ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 2. ЕКОЛОГО-БІОХІМІЧНІ ВЗАЄМОДІЇ ПРОКАРІОТІВ, МІКРОСКОПІЧНИХ ГРИБІВ І ВОДОРОСТЕЙ З ВИЩИМИ РОСЛИНАМИ ТА ТВАРИНАМИ
2.4. Механізми дії мікроорганізмів на рослини
Взаємовідношення між мікроорганізмами та вищими рослинами можуть набувати найрізноманітніших форм. Одна з таких форм — це взаємовигідність для вищої рослини. Типовим прикладом такого є добре вивчені мікоза та ризосфера.
Мікоза (гр. mykes — гриб) — взаємовигідне співжиття (симбіоз) міцелію гриба з корінням вищих рослин, створюють головним чином базидіальні гриби (агарикові та болетові), а також деякі зигоміцети, аскоміцети. Мікориза поширена серед різних груп вищих рослин, насамперед деревних. Вона трапляється також серед багаторічних трав і рідше однорічних.
За екзотрофної мікоризи гриб (міцелій) обліплює корінь і залишається на його поверхні (наприклад, симбіоз базидіальних грибів із лісовими деревами). Ендотрофна мікориза характеризується тим, що гриб проникає в тканини кореня (мікроскопічні гриби з рослинами родин архідних і верескових). Трапляється також перехідна, або ектоендотрофна, мікориза. Мікориза може бути у формі мутаалістичного симбіозу, за якого користь отримують як вищі рослини, так і гриби, або ж у вигляді обмеженого паразитизму.
Фізіологічна роль мікоризи полягає в тому, що мікроскопічні гриби забезпечують вищі рослини мінеральним живленням й органічними сполуками, сприяють регулюванню водного режиму. Гриби виробляють для вищих рослин речовини типу активаторів росту й вітамінів, а самі споживають вуглеводи з коренів рослин.
Гриби, що утворюють мікоризу в дерев, поширюються лише в умовах певного біоцинозу. Тому в лісництві і плодівництві проводять онокуляцію саджанців мікрофлорою. Для цього ґрунт, де раніше росла рослина, додають під час садіння дерев.
До кореня рослин безпосередньо прилягає шар ґрунту (2—3 см), в якому підвищений вміст мікроорганізмів. Цей шар ґрунту має назву ризосфера (гр. rhiza — корінь і sphaire — шар). Склад мікроорганізмів ризосфери залежить від типу ґрунту, виду та віку рослин. Вона збагачена виділеннями коренів, відмерлими кореневими волосками, що забезпечує живлення мікроорганізмів.
Мікроорганізми ризосфери здатні переводити деякі речовини ґрунту в легкодоступні для рослин, синтезують біологічно активні речовини, підсилюють адбсорційну здатність кореневої системи, стимулюють і активують її, беруть участь у денітрифіксації тощо.
Ще один тип взаємовигідності асоціації гриба з рослиною можна розглянути на прикладі пасовищного райграсу (Lolium perenne). Стійкість цієї рослини до шкідливих факторів Listonotus bonariensis зумовлена наявністю в організмі рослини так званого токсину Lolium — ендофіта, який продукується грибом Acremonium coenophia- lum. У рослинах костриці Festuca orientalis, яка в симбіозі живе з ідентичним грибом — ендофітом, знайдено піролізидинові алкалоїди N3-ацетиллоїл і N-форміллоїн.
Зооспори патогенних грибів принаджуються до коренів рослин за участю атрактантів (див. розд. 3.4). Установлено хімічну структуру ряду таких атрактантів. Так, наприклад, для Aphanomyces euteiches це 5,4'-диокси-7-метоксиізофлаон (прунетин) (рис. 2.2).
Спектр біологічної дії прунетину дуже широкий: естрогенна та антиоксидантна активність, стимуляція активності оксидази індолоцтової кислоти тощо.
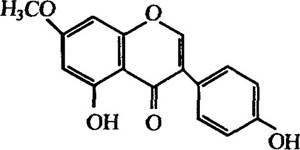
Рис. 2.2. Структура прунетину з коренів гороху Pisum sativum, який є атрактантом до зооспор Aphanomyces
Патофітотоксини. Паразитичні мікроорганізми мають цілий ряд засобів уражаючої дії на вищі рослини. Один із них — це продукування та виділення патофітотоксинів. Нині охарактеризовано значну кількість бактеріальних і грибних патофітотоксинів.
Низькомолекулярні патофітотоксини, як правило, впливають на ріст рослин і слугують причиною хвороби, яка викликає прив’ядання листя і пагонів — виші (англ. Wilt — в’янення), а високомолекулярні — зумовлюють некрози рослин, мацерацію тканин (роз’єднання клітин) рослин тощо. Низькомолекулярні та високомолекулярні патофітотоксини можуть утворюватись тими самими організмами. Наприклад, голландська хвороба в’язів спричинена грибом Ceratocystis ulmi, який переноситься комахами, зокрема короїдом Scolytus multistriatus. Гриб виділяє кілька патофітотоксинів, які викликають некротичне ураження листя та пагонів, а також їх в’янення. Вони становлять суміш глікопротеїнів і низькомолекулярних фенольних метаболітів (рис. 2.3).
Гриби різних видів Fusarium (зокрема Fusarium oxysporum) продукують мікомаразмін і фізаринову кислоту (рис. 2.3), які зумовлюють вілт у томатів. Перший із цих патофітотоксинів має хелатуючі властивості щодо металів, його транслокація та функціонування пов’язані з утворенням водорозчинного комплексу із ферумом. Утворення фузаринової кислоти залежить від наявності в середовищі цинку — за його нестачі синтез цієї кислоти пригнічується.
Фузаринову кислоту, крім томатів, виявлено і в інших рослинах (бавовни, льону і т. ін.), які були інфіковані різними видами Fusarium.
Піколінова кислота (рис. 2.3), як і фузаринова, є хелатором металів, зв’язує іони феруму та купруму, що призводить до розвитку, наприклад у рису, пірикулярозу. Ця кислота детоксикується стійкими сортами рису з утворенням складних метилових етерів.
Крім токсичних фенолів, які викликають голландську хворобу в’язів, похідних перидину, що призводять до вілту томатів і пірикуляриозу рису, існує ще цілий ряд патофітотоксинів, які спричиняють хвороби рослин. Серед них циклічні пептиди (наприклад, тентоксин
Altemaria tenuis), деякі нафтохінони, більшість терпеноїдів (наприклад, гельмінтоспорозид з Helminthosporium sativum) (рис. 2.3).
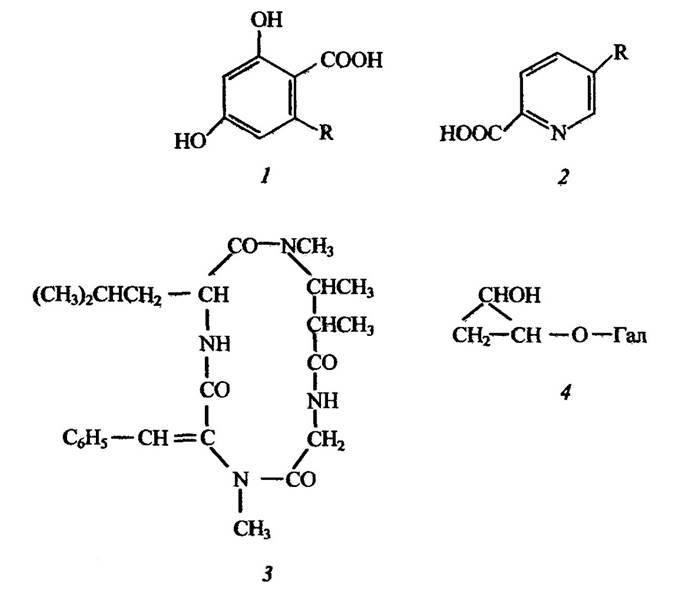
Рис. 2.3. Структура низькомолекулярних патофітотоксинів, які продукуються грибами:
1 — пагофітотоксини Ceratocystis ulmi R=CH2COCH3; CHOHCOCH3 або СОСОСНз (голландська хвороба в’язів); 2 — піколинова кислота, R = Н (Piricutaria oryzae, пірикуляриоз рису), фузаринова кислота, R = С4Н9 — бутил (Fusarium oxysporum, вілт томатів); 3 — тентоксин (Altemaria tenuis, хлороз бавовни); 4 — гельмінтоспарозид (Helminthosporium sativum, плямистість цукрової тростини): = Гал—галактоза
Мікроорганізми здатні вражати рослини навіть без вироблення патофітотоксинів. Іноді вони, як уже відомо, діють на метаболізм рослинної клітини, наслідком якого є, наприклад, нагромадження великої кількості якогось первинного метаболіту. Так, гриб Rhizopus (родина Mucoraceae) може викликати накопичення в мезокарпі рослин фумарової кислоти. Потім вона транспортується в листя та пагони, де охиснюється з утворенням токсичної епоксибурштинової кислоти. Наступний етап — перетворення цього токсину в тартарову кислоту, а її —- у щавлевооцтову, яка вступає в цикл Кребса (цикл трикарбонових кислот) (рис. 2.4).
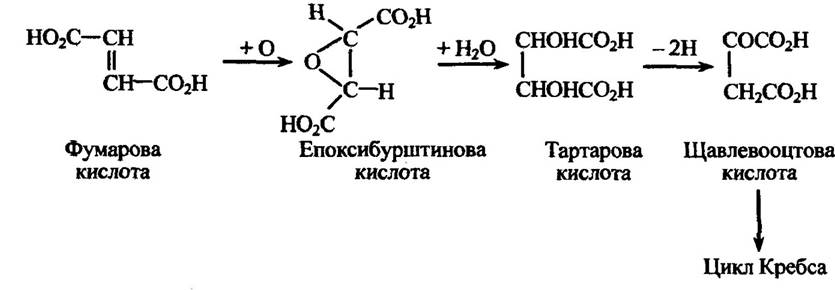
Рис. 2.4. Метаболізм фумарової кислоти, у процесі якого утворюється токсична епоксибурштинова кислота
Отже, у цьому разі за надлишку метаболіту (фумарової кислоти) утворюється патофітотоксин — епоксибурштинова кислота, яка здатна ушкоджувати клітини рослин.
Високомолекулярні патофітотоксини, як правило, високотоксичні за низьких концентрацій. Одна з таких речовин — вікторин, який продукується Helminthosporium victoriae та вражає рослини вівса. Інші подібні високомолекулярні патофітотоксини було виділено з тканин гороху, кукурудзи, сарго.
До симптомів, які можуть бути безпосередньо пов’язані з дією патофітотоксинів, належать також хлороз і некроз. Хлороз (деструкція хлоропластів) призводить до втрати зеленого кольору листя. Такий самий ефект викликає нагромадження в листі аміаку. Так, наприклад, токсин збудника рябухи тютюну бактерії Pseudomonas tabaci становить собою невеликий пептид, який інгібує глутаматсинтетазу. Як наслідок цього — аміак, який утворився за відновлення нітрату, не взаємодіє з глутаміновою кислотою та накопичується.
Порушення росту рослин у більшості випадків зумовлено збільшенням синтезу гормонів росту, які змінюють метаболізм і стимулюють швидкий ріст трав’янистих рослин у довжину. Стебла витягаються і під власного вагою згинаються, що призводить до полягання рослин. Вони слабнуть і стають досяжними для сапрофітних грибів. Прикладом продуцента-патогену для рису є гриб Gibberella fijikuroi. Гібереліни, які отримали назву від цього гриба, утворюються також іншими видами грибів, зокрема з роду Taphrina.
Синтез гормонів росту було виявлено при зараженні гороху Coiynebacterium fascians — утворення цитокініну призводить до появи фасціації. Індолилоцтова кислота (ауоксин), яка продукується Agrobacterium tumefaciens, викликає у рослин пухлини карончатого гала (кореневого раку).
Загальним симптомом захворювання є некроз — поява в тканинах листків ділянок, які мають темне забарвлення, сухих і крихких. Одна з можливих причин некрозу — інгібування первинного метаболізму. Так, наприклад, у разі зараження яблунь бактерією Erwinia amylovora виділяється аміак, який зумовлює некроз пагонів.
Серед ферментів грибів, які руйнують клітинні оболонки в рослин, можна виділити: 1) пептинази (пропектинази, пектинестерази, полігалактуронази тощо), які порушують зв’язки мікрофібрил целюлози з клітинною стінкою; 2) ферменти, що руйнують фібрили целюлози; 3) ферменти, які гідроксилюють інші полісахариди (ксилан, галакгоманан, арабіногалактан і т. ін.); 4) ферменти, які руйнують лігнін.
Ряд грибів продукують фосфатази, дегідрогенази, пероксидази та інші ферменти, що модифікують і руйнують речовини, за допомогою яких рослини захищаються від грибів.
Ферменти грибів виконують біосферну роль редуцентів, які руйнують органічні речовини і в такий спосіб забезпечують розпад в екосистемах полімерів.
Крім продукування ферментів мікроорганізми утворюють також інгібітори ферментів, які регулюють активність цих ферментів, захищають клітини грибів від їхньої дії, а також від дії інших грибів і вищих рослин.
Одним зі способів ураження рослин, що викликає вілт, є закоркування судин, які проводять воду та розчинні в ній речовини. Це відбувається внаслідок того, що деякі гриби виробляють полісахаридні смоли, які закорковують ксилему і, отже, обмежують використання рослинами води. Так, наприклад, гриб Fusarium oxyspomm продукує поряд з патофітотоксинами вазинфуксарин, який закорковує провідні судини рослин. Бактерії також здатні своїми слизовими масами закорковувати судини рослин.
Іноді патофітотоксини можуть прямо діяти на водний обмін у рослинах. Так, наприклад, вони здатні впливати на гормональний контроль функціонування продихів листків.