Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимические основы жизнедеятельности организма человека
Биохимия углеводов
Внутриклеточный обмен углеводов
Внутриклеточный обмен углеводов включает процессы синтеза и распада гликогена в скелетных мышцах и печени, распад и окисление глюкозы с высвобождением энергии и новообразование глюкозы из веществ неуглеводной природы.
Биосинтез гликогена — лепонирование углеволов
Избыточное количество глюкозы, поступившей с кровью, откладывается про запас главным образом в печени и скелетных мышцах. Синтез и накопление гликогена называется депонированием углеводов. Гликоген является основным углеводным энергетическим резервом организма. От запасов его в скелетных мышцах и печени зависит длительность выполнения мышечной работы, поэтому в практике спорта используются специальные методы накопления гликогена в тканях.
Синтез гликогена из молекул глюкозы осуществляется с помощью гликогенсинтетазы и уридинтрифосфата (УТФ) как источника энергии. Этому процессу предшествует несколько реакций превращения глюкозы, представленных на рис. 62. Глюкоза с участием фермента гексокиназы и АТФ превращается в глюкозо-6-фосфат, который под воздействием фермента фосфоглюкомутазы переходит в глюкозо-1-фосфат. Из глюкозо-1-фосфата с участием УТФ образуется активная форма УДФ-глюкозы, которая под воздействием фермента гликогенсинтетазы присоединяется к последнему остатку гликогена:
![]()
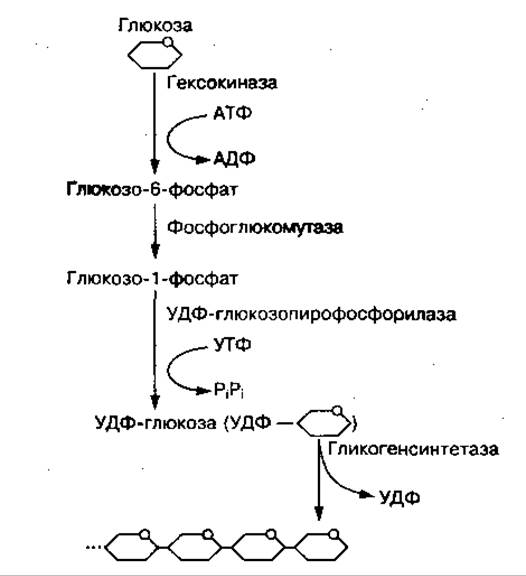
Рис. 62 Схема биосинтеза гликогена
Таким образом молекула гликогена увеличивается на один остаток глюкозы. Синтез гликогена с участием гликогенсинтетазы возможен только при наличии небольшого количества гликогена — так называемой затравки. С удлинением молекулы гликогена активность фермента увеличивается. Ветвление ее осуществляется ферментом трансгликогеназой.
Синтез гликогена требует затрат энергии АТФ, УТФ и пирофосфата. При удлинении молекулы гликогена только на один остаток глюкозы используется 41 кДж энергии. Поэтому этот процесс в тканях интенсивно протекает в аэробных условиях за счет АТФ, образующейся путем окислительного фосфорилирования. В печени он накапливается при усиленном питании, а в мышцах — после истощения его запасов, например после длительной физической нагрузки. Наибольшая скорость синтеза гликогена наблюдается в период отдыха на 30—40-й минуте после приема углеводной пищи. Это необходимо учитывать при построении режима питания во время соревнований, с тем чтобы усилившийся процесс синтеза не затормозил использование гликогена при мышечной деятельности.
Усиливается синтез гликогена под воздействием гормона поджелудочной железы — инсулина, который регулирует поступление глюкозы в ткани, а ингибируется адреналином, так как он угнетает активность фермента гликогенсинтетазы. Уровень гликогена в печени и других тканях может увеличиваться только до определенного предела, поскольку высокие концентрации его в тканях угнетают активность гликогенсинтетазы.
Распад гликогена — мобилизация углеволов
Процесс распада гликогена до молекул глюкозы называется мобилизацией углеводов. Расщепление гликогена происходит в основном путем фосфоролиза с участием фермента гликогенфосфорилазы и фосфорной кислоты (Н3РО4). От гликогена отщепляется молекула глюкозы в виде глюкозо-1-фосфата:
![]()
Образовавшийся глюкозо-1-фосфат быстро превращается в глюкозо-6-фосфат. В печени он расщепляется ферментами фосфатазами на свободную глюкозу и фосфорную кислоту. Молекулы свободной глюкозы легко поступают в кровь и используются многими тканями организма как энергетический субстрат. В скелетных мышцах такие фосфатазы отсутствуют, поэтому гликоген в них используется только для собственных нужд.
Скорость распада гликогена в мышцах зависит от их функциональной активности, а в печени — от уровня глюкозы в крови. При мышечной деятельности скорость мобилизации гликогена в печени зависит от интенсивности выполняемой нагрузки: при умеренной работе она возрастает в 2—3 раза, а при интенсивной — в 7—10 раз по сравнению с состоянием покоя.
Распад гликогена в печени продолжается и в период отдыха. Образующаяся глюкоза способствует восстановлению запасов гликогена в сердечной и скелетной мышцах, т. е. происходит перераспределение углеводов между отдельными тканями.
Гликолиз
Извлечение метаболической энергии из углеводов происходит почти во всех клетках организма человека и включает две основные фазы — бескислородное (анаэробное) окисление, которое протекает в цитозоле преимущественно скелетных мышц и называется гликолизом, и кислородное (аэробное) окисление, протекающее в митохондриях на ферментах цикла лимонной кислоты и дыхательной цепи.
Гликолиз — это постепенный распад молекулы глюкозы или гликогена (гликогенолиз) до двух молекул пировиноградной кислоты, которая в анаэробных условиях превращается в молочную кислоту. Он включает десять химических реакций, представленных на рис. 63. Этот процесс можно разделить на две основные стадии — подготовительную и окислительную. В подготовительной стадии молекула глюкозы постепенно распадается до двух молекул 3-фосфоглицеринового альдегида, при этом используется две молекулы АТФ. В окислительной стадии происходит дальнейшее их окисление с образованием пирувата и четырех молекул АТФ. Начинается гликолиз с активации молекулы глюкозы в присутствии АТФ с образованием глюкозо-6-фосфата или фосфоролиза гликогена с отщеплением глюкозо-1-фосфата. Реакция фосфорилирования глюкозы катализируется ферментом гексокиназой и требует наличия ионов Мg2+. Гексокиназа — это регуляторный аллостерический фермент, активность которого зависит от содержания АТФ в клетке. При низкой концентрации АТФ фермент активен, а при высокой ее концентрации — не активен и процесс гликолиза «выключается», так как энергия в данный момент не используется. Далее глюкозо-6-фосфат превращается в фруктозо-6-фосфат с участием фермента глюкозофосфатизомеразы. Фруктозо-6-фосфат фосфорилируется с использованием энергии АТФ, в результате чего образуется фруктозо-1,6-дифосфат. Реакция катализируется ферментом фосфофруктокиназой (ФФК).
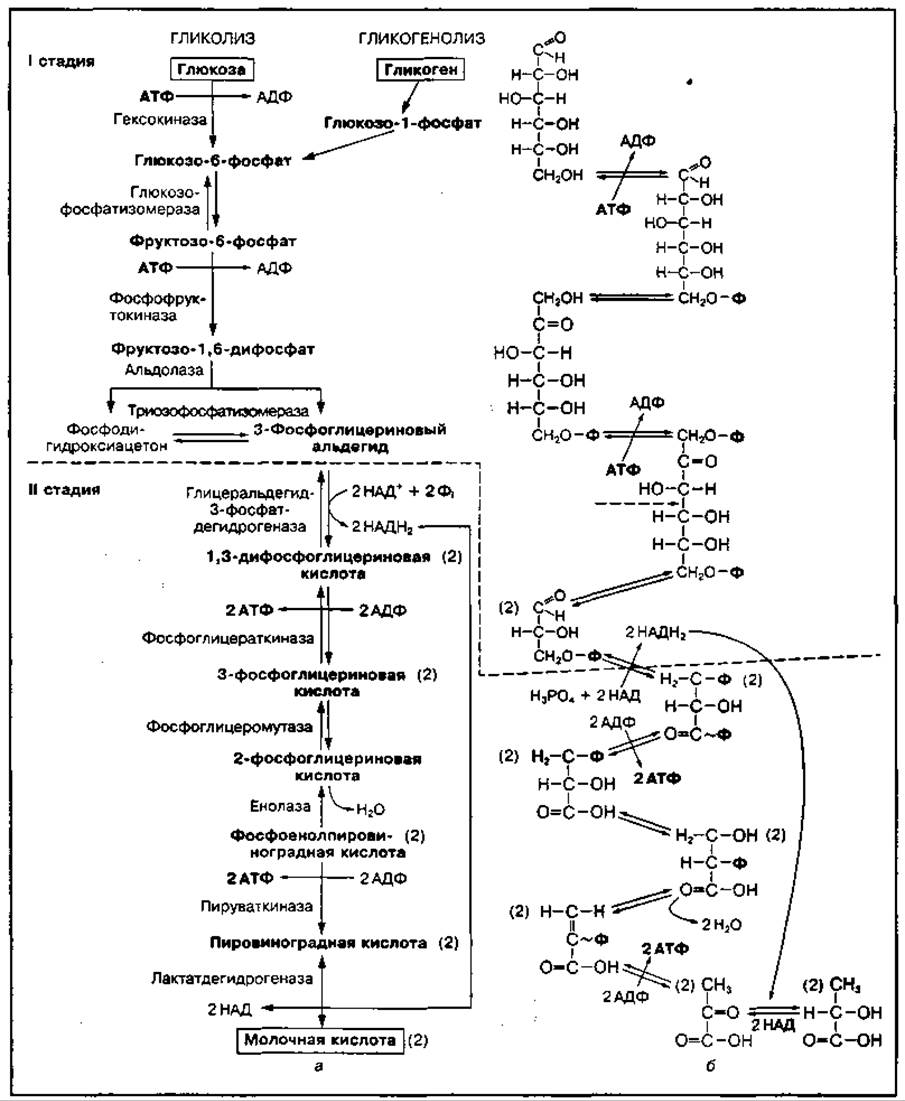
Рис. 63 Схема гликолиза (а) и его химические реакции (б)
Фосфофруктокиназа является ключевым аллостерическим ферментом, регулирующим скорость гликолиза. Его активность зависит от концентрации АТФ и других метаболитов (молочной кислоты, цитрата), которые влияют на его активность. Так, в мышцах в состоянии покоя концентрация АТФ относительно высокая и процесс гликолиза не активен. Во время работы мышцы интенсивно расходуется АТФ, что повышает активность ФФК и приводит к усилению гликолиза. Однако накопление молочной кислоты — конечного продукта анаэробного гликолиза — ингибирует этот фермент и скорость гликолиза.
Первый этап гликолиза завершает реакция расщепления фруктозо-1,6-дифосфата на две триозы — фосфоглицериновый альдегид и фосфодиоксиацетон под воздействием фермента альдолазы. Образовавшиеся триозы являются изомерами и способны взаимопревращаться. В последующие реакции гликолиза вступают две модели 3-фосфоглицеринового альдегида.
Окислительная стадия начинается с окисления 3-фосфоглицеринового альдегида при участии дегидрогеназы, содержащей кофермент НАД, и фосфорной кислоты. Кофермент НАД в этой реакции присоединяет водород и превращается в НАДН2. В аэробных условиях НАДН2 может передать водород на кислород с образованием 3АТФ. Образовавшаяся 1,3-дифосфоглицериновая кислота содержит макроэргетическую связь и способна вступить в реакцию перефосфорилирования с АДФ, ведущую к образованию АТФ и 3-фосфоглицериновой кислоты. Такой процесс образования АТФ называется субстратным фосфорилированием. Он катализируется ферментом фосфоглицераткиназой.
3-Фосфоглицериновая кислота под влиянием фермента фосфоглицеромутазы превращается в 2-фосфоглицериновую кислоту. Последняя при участии фермента энолазы теряет молекулу воды и превращается в фосфоэнолпировиноградную кислоту. В результате внутримолекулярного окислительно-восстановительного процесса у второго углеродного атома этой кислоты образуется макроэргическая связь, при разрыве которой с участием фермента пируваткиназы происходит перенос фосфорного остатка от фосфоэнолпировиноградной кислоты на АДФ (второе субстратное фосфорилирование), а также образование двух молекул пировиноградной кислоты и двух молекул АТФ.
Гликолиз в анаэробных условиях завершается реакцией восстановления пировиноградной кислоты до молочной под воздействием фермента лактатдегидрогеназы. Источником водорода служат молекулы НАДН2, образующиеся при окислении 3-фосфоглицеринового альдегида. Таким образом, конечным продуктом анаэробного гликолиза является молочная кислота. В аэробных условиях пировиноградная кислота не превращается в молочную и окисляется далее в цикле лимонной кислоты до конечных продуктов обмена. Суммарное уравнение процесса гликолиза можно представить в виде
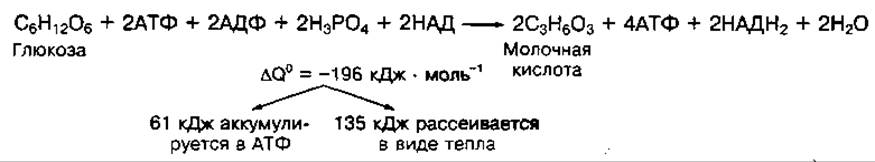
В процессе гликолиза постепенно высвобождается 196 кДж энергии. Большая часть ее рассеивается в виде тепла (135 кДж), а меньшая — накапливается в макроэргических связях двух молекул АТФ. Эффективность запасания энергии в форме АТФ при гликолизе составляет 40 %. Основная часть энергии, аккумулированной в молекуле глюкозы (2880 кДж), остается в продукте гликолиза — двух молекулах молочной кислоты и может высвобождаться только при их аэробном окислении. В гликолизе образуются многие вещества, необходимые для пластических процессов в клетках. Особенно много при этом накапливается молочной кислоты, которая быстро диффундирует из скелетных мышц в кровь и влияет на кислотно-основное состояние организма. Уровень молочной кислоты в крови только до некоторой степени отражает интенсивность гликолиза в мышцах, поскольку кислота частично метаболизирует в них. В норме концентрация молочной кислоты в крови находится в пределах 1— 1,5 ммоль ∙ л-1.
Молочная кислота в водной среде диссоциирует на протон водорода (Н+) и анион кислотного остатка:
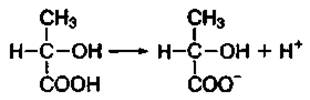
Анион кислотного остатка молочной кислоты в водной среде способен взаимодействовать с катионами металлов (Na+, К+) и образовывать соли — лактаты. Поэтому часто молочную кислоту называют лактат.
Гликолиз и гликогенолиз играют важную роль при мышечной деятельности анаэробного характера. Они обеспечивают энергией интенсивную работу в пределах от 30 с до 2—5 мин.
Молочная кислота образуется в мышцах с наибольшей скоростью в течение 40—45 с интенсивной физической нагрузки за счет максимального включения анаэробного гликолиза. Уровень молочной кислоты при этом повышается в 4—5 раз и после напряженной работы в течение 1—5 мин может достигать 10 ммоль ∙ л-1 и более. Это приводит к закислению среды организма (ацидозу). Если емкость буферных систем исчерпывается, pH крови может изменяться от 7,34 до 7,0 или даже 6,8 при истощающих нагрузках. Такое закисление среды организма влияет на функции нервной системы и скелетных мышц, приводит к снижению работоспособности и развитию утомления. Поскольку концентрация молочной кислоты в крови зависит от интенсивности гликолиза в мышцах, а последний — от интенсивности выполняемых физических нагрузок и уровня тренированности организма, показатель содержания молочной кислоты в крови широко используется в биохимическом контроле функционального состояния спортсмена (см. главу 24).
После прекращения работы около 55—70 % молочной кислоты используется тканями, в том числе мышцами, как источник энергии, около 5—7 % выводится с мочой, а остальная часть используется в печени для новообразования глюкозы и восполнения гликогена в мышцах. Выведение молочной кислоты из мышц, ее окисление после прекращения работы ускоряется при активном отдыхе.
Аэробное окисление углеводов
Аэробное окисление глюкозы — это многостадийный процесс распада ее молекулы до конечных продуктов обмена СО2 и Н2О с образованием 38 молекул АТФ и выделением тепловой энергии (рис. 64). Протекает оно при участии кислорода, который доставляется в ткани с участием белка гемоглобина. Этот процесс окисления углеводов — один из главных механизмов образования АТФ в тканях организма. Он включает такие основные стадии:
✵ гликолитический распад молекулы глюкозы до двух молекул пировиноградной кислоты (ПВК);
✵ превращение ПВК в ацетил-КоА;
✵ окисление ацетил-КоА в цикле лимонной кислоты и на дыхательной цепи.
Процесс распада молекулы глюкозы до пировиноградной кислоты протекает одинаково в анаэробных и аэробных условиях, как описано выше (см. «Гликолиз»). Образовавшаяся в гликолитической стадии пировиноградная кислота подвергается далее окислительному декарбоксилированию, в результате чего образуются макроэргическое вещество ацетил-КоА, восстановленная форма НАДН2 и одна молекула СО2. Если в процессе гликолиза образовалась молочная кислота, то в аэробных условиях она превращается в пировиноградную кислоту.
Ацетил-КоА далее включается в цикл лимонной кислоты, где расщепляется до СО2 и Н2О. Вода образуется на системе дыхательных ферментов при взаимодействии водорода, образовавшегося в реакциях биологического окисления, с атомарным кислородом вдыхаемого воздуха. Суммарное уравнение аэробного окисления молекулы глюкозы можно представить таким образом:
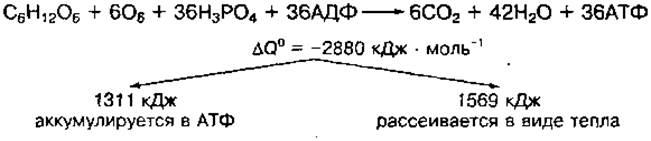
Химические реакции окисления молекулы ацетил-КоА в цикле лимонной кислоты рассмотрены в главе 3 (см. рис. 18).
Энергетическая эффективность аэробного окисления молекулы глюкозы
При полном окислении одной молекулы глюкозы до СО2 и Н2О энергия накапливается в виде 10 НАДН2 (2 из них образуются в гликолизе, 2 — при превращении ПВК в ацетил-КоА и 6 — в цикле лимонной кислоты), а также 2ФАДН2, 2 молекулы ГТФ, равноценные АТФ и 2АТФ в процессе гликолиза (см. рис. 64). Передача водорода по системе дыхательных передатчиков от НАДН2 на кислород сопряжена с образованием трех молекул АТФ, а от ФАДН2 — двух молекул АТФ. Следовательно, из 10 НАДН2 образуется 30 АТФ, а из 2ФАДН2 — 4 АТФ. Суммарный выход АТФ на одну молекулу глюкозы составляет 38 АТФ (табл. 15).
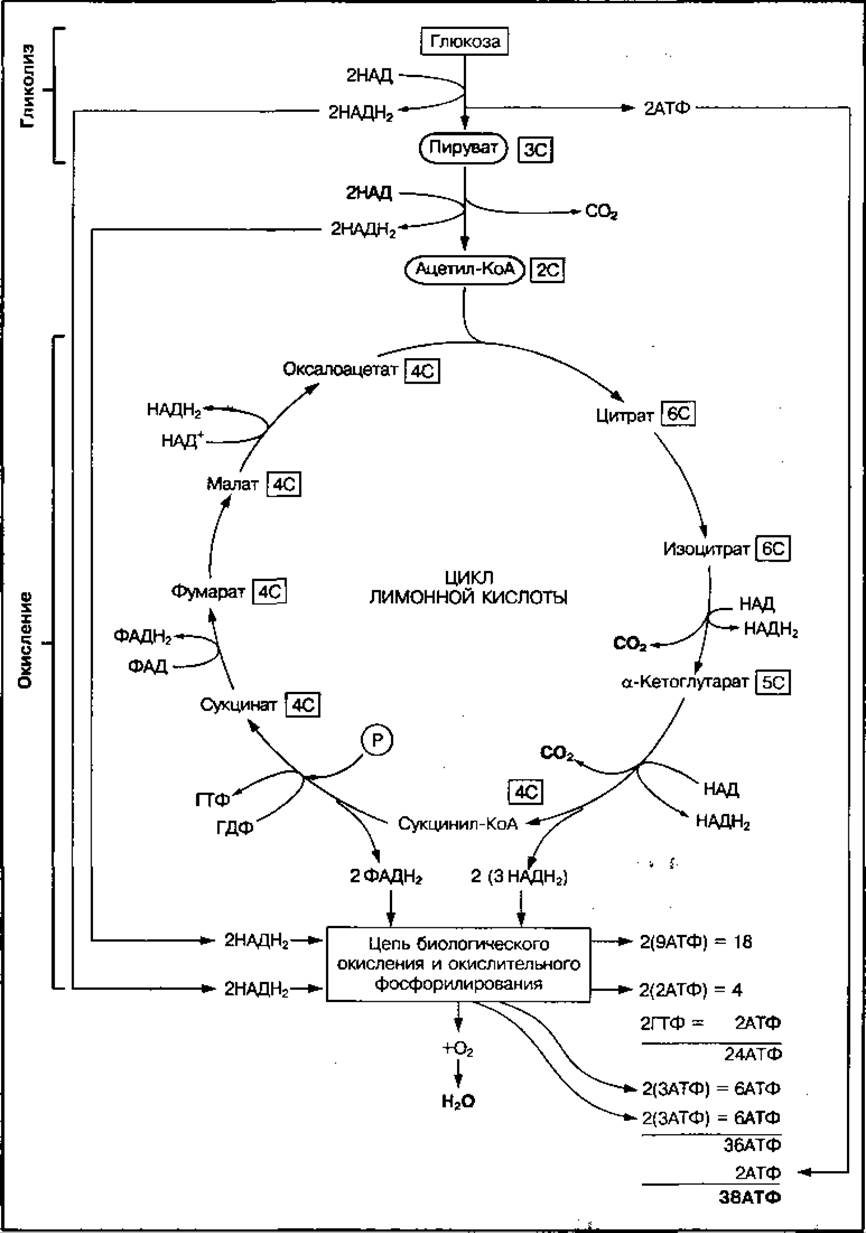
Рис. 64 Схема аэробного окисления глюкозы и энергетическая эффективность этого процесса
ТАБЛИЦА 15 Накопление энергии на отдельных стадиях окисления молекулы глюкозы
Стадия окисления |
Аккумуляция энергии |
Выход АТФ |
Гликолиз |
4АТФ образуется 2АТФ используется 2НАДН2 |
2АТФ накапливается 6АТФ |
Превращение ПВК в ацетил-КоА |
2НАДН2 |
6АТФ |
В цикле лимонной кислоты и окислительном фосфорилировании |
6НАДН2 2ФАДН2 2ГТФ |
18АТФ 4АТФ 2АТФ |
Общий выход АТФ |
38АТФ |
Однако в мышечной и нервной тканях 2 молекулы НАДН2, которые образуются в цитоплазме в процессе гликолиза, сами в митохондрии не поступают, а передают водород на переносчик ФАД, поэтому в дыхательную цепь водород уже передается от 2 ФАДН2, что сопровождается образованием не 6 АТФ, а только 4 АТФ. Поэтому в скелетных мышцах при полном окислении молекулы глюкозы образуется 36 АТФ.
Аэробный метаболизм глюкозы по накоплению АТФ в 19 (18) раз более эффективен, чем анаэробный. Он имеет большой коэффициент полезного действия (около 45 %), так как из 2880 кДж свободной энергии окисления глюкозы 1311 кДж аккумулируется в АТФ. Аэробное окисление углеводов — основной механизм энергообеспечения аэробной мышечной работы в течение нескольких часов.
Пентозный цикл окисления углеводов
В некоторых тканях организма (печени, эритроцитах, жировой ткани) возможен и другой аэробный путь прямого окисления углеводов, который называется пентозным циклом. В этом цикле накапливается энергия в виде НАДФН2, которая используется в биосинтетических процессах, а также образуются пентозы (рибоза и др.), необходимые для синтеза нуклеотидов (АТФ, НАД, ФАД, нуклеиновые кислоты), и глицериновый альдегид, который может превращаться в пировиноградную кислоту или включаться в гликолиз. Суммарное уравнение пентозного цикла имеет следующий вид:
6 глюкозо-6-фосфат + 12НАДФ + 7Н2О →
→ 5 глюкозо-6-фосфат + 12НАДФН2 + 6СО2 + Н3РО4
В процессе этого цикла молекула глюкозо-6-фосфата полностью окисляется до СО2. Основные четыре механизма пентозного цикла окисления углеводов представлены на рис. 65.
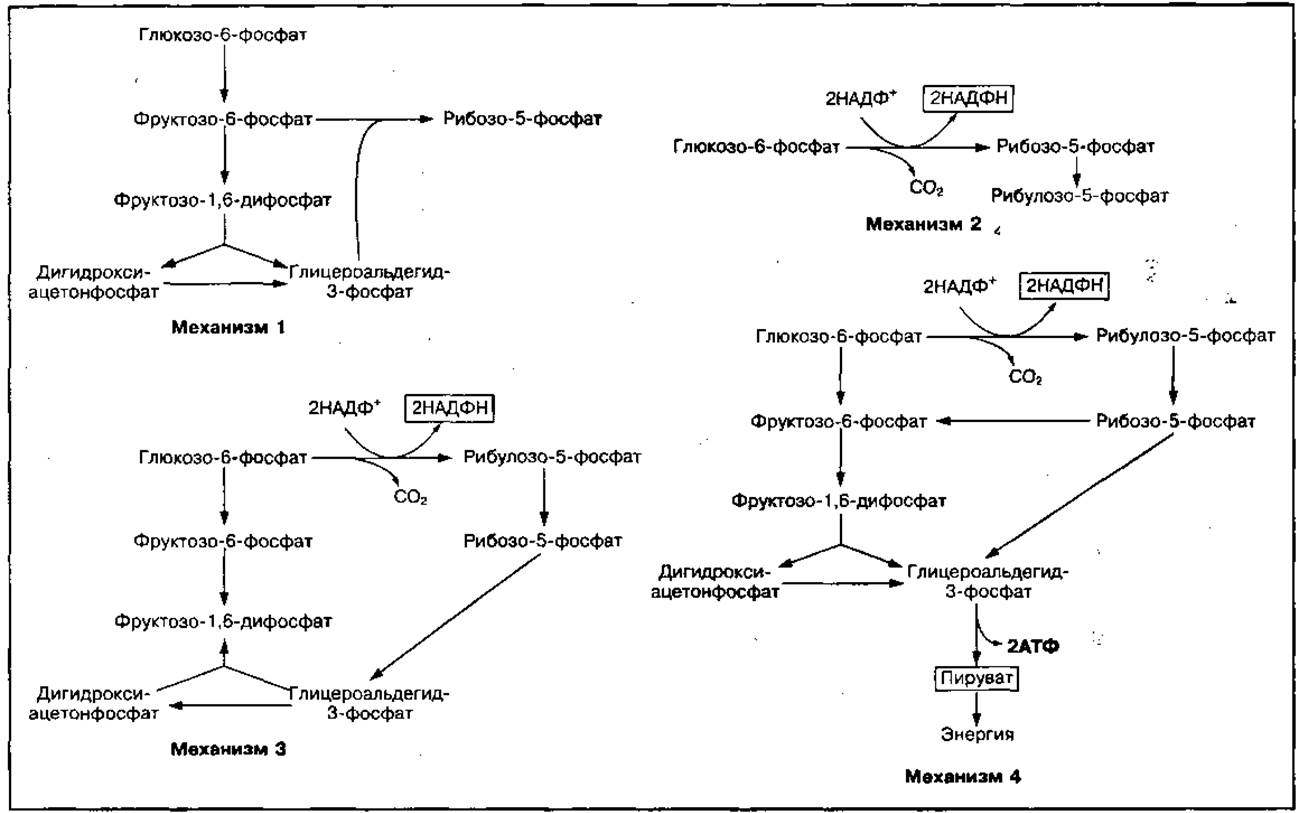
Рис. 65 Четыре механизма пентозофосфатного пути окисления углеводов
Глюконеогенез
Процесс новообразования глюкозы в тканях организма из веществ неуглеводной природы называется глюконеогенезом. Глюкоза может синтезироваться из пировиноградной и молочной кислот, а также из ацетил-КоА, глицерина и аминокислот (рис. 66). Все они, кроме глицерина, проходят через стадию образования пировиноградной кислоты. Многие реакции глюконеогенеза представляют собой обращение соответствующих реакций, имеющих место в процессе гликолиза. Однако имеются дополнительные обходные реакции, например образование фосфоенолпировиноградной и пировиноградной кислот.
Процесс новообразования глюкозы активно протекает в печени, почках, а при физических нагрузках — и в скелетных мышцах. Благодаря этому процессу предотвращается резкое снижение уровня глюкозы в крови и гликогена в печени (например, при длительной мышечной работе).
Важное значение в восстановлении уровня глюкозы и гликогена в печени и скелетных мышцах имеет процесс превращения молочной кислоты в глюкозу, который называется циклом Кори. Постепенное превращение многих аминокислот через аминокислоту аланин в молекулы глюкозы обеспечивает также другой цикл — глюкозоаланиновый (рис. 67).
Особенность этих циклов заключается в том, что молочная кислота и аланин, образуясь в скелетных мышцах, кровью транспортируются в печень, где превращаются в глюкозу. Это энергозависимый процесс. Из печени глюкоза поступает в кровь и снова используется мышцами для восстановления запасов гликогена. Данный процесс имеет существенное значение в организме при мышечной деятельности. Благодаря ему предотвращается резкое снижение запасов гликогена в мышцах и уровня глюкозы в крови.