; 9 — выходящая артерия; 10 — начало извитого канальца
Относительные размеры почек находятся в обратной зависимости от величины тела. Они велики у самых мелких и наоборот. Такая зависимость вполне понятна, если учесть, что у близких филогенетически и экологически видов интенсивность обмена веществ будет тем выше, чем меньше размеры тела. Хорошим, хотя и косвенным, подтверждением сказанному служит ясная корреляция между показателями относительной массы почки и сердца — органов, в одинаковой мере ответственных за уровень метаболизма.
Основной конечный продукт белкового обмена у млекопитающих (как и у рыб и амфибий) в отличие от рептилий и птиц не мочевая кислота, а мочевина. Если у птиц долевое значение мочевой кислоты равно 63 — 80 % и на долю мочевины приходится 1 — 10 %, то у млекопитающих соотношение обратное: мочевина — 68 — 91 %, мочевая кислота — 0,1 — 8 %.
Такой тип белкового обмена у млекопитающих, несомненно, возник в связи с наличием плаценты, через которую развивающийся эмбрион может получать в неограниченном количестве воду из крови матери. С другой стороны, через посредство плаценты (точнее, системы ее кровеносных сосудов) из развивающегося эмбриона могут также неограниченно выводиться токсические продукты белкового обмена.
Напомним, что мочевина значительно токсичнее мочевой кислоты, и для выведения мочи такой тип обмена требует очень большого расхода воды. Кроме того, это свидетельство близости млекопитающих амфибиям.
В мозговом слое находятся прямые собирательные канальцы, которые концентрируются в группы (пирамиды) и открываются на конце сосочков, выдающихся в почечную лоханку. От почечной лоханки отходит мочеточник, впадающий в мочевой пузырь. У однопроходных мочеточник впадает в мочеполовой синус, из которого он попадает в мочевой пузырь. Из мочевого пузыря моча выводится по самостоятельному мочеиспускательному каналу.
Выделительную функцию частично выполняют и потовые железы, через которые выводятся растворы солей и мочевины. Этим путем выводится не более 3 % азотистых продуктов белкового обмена. Органы воспроизведения. Половые железы самца — парные семенники — имеют характерную овальную форму (рис. 182, А). У однопроходных, некоторых насекомоядных и неполнозубых, у слонов и китообразных они в течение всей жизни находятся в полости тела. У большинства других зверей семенники первоначально располагаются в полости тела, но по мере полового созревания они опускаются вниз и попадают в особый, расположенный снаружи, мешочек — мошонку, сообщающуюся с полостью тела паховым каналом. К семеннику прилегает вытянутое по его оси зернистое тело — придаток семенника, морфологически представляющий клубок сильно извитых семявыносящих каналов семенника, гомологичный переднему отделу туловищной почки. От придатка отходит гомологичный вольфову каналу семяпровод, который впадает у корня полового члена в мочеполовой (мочеиспускательный и семя-извергательный) канал. Семяпроводы в своем нижнем отделе, перед впадением в мочеполовой канал, образуют парные компактные тела с ребристой поверхностью — семенные пузыри. У млекопитающих они представляют железы, секрет которых принимает участие в образовании жидкой части спермы; кроме того, он обладает клейкой консистенцией и благодаря этому служит, видимо, для предотвращения вытекания спермы из половых путей самки.
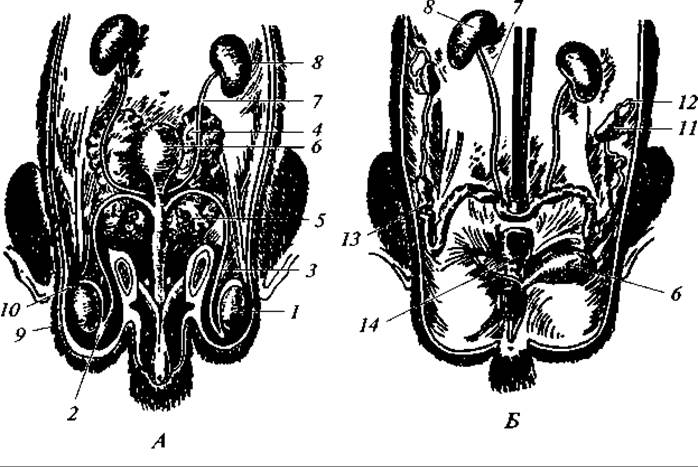
Рис. 182. Мочеполовые органы самца (А) и самки (Б) крысы:
1 — семенник; 2 — придаток семенника; 3 — семяпровод; 4 — семенные пузыри; 5 — предстательная железа; 6 — мочевой пузырь; 7 — мочеточник; 8 — почка; 9 — мошонка; 10 — паховый канал; 11 — яичник; 12 — фаллопиева труба; 13 — матка; 14 — влагалище
У основания полового члена лежит вторая парная предстательная железа, протоки которой впадают также в начальную часть мочеполового канала. Секрет предстательных желез — это основная часть жидкости, в которой плавают выделенные семенниками сперматозоиды. В итоге сперма, или эйякулят, представляет собой комбинацию жидкости, выделенной предстательной железой и семенными пузырями (и некоторыми другими железами), и самих сперматозоидов.
На нижней стороне совокупительного члена располагается упомянутый уже мочеполовой канал. Вверх и по бокам от этого канала лежат пещеристые тела, внутренние полости которых во время полового возбуждения наполняются кровью, в результате чего половой член становится упругим и увеличивается в размерах. У многих млекопитающих прочность полового члена обусловливается еще особой длинной костью, расположенной между пещеристыми телами. Таковы хищные, ластоногие, многие грызуны, некоторые летучие мыши и др.
Половые железы самки — парные яичники — всегда лежат в полости тела и прикреплены к спинной стороне брюшной полости брыжейками (рис. 182, Б). Парные яйцеводы, гомологичные мюллеровым каналам, открываются передними своими концами в полость тела в непосредственной близости от яичников. Здесь яйцеводы образуют широкие воронки. Верхний извитый отдел яйцеводов представляет фаллопиевы трубы. Далее идут расширенные отделы матки, которые открываются в непарный у большинства зверей отдел — влагалище. Последнее переходит в короткий мочеполовой канал, в который кроме влагалища открывается мочеиспускательный канал. На брюшной стороне мочеполового канала располагается небольшой вырост — клитор, обладающий пещеристыми телами и соответствующий пенису самца. Любопытно, что у некоторых видов в клиторе имеется кость.
Строение женских половых путей существенно различается у разных групп млекопитающих. Так, у однопроходных яйцеводы на всем протяжении парные и дифференцированы только на фаллопиевы трубы и рога матки, которые самостоятельными отверстиями открываются в мочеполовой синус. У сумчатых обособляется влагалище, но часто и оно остается парным. У плацентарных влагалище всегда непарное, а более верхние отделы яйцеводов в той или иной мере сохраняют парный характер. В простейшем случае матка парная, левый и правый ее отделы открываются во влагалище самостоятельными отверстиями. Такая матка называется двойной; она свойственна многим грызунам, некоторым неполнозубым. Рога матки могут быть соединены только в нижнем отделе — это двураздельная матка некоторых грызунов, летучих мышей, хищников. Слияние значительной части левого и правого рогов матки приводит к образованию двурогой матки хищных, китообразных, копытных. Наконец, у приматов, полуобезьян и некоторых летучих мышей непарная простая матка, и парными остаются только верхние отделы яйцеводов — фаллопиевы трубы. Плацента. Во время развития эмбриона в матке млекопитающих формируется крайне характерное образование, известное под названием детского места, или плаценты (рис. 183). Только у однопроходных плацента отсутствует. У сумчатых есть зачатки плаценты. Плацента возникает путем срастания наружной стенки аллантоиса с серозой, в результате чего формируется губчатое образование — хорион. Хорион образует выросты — ворсинки, которые соединяются или срастаются с разрыхленным участком эпителия матки. В этих местах кровеносные сосуды детского и материнского организмов сплетаются (но не сливаются), и таким образом устанавливается связь между кровяными руслами эмбриона и самки. В результате этого обеспечиваются газообмен в теле зародыша, его питание и удаление продуктов распада.
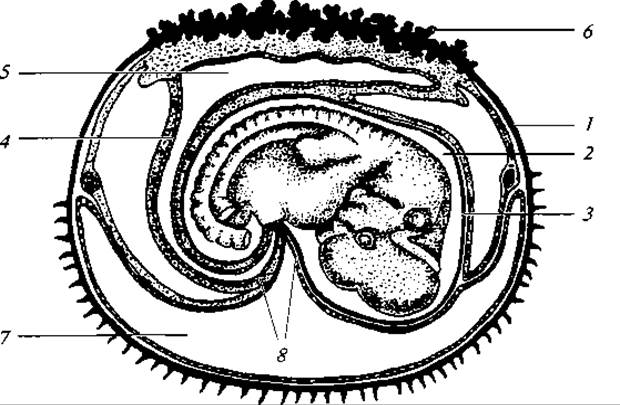
Рис. 183. Зародыш кролика в конце двенадцатого дня:
1 — серозная оболочка; 2 — амниотическая полость; 3 — амнион; 4 — аллантоис; 5 — полость аллантоиса; 6 — хорион; 7 — полость желточного мешка; 8 — пупочный канатик
Плацента свойственна уже сумчатым млекопитающим, хотя у них она еще примитивная; ворсинок в хорионе не образуется, и имеется, как и у яйцеживородящих низших позвоночных, пристеночная связь между кровеносными сосудами матки и желточного мешка (так называемая «желточная плацента»), у высших плацентарных зверей хорион всегда образует выросты — ворсинки, соединяющиеся со стенками матки. Характер расположения ворсинок различен у разных групп зверей. Основываясь на этом, выделяют три типа плаценты: диффузная, когда ворсинки распределяются равномерно по хориону (китообразные, многие копытные, полуобезьяны); дольчатая, когда ворсинки собраны в группы, распределенные по всей поверхности хориона (большинство жвачных); дискоидальная — ворсинки располагаются на ограниченном, дисковидном, участке хориона (насекомоядные, грызуны, обезьяны).
Общее число ныне живущих видов млекопитающих в мире более 4 000. В классе Млекопитающие различают два подкласса: Первозвери (Prototheria) с одним отрядом Однопроходные (Monotremata) и Настоящие звери (Theria) с двумя инфраклассами и многими отрядами.