ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 2. ЕКОЛОГО-БІОХІМІЧНІ ВЗАЄМОДІЇ ПРОКАРІОТІВ, МІКРОСКОПІЧНИХ ГРИБІВ І ВОДОРОСТЕЙ З ВИЩИМИ РОСЛИНАМИ ТА ТВАРИНАМИ
2.5. Засоби стійкості рослин до дії мікроорганізмів
Ряд речовин, які продукуються та виділяються рослинами, здатні впливати на життєдіяльність організмів.
До так званих вторинних сполук рослин, на відміну від білків, вуглеводів, ліпідів і гормонів, належать терпени (терпеноїди), алкалоїди, фенольні сполуки. Хімічна структура та функції деяких із цих сполук викладено як у цьому розділі, так і в розділах 3 і 4. Тут ми наведемо тільки загальні відомості про них.
Терпени. Ці сполуки є похідними так званої ізопренової «C5-одиниці» (рис. 2.5).
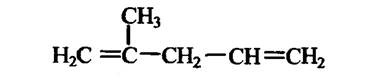
Рис. 2.5. «C5-одиниця» терпенів
Класифікують терпени за кількістю «Cs-одиниць» у молекулі: монотерпени (10 атомів С), сексвітерпени (15 атомів С), дитерпени (20 атомів С) і т. д. до політерпенів (від 7,5 • 103до 3 • 105атомів С).
До терпенів належать етерні олії (зокрема, циклічний монотерпен ментол олії м’яти), фітол, каніфоль, вітамін А і хімічноспоріднені вітаміни E і К (дитерпени), каротиноїди (тритерпени) та багато інших речовин, яких відомо понад 10 000.
Багато терпенів (цитокінін, гіберелін, обсцизна кислота тощо) проявляють гормональні властивості.
Алкалоїди. Ця група рослинних речовин містить нітроген найчастіше у складі гетероциклічного кільця. Як правило, алкалоїди існують у вигляді солей з органічними кислотами (бурштиновою, цитриновою, щавлевою, фумаратною та ін.).
Алкалоїди, які не містять гетероциклічних кілець, називають проалкалоїдами. Як правило, попередниками алкалоїдів є амінокислоти, а в деяких випадках — ацетат. Ті з них, які утворюються без участі амінокислот, називаються псевдоалкалоїдами. їх попередниками є здебільшого терпени.
На даний момент відомо понад 2000 видів алкалоїдів, хімічна будова й біологічна функція приблизно 90% з яких ще не досліджена.
Рослинні феноли. Феноли — це похідні ароматичних сполук, у яких гідроксильна група безпосередньо зв’язана з ароматичним кільцем. Найпростіша з цих сполук — фенол (оксибензол) — С5Н5ОН. Більшість фенолів — безбарвні речовини, іноді рідини з різким запахом. У рослинах в основному поширені фенольні кислоти (прото- катехова, ванілінова та ін.), їхні альдегіди та спирти.
Флавоноїди — це водорозчинні фенольні похідні. Вони мають, як правило, яскраво-червоний, темно-червоний або жовтий колір. Основу їхньої структури становить флавон, який складається з двох ароматичних кілець, з’єднаних С3-залишком. Одна або більше гідроксильних груп у флавоноїдах з’єднується глікозидними зв’язками з вуглеводами.
Таніни — це ароматичні сполуки, які фенольними гідроксильними групами здатні зв’язуватися з білками. Водорозчинні таніни становлять похідні простих фенольних кислот (галової, елагової та ін.), зв’язаних із вуглеводами (глюкозою).
З фенолів флавонового типу утворюються конденсовані таніни — флаволани, які є полімерами флавонів і не містять залишків вуглеводів.
Особливе місце серед сполук, які продукують і виділяють рослини, належить тим, що захищають їх від захворювань, які викликаються мікроорганізмами. Зауважимо, що багато вищих рослин, котрі проростають у природних біогеоценозах, мають велику стійкість до зараження мікроорганізмами або існують симбіози з ними без істотних симптомів захворювань. Водночас культурні рослини значно більше здатні до подібних захворювань. Тому такою актуальною проблемою екологічної біохімії є встановлення речовин рослин, спроможних пригнічувати життєдіяльність мікроорганізмів-паразитів, визначення шляхів їх синтезу, виведення стійких сортів культурних рослин шляхом уведення генетичного матеріалу з диких близькородинних форм, які не сприймають захворювання.
Речовини рослин, які здатні пригнічувати проростання або впливати на ріст мікроорганізмів, поділяють на дві групи — преінфекційні та постінфекційні сполуки.
Перші з них постійно наявні у вищих рослинах незалежно від інфікування. Других у неінфікованих рослинах немає, вони синтезуються тільки після зараження мікроорганізмами.
Преінфекційні сполуки. Серед цих сполук вирізняють дві групи: прогібінти та інгібінти (табл. 2.1).
Таблиця 2.1
КЛАСИФІКАЦІЯ ПРЕІНФЕКЦІЙНИХ ІПОСТІНФЕКЩЙНИХ СПОЛУК
У ВИЩИХ РОСЛИНАХ (за Харборном, 1985)
Клас речовин |
Характеристика класу речовин |
Преінфекційні речовини |
|
Прогібінти |
Метаболіти, які в наявній концентрації зменшують або цілком зупиняють розвиток патогенних мікроорганізмів |
Інгібінти |
Метаболіти, концентрація яких для прояву токсичної дії після іифекціювання рослин різко збільшується |
Постінфекційні речовини |
|
Постінгібінти |
Метаболіти, які утворюються під час гідролізу або окисиення з нетоксичиих попередників |
Фітоалексини |
Метаболіти, які утворюються після інфікування рослин знову після дерепресії генів або активації латентної ферментативної системи |
Більшість прогібінтів — це сполуки фенольної природи, але є також інші. Прикладами прогібінтів можуть слугувати катехол і протокатехова кислота, піносильвін, лутеон, хордатин А і Б, авенацин (рис. 2.6).
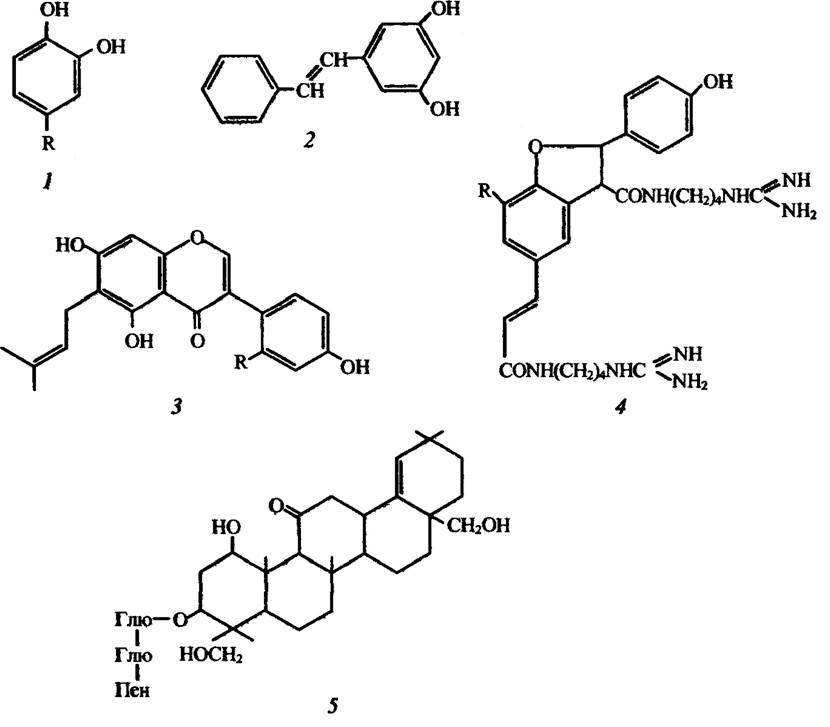
Рис. 2.6. Структура деяких преінфекційніїх речовин, які наявні у вищих рослинах:
І — катєхол (R = Н) і протокатехова кислота (R = СО2Н); 2 — піносильвін; 3 — лутеон (R = ОН) і дезоксилутеон (R = Н); 4 — хордатин A (R = Н) і хордатин В (R = ОСН3); 5 — авенацнн; Глю — глюкоза, Пен — пентоза
Катехол і протокатехова кислота токсичні для спор Colletotrichum circirans. Їхній вміст у стійких до цього гриба сортах цибулі корелює з наявністю в них антицианінів, які потенційно токсичні.
Піносильвін належить до гідроксистильоенів, які надають деревині (зокрема, сосни) стійкість до природного розкладання.
У тканинах люпину білого (Lupinus albus), зокрема в листі, ідентифіковано лутеон (ізопентилізофлавон) і його 2-дезоксипохідну сполуку, які пригнічують ріст міцелію Helminthosporium carbonum.
Хордатини А і Б виконують захисну функцію в проростках ячменю, Hordeum vulgare, який уражається Helminthosporium sativum.
До прогібінтів нефенольної природи належить пентациклічний тритерпеновий глікозид авенацин, який забезпечує стійкість коренів вівса Avena sativa до гриба Ophiobolus graminis.
Розглянуті терпеноїдні та алкалоїдні токсини локалізуються, як правило, у вакуолях листків і коренів. Водночас є прогібінти, які розміщуються на поверхні рослин і, отже, є першими захисниками від мікроорганізмів. Так, наприклад, партенолід (сесквитерпеновий лактон) локалізований у двочасткових залозах на поверхні листків і насіння Pyrethrum parthenium і токсичний для грампозитивних бактерій і нитчастих грибів. Це стосується також скларелу та ізоскларелу — дитерпенів, локалізованих на поверхні листків Nicotiana glutinosa. Нобілетин (тексаметоксифлавон), який розміщується на поверхні листків цитрусових (Citrus sp.), токсичний для гриба Deuterophoma tracheiphila.
Ще одним класом преінфекційних сполук є, як уже зазначалося, інгібінти. Це речовини, концентрація яких збільшується в місцях проникнення інфекції. Так, наприклад, за взаємодії бульби картоплі Majestic зі спорами гриба фітофтори Phytophthora infestans поблизу місця інфікування концентрація кумаріну скополетину збільшується в 10—20 разів, а хлорогенової кислоти — у 2—3 рази.
Подібні збільшення вмісту скополетину, скополину та хлорогенової кислоти характерні також для батату (Ipomea batatas), тютюну (Nicotiana tabacum). Існують також інші інгібінти, які підвищують стійкість рослин до захворювань, що їх викликають мікроорганізми.
Постінфекційні сполуки. Ці сполуки також забезпечують стійкість рослин до дії мікроорганізмів (табл. 2.1). Вони поділяються на постінгібінти та фітоалексини. Постімгібінти наявні в здорових тканинах рослин у вигляді, як правило, неактивних глікозидів. їх перехід в активну форму після проникнення патогенних мікроорганізмів у клітини рослин відбувається в результаті ферментативного гідролізу або окиснення. Прикладом такого процесу може бути гідроліз специфічною β-глкжозидазою ціаногенного глікозиду лінамарину в листках лядвенця рогатого (Lotus corniculatus) за інфікування патогенним грибом Stemphylium loti. Під дією лінамаринази з лінамарину утворюється ціаногідрин, який спонтанно розпадається з утворенням HCN (синильної кислоти), яка захищає рослини від патогенного гриба (рис. 2.7). Проте в цьому разі патоген може адаптуватися до ціаніду. Він здатний синтезувати фермент формамідгідролазу, який перетворює HCN у формамід HCONH2.
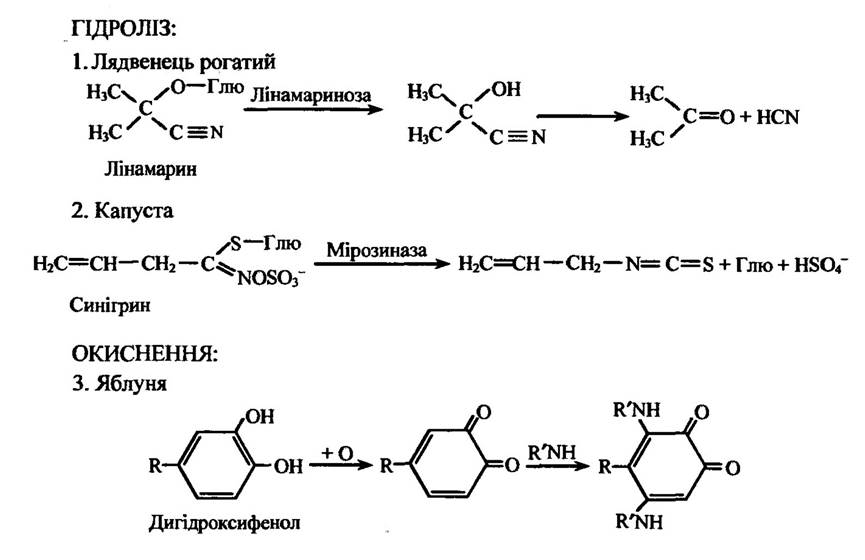
Рис 2.7. Структура та механізми утворення постінгібінтів:
1 — під час гідролізу лінамарину в лядвенця рогатого; 2 — під час гідролізу синігрину в капусті; 3 — при окисненні дигІдроксифенолу (R = СН2СН2СОС6Н4(ОН)2)
Відомі також інші ціаногенні глікозиди. Так, у диких і культурних видів Brassica (капуста, ріпа, турнепс та ін.) при зараженні несправжньою борошнистою росою Peronospora parasitica відбувається виділення леткої олії алилизотіоціанату, яка утворюється під час гідролізу глюкозиду синігрину під дією мірозинази.
Феноли, крім виконання ролі хемоефекторів і хемомедіаторів, також можуть бути постінгібінтами. До таких речовин належать дигідроксифенол, окиснення якого фенофазою призводить др утворення високотоксичного о-хінону, який може конденсуватися з аміносполуками та утворювати ще токсичніші речовини (рис. 2.7).
Постінфекційні сполуки. До них належить велика група фіто- алексинів (гр. aleksos — відбивати напад і phyton — рослина), які утворюються після зараження рослини мікроорганізмами шляхом активації генів або латентної ферментної системи (див. табл. 2.1).
За своєю хімічною структурою ці речовини надзвичайно різні (рис. 2.8). Загальна їхня властивість — розчинність у ліпідах.
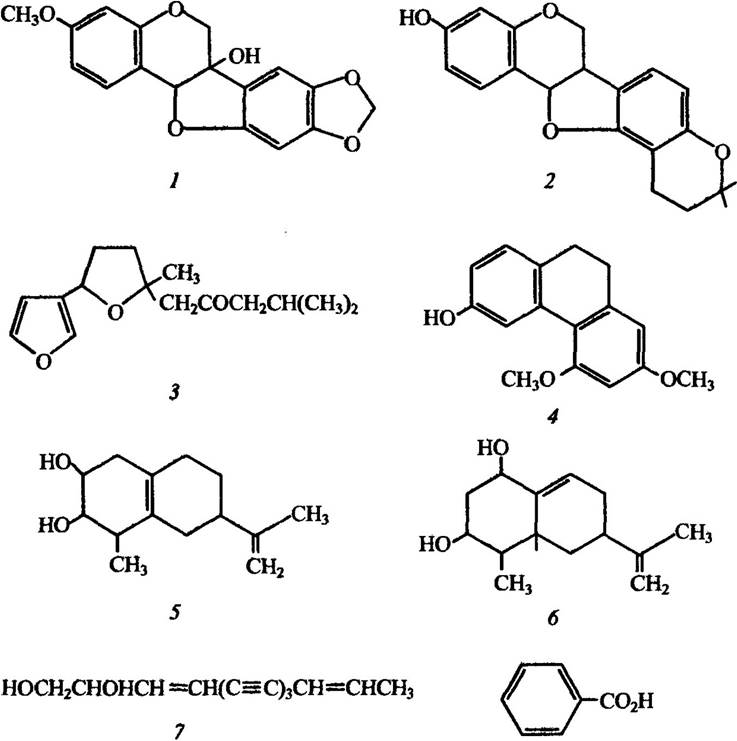
Рис. 2.8. Структура деяких фітоалексинів вищих рослин:
1 — пізатин; 2 — фазеолін; 3 — іпомеамарон; 4 — орцинол; 5 — ришитин;
6—капсидол; 7—сафітол; 8 — бензойна кислота
Типовим прикладом фітоалексинів є пізатин — птерокарпанова похідна, яка утворюється у стручках Pisum sativum, інфікованих грибом коричневої гнилі Monilinia fructigena (рис. 2.8). Інші бобові, зокрема Phaseolus vulgaris, продукують подібну речовину — фазеолін. Утворення фітоалексинів характерне також для Ipomoea batatas і представників родини Convolvulaceae — іпомеамарону, Orchidaceae — орцинолу, Solanaceae: Solanum tuberosum і ришитину, Capsicum frutescens — капсидіолу, Carthamnus tinctorius — сафінолу, Malus pumila i Rosaceae — бензойної кислоти.
Фітоалексини утворюються в разі зараження рослин бактеріями, вірусами та грибами. Крім цього, фітоалексини здатні синтезуватися
під впливом різних абіотичних факторів — дії ультрафіолетового випромінювання, різкої зміни температури, при пораненні тощо. Ці речовини є часткою загальної захисної та відновлювапьної системи в рослинах. Синтез фітоалексинів у клітинах рослин активується під дією хімічних речовин, які називають еліситорами. Ними є широке коло речовин: вуглеводи (глюкоманани, β-1,3-глюкан, гепа-β-глюкозид та ін.), глікопротеїни, ліпіди, ліпоглікопротеїнові комплекси та деякі жирні кислоти (ейкозапентанова та арахідонова) тощо. Властивості еліситорів мають також солі важких металів, ряд детергентів і пестицидів, ультрафіолетове випромінювання тощо.
Місцем синтезу глюканових еліситорів, мабуть, є клітинні стінки грибів і бактерій. Припускають, що у взаємодії еліситорів з клітинами рослин бере участь певна рецепторна ділянка цих клітин, в якій еліситор здатний ініціювати синтез фітоалексину.
Посилення утворення рослинами фітоалексинів за допомогою природних і штучних речовин, так званих сенсибілізаторів, є новим та перспективним шляхом боротьби із захворюваннями рослин. Такі речовини, як правило, мають тривалий термін дії. Тому оброблення посівного матеріалу сенсибілізаторами може попереджувати їх інфікування патогенними мікроорганізмами.
біохімічній взаємодії рослин і мікроорганізмів беруть участь також так звані супресори-метаболіти мікроорганізмів, здатні пригнічувати утворення фітоалексинів.
Отже, біохімічні засоби захисту рослин, як і біохімічні способи паразитування мікроорганізмів на рослинах, виконують функцію хеморегуляторів, а певною мірою — також ценозорегуляторів.