БІОНЕОРГАНІЧНА ТА БІООРГАНІЧНА ХІМІЯ - М.В. Яцков - 2014
II. ПРЕДМЕТ, ЗАВДАННЯ, МЕТОДИ ТА ОСНОВНІ НАПРЯМКИ ДОСЛІДЖЕНЬ БІООРГАНІЧНОЇ ХІМІЇ
2.3. Нуклеїнові кислоти
Нуклеїнові кислоти (лат. Nucleus - ядро) - біологічно активні полімери (полінуклеотиди), що складаються з мононуклеотидів, з’єднаних між собою фосфорноестерними зв’язками. Залежно від хімічної природи пентоз, що входять до складу молекули нуклеїнової кислоти, розрізняють дезоксирибонуклеїнову (ДНК) і рибонуклеїнову (РНК) кислоти. Нуклеїнові кислоти містяться у всіх клітинах живих організмів та виконують ряд найважливіших функцій по зберіганню і передачі генетичної інформації, беруть участь в механізмах, за допомогою яких вона реалізується в процесі біосинтезу клітинних білків. У живих клітинах нуклеїнові кислоти знаходяться у вільному стані та в комплексі з білковими сполуками (у складі нуклеопротеїдів).
Структура рівнів організації будови молекул нуклеїнових кислот
Для нуклеїнових кислот характерні первинна, вторинна і третинна структура молекул.
Перинна структура молекул ДНК і РНК створюється внаслідок послідовного з єднання нуклеотидів один з одним за допомогою 3’-5’-фосфодіестерних зв’язків.
Первинна структура ДНК і РНК значною мірою визначає вищі рівні їх організації - вторинну і третинну.
Вторинна і третинна структури ДНК і РНК. ДНК - біополі- мер, молекула якого побудована з великої кількості мононуклеотидів. Міститься переважно в ядрах клітин і, як правило, в комплексі з білками (гістонами і протамінами, зрідка - альбумінами і глобулінами), створюючи хімічну основу хромосом. ДНК є носієм і зберігачем генетичної інформації клітини, а фрагменти молекули, що здійснюють ці життєво важливі функції, називають генами. Схему будови молекули ДНК відображає модель Кріка і Уотсона (рис. 4).
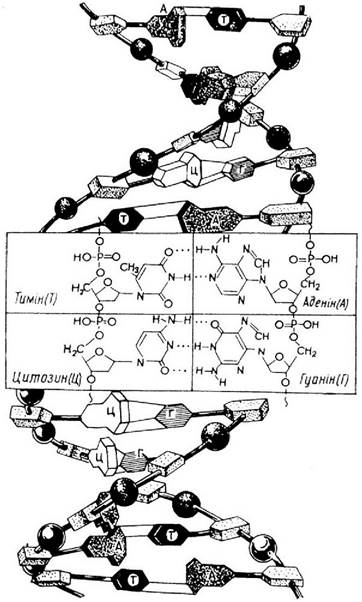
Рис. 4. Схема будови молекули ДНК (за Ф.Кріком і Д.Уотсоном)
ДНК - універсальний зберігай і джерело генетичної інформації, що записана самою Природою у вигляді певної послідовності розміщення основ і визначає властивості живого організму. ДНК здатна при діленні клітин до точного самокопіювання (конваріантна редуплікація). У деяких вірусів такі функції виконує РНК. На молекулі ДНК, як на матриці, синтезуються матричні, або і РНК (мРНК), що є, в свою чергу, матрицями для біосинтезу білка (на рибосомах). На ДНК синтезуються рибосомальні РНК (рРНК), що створюють структурну (частково і функціональну) основу рибосом, білкосинтезуючого апарату клітини. На ДНК синтезуються також транспортні РНК (тРНК), що беруть участь у біосинтезі білка, виконуючи функції акцепторних молекул-переносників амінокислот до місця біосинтезу білків (рибосом).
Отже, існує три типи РНК - рибосомальна (рРНК, або rРНК — 75-85% РНК клітини), транспортна РНК (тРНК, або sPHK - 15%) і інформаційна РНК (ІРНК, або РНК - 1-5%). Всі типи РНК побудовано за однією схемою: їх молекула - продукт поліконденсації нуклеотидів за рахунок утворення естерних зв’язків між C3 одного нуклеотиду і C5 другого за допомогою залишку ортофосфатної кислоти.
Рибосомальна РНК становить хімічну основу органоїдів біосинтезу білка рибосом (50-65% загальної маси). Майже вся РНК рибосом знаходиться у вигляді магнієвої солі. Транспортні РНК іноді називають розчинними РНК (sPHK). їх є кілька десятків (декілька на амінокислоту). Транспортні РНК мають форму листка клена або конюшини.
Інформаційна РНК переносить інформацію від генів (частинок ДНК) до рибосом, з’єднується з ними (а часто і об’єднує їх в полісоми) і кодує біосинтез певних білків.
Фізико-хімічні властивості нуклеїнових кислот
Нуклеїнові кислоти — високомолекулярні речовини, розчиняються у воді, утворюючи колоїдні розчини. Вони мають всі властивості, що характерні для високомолекулярних органічних сполук, зокрема розчинів білків. Гідрофільність макромолекул нуклеїнових кислот значною мірою зумовлена наявністю у них залишків ортофосфатної кислоти. В розчинах нуклеїнові кислоти ведуть себе як поліаніони з різко вираженими кислотними властивостями. Розчинність ДНК гірша, ніж РНК. При нагріванні та дії коагулюючих засобів нуклеїнові кислоти денатурують. Нуклеїнові кислоти - оптично активні речовини.
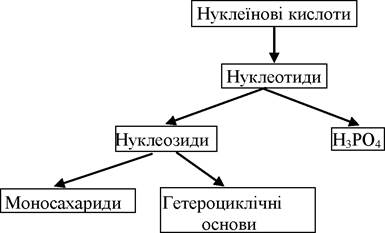
При гідролізі нуклеїнових кислот утворюються структурні одиниці - нуклеотиди, нуклеозиди, фосфатна кислота, моносахариди та гетероциклічні основи:
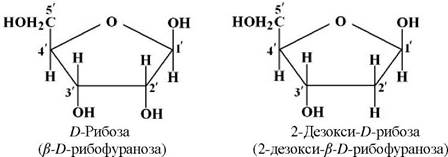
Залежно від виду моносахариду (пентоди) розрізняють два типи нуклеїнових кислот: рибонуклеїнову кислоту (РНК), яка містить D-рибозу та дезоксирибонуклеїнову кислоту (ДНК), яка містить 2-дсзокси-D)-рибозу:
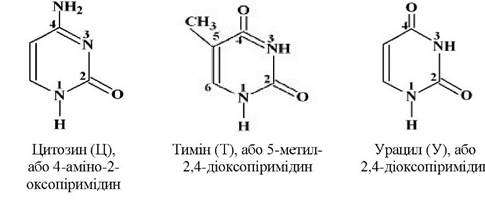
В обох типах нуклеїнових кислот містяться азотисті основи груп пурину - аденін (А), гуанін (Г) і піримідину - цитозин (Ц), тимін (Т) або урацил (У):
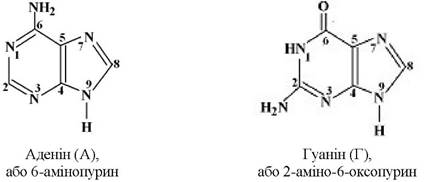
Всі азотисті основи прийнято позначати першою літерою їхньої назви: А, Г, Ц, Т, У. Слід зазначити, що урацил завжди міститься лише в РНК, а тимін - в ДНК. Назву «кислоти» нуклеїнові кислоти дістали завдяки наявності в їхній молекулі залишку фосфатної кислоти.
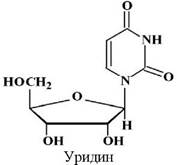
Нуклеозиди та нуклеотиди
Нуклеозиди побудовані з цукру та основи, а нуклеотиди - з цукрів, основи та фосфатної кислоти.
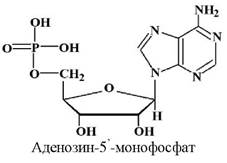
Приклад нуклеотиду:
Встановлено, що в молекулі нуклеозиду гетероциклічні основи сполучаються з рибозою N-глікозидним зв'язком за участю NH-групи дев’ятого за нумерацією атома Нітрогену для пуринових і першого для піримідинових основ (див. табл. 17).
Таблиця 17
Назви нуклеотидів
Гетероциклічна основа |
Монофосфати |
Кислоти |
Позначення |
Урацил |
Уридин-5’- монофосфат |
5’-Уридилова |
УМФ |
Цитозин |
Цитидин-5’- монофосфат |
5’-Цитидилова |
ЦМФ |
Цитозин |
Дезоксицитидин- 5’-монофосфат |
Дезоксицитидилова |
дЦМФ |
Тимін (тільки до ДНК) |
Тимідин-5’- монофосфат |
Тимідилова |
ТМФ |
Аденін |
Аденозин-5’- монофосфат |
5’-Аденілова |
АМФ |
Гуанін |
Гуанозин-5’- монофосфат |
5’-Гуанілова |
ГМФ |
Аденін |
Дезоксиаденозин- 5’-монофосфат |
Дезоксиаденілова |
дАМФ |
Гуанін |
Дезоксигуанозин- 5’-монофосфат |
Дезоксигуанілова |
дГМФ |
Аденін |
Аденозин-5’- монофосфат |
3’-Аденілова |
3-АМФ |
Утворення N-глікозидних для цитозину, тиміну й урацилу вимагає таутомерізації їх до лактамної форми.
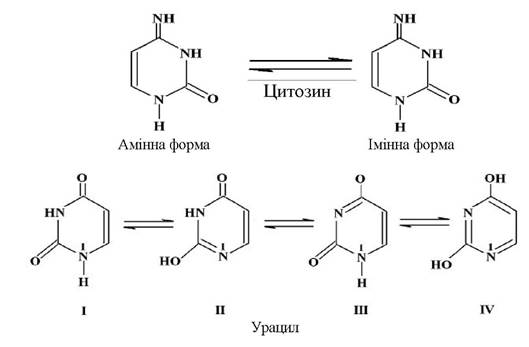
Здатність гетероциклічних основ до таутомерних перетворень відіграє важливу роль при формуванні просторової структури ДНК і РНК. Для пуринових основ можливе існування відповідних таутомерних форм (на прикладі гуаніну):
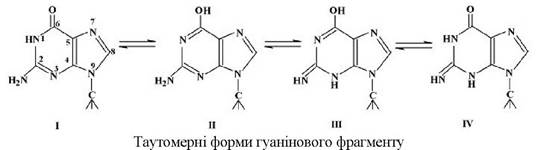
У випадку піримідинових основ спостерігається лактим-лактамна й імін-енамінна таутомерії. Наприклад, для цитозину можлива імінна таутомерія з двох форм, а для урацилу можливе існування чотирьох таутомерних форм:
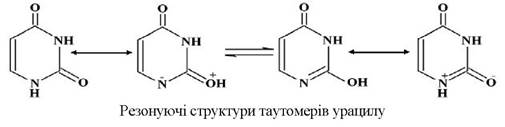
Кожну з таутомерних форм можна показати двома резонуючими структурами (на прикладі урацилу):
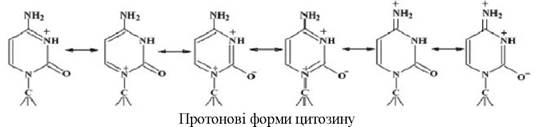
Гетероциклічні основи можуть також утворювати протонові форми (на прикладі цитозину):