Биохимические основы жизнедеятельности организма человека - Волков Н.И., Несен Э.Н. 2000
Биохимия спорта
Биохимические изменения в организме при выполнении упражнений различной мощности и продолжительности
Транспорт кислорода к работающим мышцам и его потребление при мышечной деятельности
Доставка кислорола к мышцам
Скорость доставки кислорода к тканям — один из важнейших факторов, влияющих на энергообеспечение мышц, так как скорость ресинтеза АТФ в митохондриях скелетных мышц, где образуется около 90 % всей необходимой энергии, находится в определенной зависимости от концентрации или напряжения кислорода в клетке. На рис. 140 показана зависимость скорости ресинтеза АТФ, обеспечиваемая за счет различных источников энергии от напряжения кислорода в клетке. При низком уровне метаболизма в клетке, что имеет место в покоящейся мышце, изменения скорости доставки О2 в ткани не влияют на скорость ресинтеза АТФ (зона насыщения). Однако, когда напряжение кислорода (рО2) в клетке ниже некоторого критического уровня, то поддержание скорости ресинтеза АТФ возможно только за счет адаптивных сдвигов внутриклеточного метаболизма, что неизбежно требует увеличения скорости доставки О2 к мышцам и его потребления митохондриями. Максимальная скорость потребления О2 митохондриями скелетных мышц может быть поддержана только до определенного критического значения рО2 в клетке, составляющего 0,5—3,5 мм рт. ст. Если уровень метаболической активности при мышечной работе превысит значение максимально возможного усиления аэробного ресинтеза АТФ, то возрастающая потребность в энергии может быть компенсирована за счет анаэробного ресинтеза АТФ. Однако диапазон анаэробной метаболической компенсации весьма узок и дальнейшее увеличение скорости ресинтеза АТФ в работающей мышце, как и функционирование мышц, становится невозможным. Диапазоны метаболической активности, в пределах которых доставка О2 недостаточна для поддержания необходимого уровня ресинтеза АТФ, обычно обозначаются как гипоксические состояния (т. е. состояния кислородной недостаточности) различной тяжести.
Для поддержания напряжения О2 в митохондриях на уровне выше критического значения, при котором еще сохраняются условия для адаптивной регуляции клеточного обмена, напряжение О2 на наружной клеточной мембране должно составлять не менее 15—20 мм рт. ст. Для его поддержания и нормального функционирования мышц напряжение кислорода в артериолах, доставляющих кровь непосредственно к работающим мышцам, должно составлять около 40 мм рт. ст., а в магистральных артериях — 80—90 мм рт. ст. В легочных альвеолах, где осуществляется газообмен между кровью и атмосферным воздухом, напряжение О2 составляет примерно 110 мм рт. ст., во вдыхаемом воздухе — 150 мм рт. ст. Как видно из рис. 141, на котором указаны величины критического напряжения О2 на различных этапах транспорта кислорода в организме, обозначенные в виде нисходящих ступеней с постепенно понижающимися уровнями рО2 и скоростью доставки О2 к тканям, "кислородный каскад" обеспечивает постоянную накачку О2 в работающие ткани в соответствии с их метаболической потребностью. Сохранение критического напряжения кислорода на каждом из обозначенных на схеме этапе транспорта кислорода к тканям обеспечивается совокупной деятельностью различных физиологических систем организма.
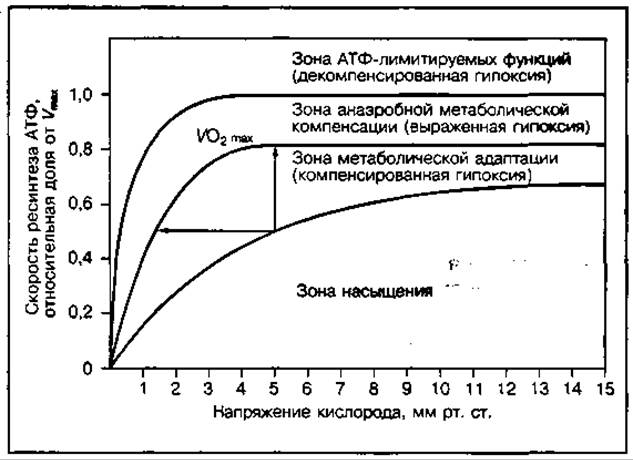
Рис. 140 Изменение скорости ресинтеза АТФ в аэробном и анаэробном процессах в зависимости от внутриклеточного напряжения кислорода
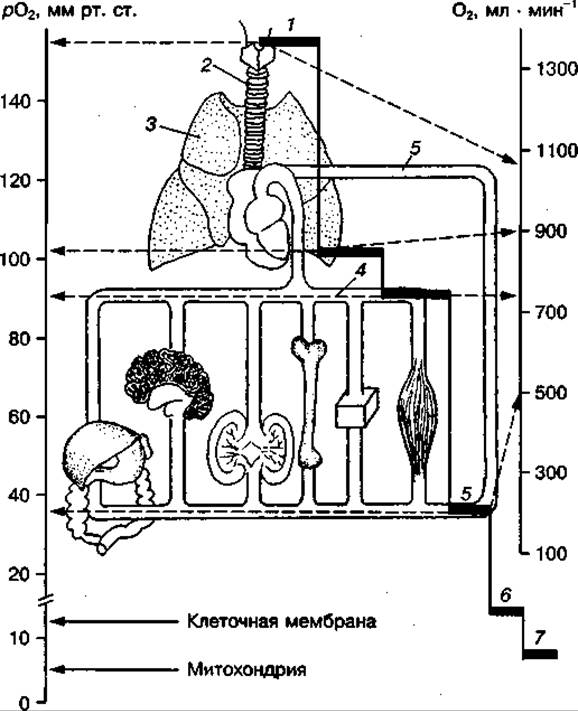
Рис. 141 «Кислородный каскад» организма:
1 — вдыхаемый воздух; 2 — дыхательные пути; 3 — легкие (альвеолы); 4 — артериальная кровь; 5 — смешанная венозная кровь; 6 — на наружной клеточной мембране; 7 — в митохондриях скелетных мышц
Кислород, содержащийся во вдыхаемом воздухе, диффундирует в кровь через стенки легочных альвеол и кровеносных капилляров вследствие разницы парциального давления О2 в альвеолярном воздухе и крови. Только небольшая часть поступающего в кровь кислорода растворяется в плазме (около 0,3 мм О2 на 100 мл крови); большая же часть связывается в эритроцитах с гемоглобином. При нормальных условиях каждый 1 г гемоглобина крови может связать 1,34 мл О2. В 100 мл крови взрослого человека содержится около 14—16 г гемоглобина, поэтому кислородная емкость крови, т. е. общее количество связанного ею кислорода, при полном насыщении гемоглобина может составлять 21—22 мл О2 на 100 мл крови.
На способность гемоглобина связывать кислород влияет температура крови и концентрация водородных ионов в ней: чем ниже температура и выше pH, тем больше кислорода может быть связано гемоглобином. Выделение СО2 из крови в выдыхаемый воздух способствует подщелачиванию крови и насыщению гемоглобина кислородом. В молекуле гемоглобина содержится 4 гема, которые могут связывать 4 молекулы О2, при этом взаимодействие первого гема с кислородом увеличивает сродство гемоглобина к кислороду и скорость присоединения последующих молекул О2.
Обогащенная кислородом кровь поступает в большой круг кровообращения. Сердце в состоянии покоя перекачивает ежеминутно 5—6 л крови, а значит, переносит от легких к тканям 250—300 мл кислорода в минуту. Во время работы объем крови возрастает до 30—40 л ⋅ мин-1, а количество переносимого кровью кислорода — до 5—6 л ⋅ мин-1.
Концентрация свободного кислорода в капиллярах тканей выше, чем во внутриклеточном пространстве, поэтому в них происходит освобождение кислорода от гемоглобина и диффузия его в клетки. Увеличение содержания СО2 и кислых продуктов обмена, а также местное повышение температуры крови в капиллярах тканей усиливают распад оксигемоглобина и освобождение кислорода.
В клетках мышц кислородный обмен осуществляется при участии белка миоглобина, имеющего сходную с гемоглобином структуру. Миоглобин переносит кислород к митохондриям, где протекают окислительные процессы, и частично депонирует его. Он обладает большим химическим сродством к кислороду, чем гемоглобин (рис. 142), что обеспечивает более полное использование мышцами кислорода, поставляемого кровью.
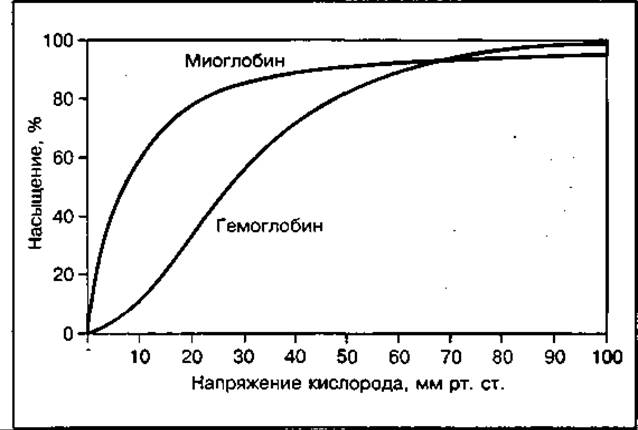
Рис. 142 Кривые диссоциации гемоглобина и миоглобина
Потребление кислорода мышцами
При переходе от состояния покоя к интенсивной мышечной деятельности потребность в кислороде возрастает во много раз, однако сразу она не может быть удовлетворена. Необходимо время, чтобы усилилась деятельность систем дыхания и кровообращения и чтобы кровь, обогащенная кислородом, могла дойти до работающих мышц. По мере усиления активности этих систем постепенно увеличивается потребление кислорода в работающих мышцах. Если при работе ЧСС превышает 150 уд ⋅ мин-1, то скорость потребления О2 возрастает до тех пор, пока не наступит истинное устойчивое состояние метаболических процессов, при котором потребление О2 в данный момент времени точно соответствует потребности организма в нем.
Уровень потребления О2 в устойчивом состоянии зависит от мощности выполняемого упражнения (рис. 143). При интенсивной работе мощностью более 200 Вт (ЧСС 150—180 уд ⋅ мин-1) устойчивое состояние не устанавливается и потребление О2 может возрастать до конца работы либо до достижения максимально возможного уровня. В последнем случае может наблюдаться "ложное устойчивое состояние", когда потребление О2 некоторое время (6—10 мин) поддерживается на максимальном уровне не потому, что потребность организма в кислороде полностью удовлетворяется, а потому, что исчерпаны возможности сердечно-сосудистой системы доставлять его к тканям. Эта система существенно ограничивает доставку О2 к мышцам. Однако основным лимитирующим фактором на уровне мышечных волокон является способность митохондрий утилизировать кислород и способность окислительных ферментов использовать его в работающих мышцах. Максимальный уровень потребления О2 не может поддерживаться долго: во время длительной работы он снижается из-за утомления.
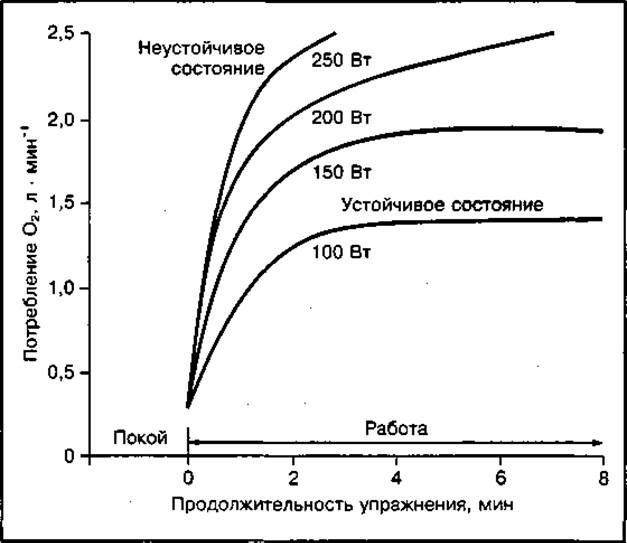
Рис. 143 Потребление кислорода при выполнении упражнений различной интенсивности и продолжительности
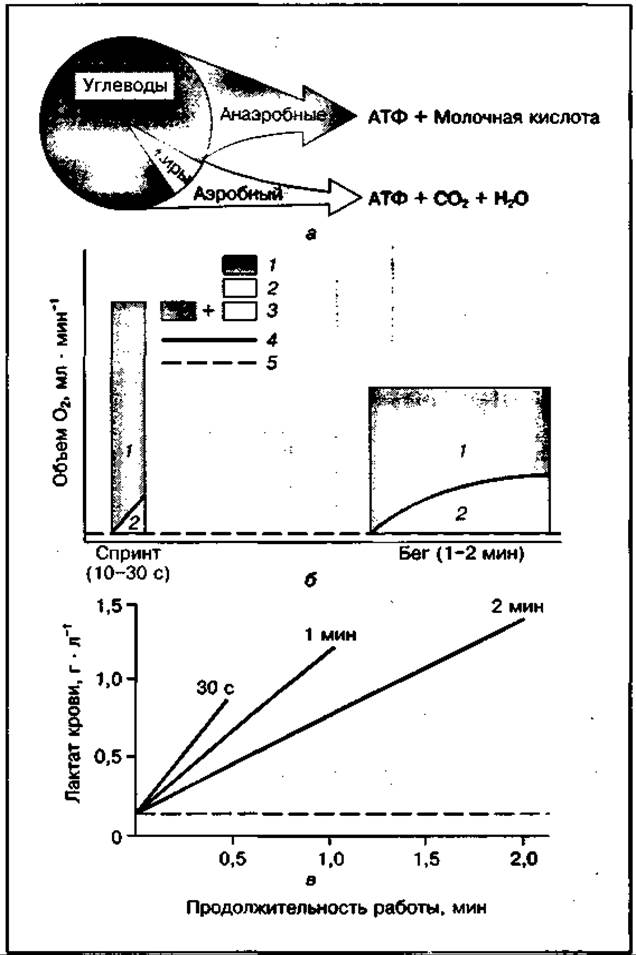
Рис. 144 Главные механизмы энергообразования (а), кислородный приход и кислородный дефицит (б), накопление лактата в крови (в) при кратковременной работе максимальной интенсивности: 1 — О2-дефицит; 2 — О2-потребление (приход); 3 — О2-запрос; 4 — потребление О2 в течение работы; 5 — потребление О2 в период отдыха
Количество кислорода, необходимое организму для полного удовлетворения энергетических потребностей за счет аэробных процессов, называется кислородным запросом работы. При интенсивной работе реальное потребление кислорода — кислородный приход — составляет только небольшую часть кислородного запроса. Разность между кислородным запросом работы и реально потребляемым кислородом составляет кислородный дефицит организма. В условиях кислородного дефицита активируются анаэробные реакции ресинтеза АТФ, что приводит к накоплению в организме недоокисленных продуктов анаэробного распада (рис. 144).
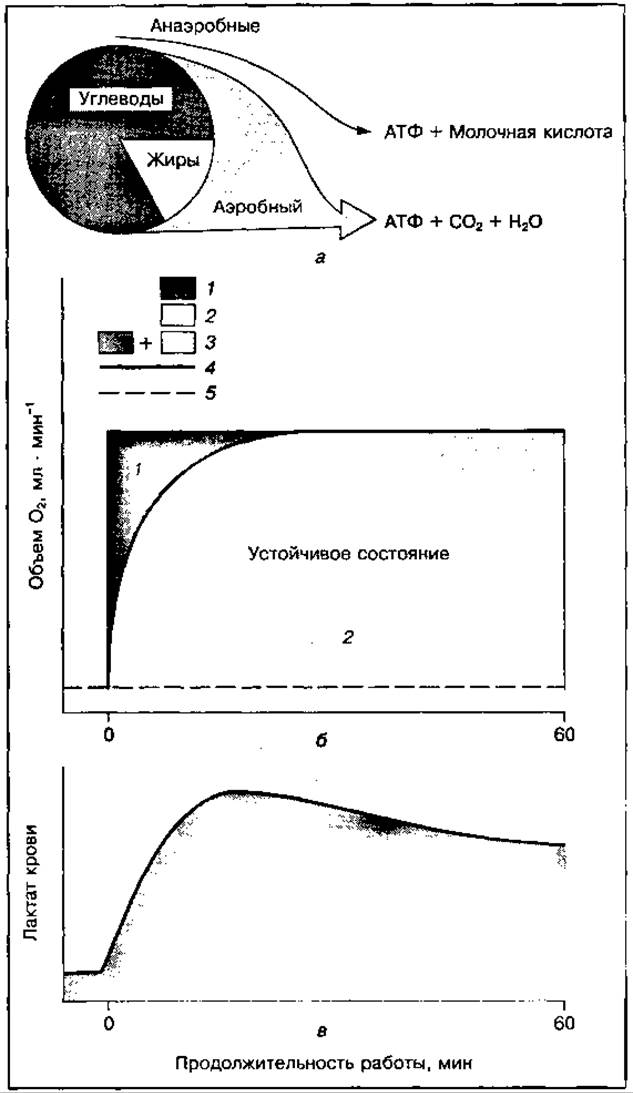
Рис. 145 Главные механизмы энергообразования (а), кислородный приход и кислородный дефицит (б), накопление лактата в крови (в) при длительной работе. Обозначения те же, что и на рис. 144
При работе в устойчивом состоянии часть анаэробных метаболитов может окисляться за счет усиления аэробных реакций в процессе работы, а другая их часть устраняется после работы. При работе, когда не устанавливается устойчивое состояние или при «ложном» устойчивом состоянии, количество недоокисленных продуктов постоянно увеличивается и устраняются они только в восстановительном периоде (рис. 145). Для восстановления энергетических источников и окисления недоокисленных продуктов требуется дополнительное количество кислорода, поэтому некоторое время после окончания работы потребление его продолжает оставаться повышенным по сравнению с уровнем покоя. Этот излишек кислородного потребления в период восстановления получил название «кислородный долг". Кислородный долг всегда больше кислородного дефицита, и чем больше интенсивность и продолжительность работы, тем значительнее это различие (рис. 146).
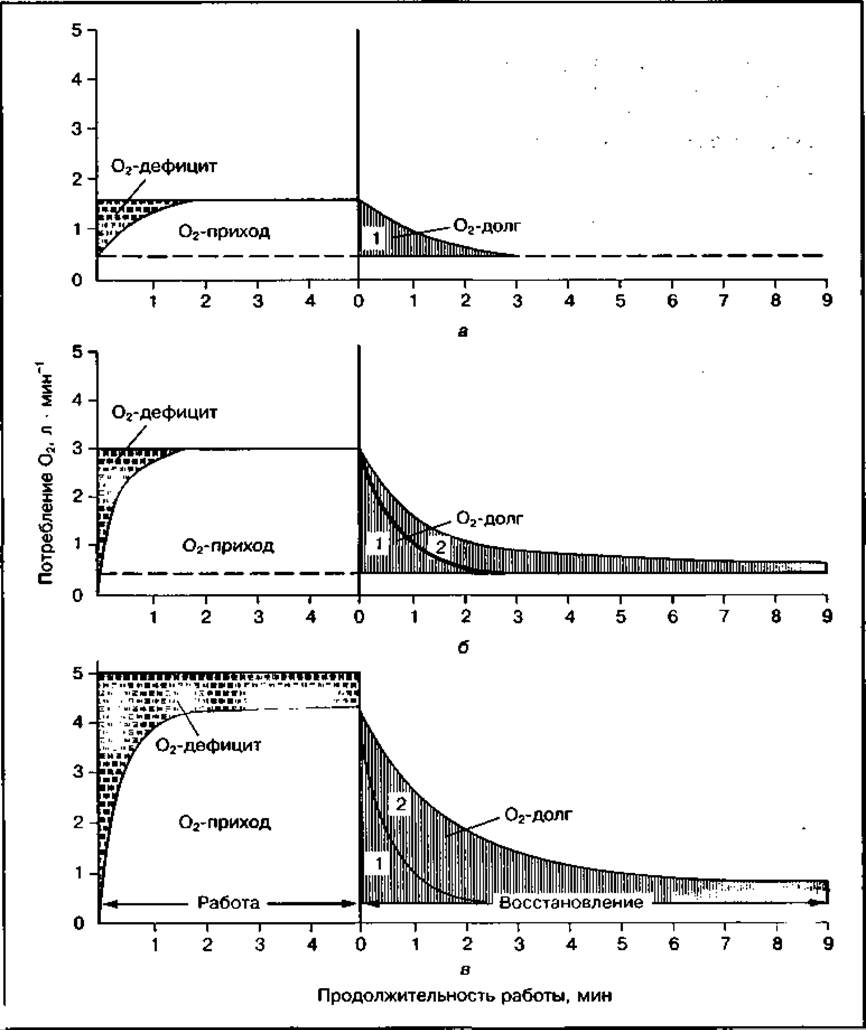
Рис. 146 Кислородный приход, кислородный дефицит и кислородный долг при работе разной интенсивности — легкой (а), тяжелой (б), истощающей (в), быстрый (1) и медленный (2) компоненты О2-долга
В период восстановления после мышечной работы, когда в организме имеется достаточное количество субстратов биологического окисления и поставка кислорода к митохондриям клеток не ограничена, уровень кислородного потребления зависит от количества свободной АТФ, осуществляющей дыхательный контроль в митохондриях. Субстратами окислительных энергетических превращений являются накопившиеся во время работы анаэробные метаболиты: молочная кислота, янтарная кислота, а-глицеро- фосфат, глюкоза, а на поздних стадиях восстановления — и жирные кислоты. Источником АДФ являются энергопотребляющие процессы, в первую очередь ресинтез КрФ из креатина, восстановление запасов гликогена и глюкозы, восстановление нарушенной во время работы структуры клеточных мембран, функционирование дыхательной и сердечно-сосудистой систем, активность которых некоторое время после работы сохраняется повышенной.
Часть потребленного в период отдыха кислорода используется для восстановления миоглобинового и гемоглобинового запаса. При работе, вызывающей утомление, эффективность использования кислорода снижается вследствие частичного разобщения процессов окисления энергетических субстратов и образования АТФ. Разобщение может сохраняться и после работы, что обусловливает увеличение потребления кислорода по сравнению с уровнем покоя.
Кислородный долг при работе различной интенсивности может вызываться разными факторами. Так, при кратковременной интенсивной работе в образовании кислородного долга решающую роль играет израсходование запасов КрФ и АТФ, а при продолжительной работе — расходование гликогена и смещение кислотно-основного равновесия. После работы, выполненной в устойчивом состоянии, кислородный долг наполовину восполняется уже за 30 с, а полностью — за 3—5 мин. После интенсивной работы "погашение" кислородного долга происходит в две фазы: сначала быстрое, а затем — медленное, затяжное возвращение к дорабочему уровню потребления кислорода (см. рис. 146).
Быстрый компонент О2-долга (алактатный) включает то количество кислорода, которое необходимо для ресинтеза АТФ и КрФ. Он характеризует вклад креатинфосфатного механизма в энергетическое обеспечение работы. Медленный компонент О2-долга (лактатный) включает то количество кислорода, которое необходимо для окисления образовавшейся молочной кислоты при выполнении работы. Его величина может характеризовать участие гликолитического механизма в энергетическом обеспечении мышечной работы, а при длительной работе — и других процессов, долю которых оценить весьма затруднительно. Медленный компонент 02-долга восполняется наполовину за 15—25 мин, а полностью устраняется за 1,5—2 ч.
Установление величин кислородного прихода во время работы, а также кислородного запроса и кислородного долга позволяет оценить относительную мощность выполняемого упражнения. Для этого рассчитывают энергетические затраты в единицах максимального метаболического уровня (ММУ). Эта величина представляет собой отношение уровня кислородного запроса (RO2 = (EVO2 + O2D) / Тупр) к индивидуальному МПК (VO2 max):
ММУ = (EVO2 + O2D) / VO2max⋅ Tyпp.