Основы биохимии - А. А. Анисимов 1986
Биомембраны
Строение мембран
Мембраны — неотъемлемый компонент всех клеток. Они окружают клетку, отделяют один клеточный компартмент от другого, образуют структуры разнообразных клеточных органелл. Сеть внутриклеточных мембранных систем особенно хорошо развита у высокодифференцированных клеток. Мембраны представляют собой сложные структуры толщиной 6—10 нм, состоящие в основном из белков и липидов. Кроме того, в них присутствуют углеводы, неорганические соли, вода и ряд других соединений; в некоторых мембранах, в частности, обнаружены следы РНК (до 0,1%).
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 г. С. Синджером и Дж. Николсоном. Согласно данной модели мембрана — это подвижная мозаика, образованная вязкой липидной фазой и погруженными в нее белками (рис. 9.1). Часть белков глубоко расположена в липидном слое или даже пронизывает его насквозь — интегральные мембранные белки, другие лишь частично соприкасаются с липидами, они рыхло соединены с мембраной — периферические белки.
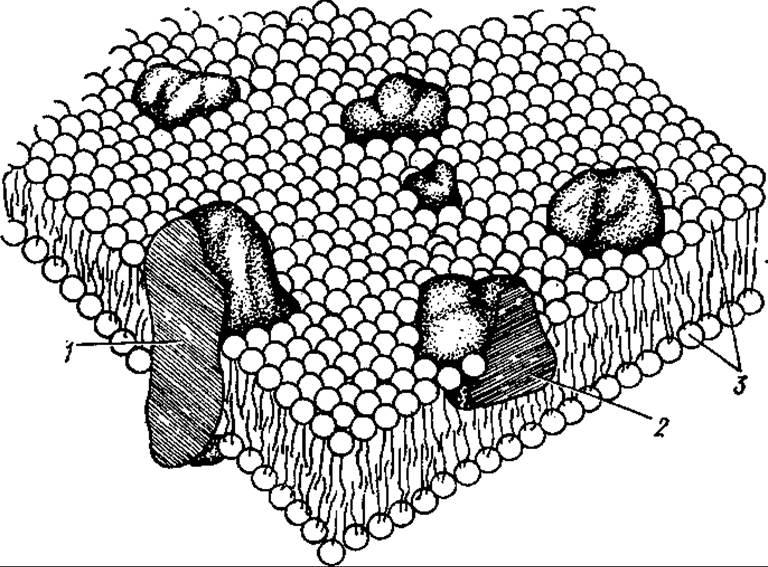
Рис. 9.1. Мозаичная модель структуры мембраны:
1 — интегральный белок, 2 — периферический белок, 3 — липидный бислой
Среди мембранных белков лишь очень небольшая часть (в основном периферические белки) водорастворима и легко извлекается из мембран. Интегральные белки отделяются от липидов только при применении детергентов, органических растворителей. На долю белков может приходиться от 20 до 80% от общей массы белков и липидов. Их содержание зависит от происхождения, строения и функции конкретных мембран. Главную часть липидной фракции мембран составляют фосфолипиды (до 80—90% от общего количества липидов). Помимо них в мембранах часто встречаются и такие нейтральные липиды, как гликолипиды и холестерин. В некоторых случаях до 16% липидов могут составлять моно-, ди- и триацилглицерины.
В липидах мембран многих прокариот обнаружены специфические для них жирные кислоты: с разветвленной цепью и циклопропановой. Липидный состав мембран, особенно степень насыщенности жирнокислотных остатков, изменяется в процессе адаптации организмов к новым температурным условиям среды. В этом заключается одна из функций липидов. Липидный слой (матрикс) мембран двойной, углеводородные длинные цепи (хвосты) жирных кислот обращены друг к другу и образуют гидрофобную толщу мембраны, а полярные группы (фосфатные и спиртовые — холин, этаноламин и т. д.) направлены кнаружи. Интегральные мембранные белки тоже содержат гидрофобные и гидрофильные части молекул, первые вступают во взаимодействие с гидрофобными частями липидных молекул, вторые — располагаются в поверхностном слое мембран, контактируют с полярными «головками» липидов.
Новейшие данные, полученные методом рентгеноструктурного анализа, показали, что цепи мембранных белков сворачиваются, по-видимому, так, что а-спиральные и ß-структурные участки оказываются погруженными в гидрофобную область мембраны; находящиеся вне мембраны части молекулы образованы преимущественно неупорядоченными структурами.
У мембран различают наружную и внутреннюю стороны, которые в большинстве случаев имеют неодинаковый состав, т. е. мембраны асимметричны. Липиды и белки, расположенные на наружной стороне плазматической мембраны, обычно имеют ковалентно связанные с ними углеводы. Внутренняя сторона мембраны и внутриклеточные мембраны, как правило, лишены углеводов. Углеводная часть представлена полисахаридами, включающими обычно не более 15 моносахаридных остатков, которые часто образуют разветвленные структуры. В плазмалемме эукариотических клеток часто обнаруживаются галактоза, манноза, фукоза, N-ацетилглюкозамин, N-ацетилгалактозамин, арабиноза, ксилоза, нейраминовая кислота. Гликолипиды представлены гликозилдиацилглицеринами (преимущественно в бактериальных мембранах) и гликосфинголипидами: цереброзиды, ганглиозиды и др. (в основном у эукариотических клеток).
Мембрана представляет собой динамичную структуру. Наиболее подвижным компонентом в ней являются липиды. Они довольно свободно двигаются в плоскости липидного слоя (латеральное перемещение), меняя своих «соседей» в среднем 106 раз/с. Молекулы белков также могут перемещаться латерально в плоскости мембраны. Возможно также, что белковые молекулы вращаются вокруг перпендикулярных и параллельных плоскости бислоя осей, что может иметь большое значение при функционировании макромолекул и мембран в целом.
Однако белки распределены в мембране не статистически, образуя участки с различными функциями. Иначе говоря, белковые молекулы не абсолютно свободно перемещаются в плоскости мембраны, поскольку могут существовать взаимодействия между отдельными белковыми молекулами и, кроме того, между белками мембран и цитоскелетом клетки: структурными белками, микрофиламентами, микротрубочками, примыкающими к мембране изнутри. В свою очередь расположение белковых молекул в мембране оказывает влияние на распределение и ориентацию липидных молекул в зависимости от сродства конкретных белков и липидов.
Подвижность мембранных молекул в значительной мере зависит от состава жирных кислот. Более упорядоченной и стабильной является структура мембран, содержащая большое число насыщенных жирных кислот в фосфолипидах, менее упорядоченной — содержащая значительные количества ненасыщенных жирных кислот. При оптимальных для жизнедеятельности живых организмов температурах мембрана, как правило, имеет жидкокристаллическое состояние (промежуточное между жидким и твердым). Это состояние обусловлено прежде всего наличием в мембранах системы липид — белок — вода, формирующей различного типа упорядоченные структуры, обладающие в то же время определенной подвижностью. Такое состояние мембран оказывает существенное влияние на их функционирование и объясняет большую чувствительность к различным внешним факторам.
Соседние клетки одной ткани должны сообщаться друг с другом для того, чтобы координировать свою жизнедеятельность и функционировать как целое в соответствии со спецификой ткани. Такое сообщение достигается с помощью специальных коротких ятрубочекя, которые собраны в дискообразные структуры в местах так называемых щелевых контактов. Каждая трубочка состоит из двух цилиндрических белковых молекул — коннексонов. Молекула коннексона частично погружена в клеточную мембрану, а ее выступающая часть способна связываться в межклеточном пространстве с коннексоном соседней клетки, так что образуется непрерывный канал, соединяющий внутреннее пространство двух клеток.
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов: плазматическая мембрана (наружная клеточная мембрана, плазмалемма), ядерная мембрана, эндоплазматический ретикулум, мембраны аппарата Гольджи, митохондрий, хлоропластов, миелиновых оболочек, возбудимые мембраны. У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются инвагинацией (впячиванием) внутрь наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Хороший объект для изучения строения и свойств плазмалеммы представляют собой мембраны эритроцитов, их сравнительно легко получить в чистом виде, поскольку эритроциты не содержат внутриклеточных мембран. Эритроцитарная мембрана состоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная — с липидами. Мембрана асимметрична, так как углеводные компоненты расположены исключительно на наружной стороне. Асимметрична мембрана и в отношении ферментативной активности, липидного состава. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружной — фосфатидилхолин.
Хорошо исследованным интегральным белком эритроцитарных мембран является гликофорин-амфипатический белок. Его N- и С-концы гидрофильны и располагаются соответственно на наружной и внутренней стороне мембраны. Средняя часть гидрофобна, она пронизывает мембрану (рис. 9.2). К N-концевой части присоединено 20—30 молекул полисахаридов, состоящих преимущественно из N-ацетилгалактозамина, галактозы и сиаловой кислоты, расположенной на конце полисахарида. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов.
В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин — белок, выполняющий структурную роль. Он прочно ассоциирован с актиноподобными белками эритроцитарной мембраны, образуя сходную с актомиозином АТФ-зависимую систему.
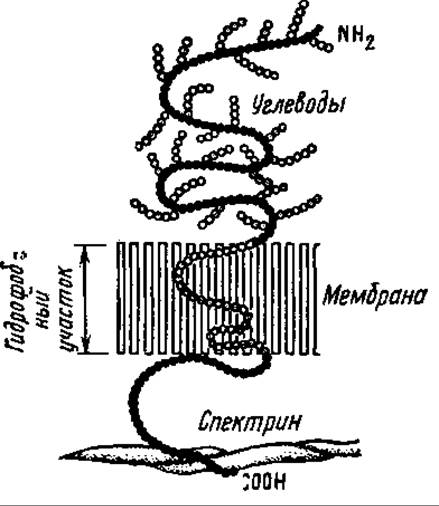
Рис. 9.2. Расположение молекулы гликофорина в эритроцитарной мембране
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них — фосфолипиды). Белки этих мембран важны для фиксации лежащих друг над другом мембранных слоев.
Хлоропласты покрыты двухслойной мембраной. Наружная имеет некоторое сходство с таковой у митохондрий (см. разд. 7.2). Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система — ламеллы. Ламеллы образуют или уплощенные пузырьки — тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы образованы глобулярными липопротеиновыми субъединицами. На наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки белков, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте с гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, окислительного фосфорилирования, активного транспорта; ферменты, участвующие в образовании компонентов мембраны, например гликозилтрансферазы, катализирующие синтез липополисахаридов наружной мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3 : 1. Состав белков мембраны регулируется в зависимости от потребностей клетки и внешней среды. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (того-, микровирусы) в мембраны входит 70—80% всех белков, остальные белки содержатся в нуклеокапсиде.
Белки вирусных мембран — это специфические белки, например гемагглютинин, нейраминидаза. Значительную часть массы вирусной мембраны составляют углеводы. Специфичность олигосахаридов на поверхности вируса определяется в некоторой степени клеткой-хозяином, так как в присоединении углеводов к белкам оболочки участвуют трансферазы сахаров клеток-хозяев. Липиды вирусных мембран также происходят от мембран инфицированных клеток.
Мембраны митохондрий охарактеризованы в разд. 7.2.