ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 2. ЕКОЛОГО-БІОХІМІЧНІ ВЗАЄМОДІЇ ПРОКАРІОТІВ, МІКРОСКОПІЧНИХ ГРИБІВ І ВОДОРОСТЕЙ З ВИЩИМИ РОСЛИНАМИ ТА ТВАРИНАМИ
2.6. Токсична дія мікроорганізмів на організм тварин
Бактеріальні токсини. Серед токсинів патогенних бактерій білкові екзотоксини найнебезпечніші та здатні викликати у ссавців, у тому числі людей, серйозні захворювання — холеру, дифтерію, бутулізм, правець тощо. Вони дуже різноманітні за структурою та механізмом дії. Більшість із них синтезується бактеріями у вигляді неактивних попередників — протоксинів, які активуються для прояву своєї дії. Під дією протеїнкіназ на протоксин утворюються фрагменти, які формують біфункціональну систему. Так, наприклад, молекули холерного, дифтерійного, бутулінічного, правцевого та деяких інших білкових токсинів містять зв’язані між собою фрагмент А, який має ферментативну активність, і фрагмент
В, що відповідає за зв’язування токсину з рецепторами на поверхні клітини та бере участь у транслокації фрагмента А всередину клітини-«мішені».
Зауважимо, що в патогенних бактерій протеоліз протоксинів обмежений. Напевно, їх активація відбувається за участю як бактеріальних протеолітичних ферментів, так і клітинних протеїназ.
На відміну від простих токсинів, які спочатку синтезуються у вигляді одного поліпептидного ланцюга протоксину, що перетворюється в біфункціональну систему після протеолізу, у складних токсинів активний і рецепторний фрагменти синтезуються незалежно, а відтак з’єднуються в комплекс.
Свою біологічну дію токсини бактерій проявляють у кілька етапів: зв’язування з рецепторами, активація протоксинів, ендоцитоз і транспорт токсину, транслокація каталітичного фрагмента токсину в цитозоль і дія на внутрішньоклітинну «мішень».
Токсини, як правило, зв’язуються з рецепторами на поверхні клітини-«мішені». Так, наприклад, для дифтерійного токсину рецептором є попередник фактора росту, який зв’язує гепарин, а для холерного токсину — один із гангліозидів.
Рецептори можуть брати участь не тільки в забезпеченні зв’язування токсину з поверхнею клітини, а й у транспорті токсину до внутрішньоклітинної «мішені» на різних етапах цього процесу.
Після зв’язування токсинів з рецепторами на поверхні клітин відбуваються в більшості випадків ендоцитоз і транспорт усередину клітини для подальшої транслокації через мембрану в цитозоль до внутрішньоклітинної «мішені». Ще один із можливих механізмів — це зворотний транспорт секреторним шляхом клітини через ендо- соми, апарат Гольджі до ендоплазматичного ретикулуму з подальшою трансолокацією крізь його мембрану. Незалежно від того, яким механізмом ендоцитозу здійснюється транспорт бактеріальних токсинів, вони можуть минати стадію ендосом і досягати місця транслокації інакше. Так, наприклад, для транспорту каталітичного ферменту всередину клітин у мембрані формується канал (пора). Через такі канали може відбуватися також витікання цитозолю.
За механізмом дії бактеріальні токсини поділяють на ті, які пошкоджують мембрани (фосфоліпази, гемолізини, гіалуронідази, колагенази та ін.), а також ті, які діють на внутрішньоклітинні «мішені», усередині клітин. Прикладом останніх може бути стрептокіназа Streptococcus pyogenes, яка гідролізує плазміноген, клостридіальні колагенази та ін.
Внутрішньоклітинними «мішенями» бактеріальних токсинів можуть бути G-білки, актин, рибосоми тощо. Вони здатні каталізувати
певні біохімічні реакції, результатом яких є порушення внутрішньоклітинних «мішеней» (табл. 2.2).
Таблиця 2.2
ДІЯ ДЕЯКИХ БАКТЕРІАЛЬНИХ ТОКСИНІВ (за Позур та ін., 2003)
Токсини |
Ферменти |
Внутрішньоклітинна «мішень» |
Дифтерійний токсин |
АДФ-ризилтрансфераза |
Фактор елонгації EF-2 |
Екзотоксин А Pseudomonas |
АДФ-ризилтрансфераза |
Фактор елонгації EF-2 |
Набряковий антракстоксин |
Аденілатциклаза |
ц-АМФ-модулюючі білки |
Летальний антракстоксин |
Цинкова ендопротеїназа |
Кіназа мітоген-активованої протеїнкінази |
С2 токсин С. botulinum |
АДФ-ризилтрансфераза |
G-актин |
Холерний токсин |
АДФ-ризилтрансфераза |
Гетеротримерний G-білок |
Коклюшовий токсин |
АДФ-ризилтрансфераза |
Гетеротримерний G-білок |
Нейротоксин клостридії |
Цинкова ендопротеїназа |
SNAP-25, сітаксин 1 |
Взагалі існує багато механізмів, за допомогою яких токсини бактерій пошкоджують клітини-«мішені».
Гриби. Ці організми здатні до утворення великої кількості токсинів —мітотоксинів. Деякі з них наведено в табл. 2.3.
Мікотоксини грибів, які утворюють великі плодові тіла, слугують, мабуть, захистом від тварин, що можуть живитися ними до періоду розсіювання спор.
Токсичні гриби, які не утворюють великих плодових тіл (аспаргіла, пеніциліум, фузаріум та ін.), певною мірою, мабуть, захищають не тільки себе, а й кормову базу, на якій вони живляться, від інших консументів. Отже, вони є екологічними хеморегуляторами. Це має істотне значення у взаємодії хижих грибів і нематод. Останні здатні продукувати та виділяти речовини (неміни), які індукують утворення пасток у хижих грибів. Водночас, самі хижі гриби виділяють атрактанти, які приваблюють нематод до пасток. До таких речовин належать, наприклад, леткі терпени, що продукуються Arthrobotrys oligospora та Candelabriella musiformis. У деяких видів грибів ті самі речовини є атрактантами та токсинами для нематод, у інших — ці функції виконують різні речовини.
Таблиця 2.3
ПРИКЛАДИ МІТОТОКСИНІВ (за Барб’є, 1978 з допов.)
Гриб-продуцент |
Токсини |
Спори жита (Claviceps puipureal) |
Алкалоїди, похідні лізергинової кислоти |
Aspergillus spp. |
Афлатоксин |
Aspergillus giganteus |
Білок α-сарпцин |
Aspergillus restricus |
Білки рестроктоцин і мітогілін |
Penicillium islandicum |
Флавоскирин, лютеоскирин, ругулозин, емодин, скирин |
Fusarium solani |
Нематоксини (М1 М2, М3, М4, М5) |
Fusarium gramilearum (Gibberella) |
Зеараленон |
Бліда поганка (Amanita phalloides та ін.) |
Фалотоксини (наприклад, фалоїдин), аматоксини (наприклад, α-аманітин), аманіти-біциклічні октапептиди |
Мухомори (Amanita spp.) |
Мускарин, похідні ізоксазолу |
Гриб теонакатль (Psilocybe) |
Похідні триптаміну |
Біохімічні механізми дії мітотоксинів надзвичайно різноманітні. Це інгібування різних ферментів (у тому числі РНК-полімераз), синтез білків, порушення енергетичних процесів тощо.
Серед речовин, які продукуються та виділяються грибами, є такі, що мають протипухлинну активність. Серед них — полісахариди (приміром, β-1,6-глюкан з Giyfola frondosa), деякі алкалоїди (наприклад, агроклавін, що продукується Claviceps, рядом видів родини Penicillum і Aspergillus та ін).
Разом з тим деякі мікотоксини викликають онкологічні хвороби. До таких речовин належать афлотоксини (рис. 2.9).
Деякі гриби-симбіонти, а також бактерії та дріжджі, які мешкають на шкіряних покривах і шкіряних залозах ссавців, беруть участь у формуванні пахучості цих тварин, що важливо для їх комунікації.
Водорості та ціанобактерії. Одноклітинні водорості та ціанобактерії здатні синтезувати високотоксичні для тварин речовини. Деякі з них наведено на рис. 2.10.
За своєю дією на тварин токсини водоростей поділяють на паралітичні, діаретичні, нейротоксичні та ін.
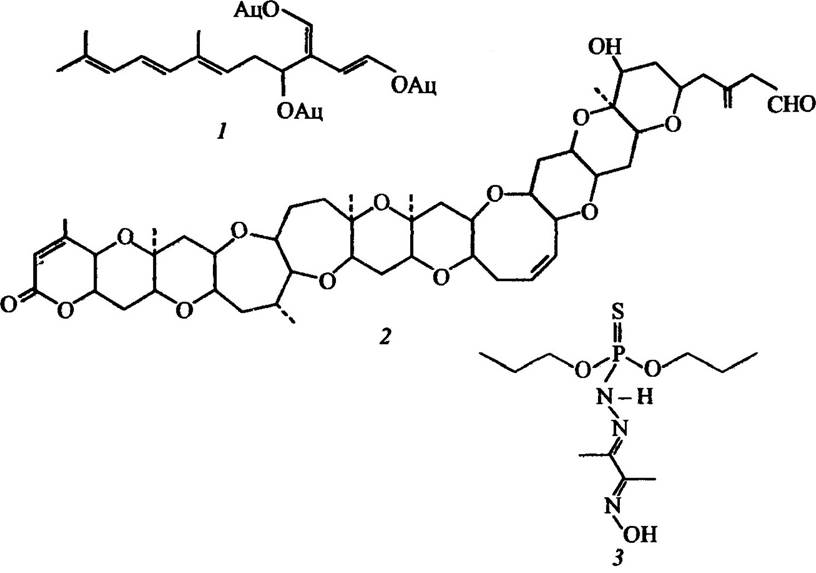
Рис. 2.9. Структура деяких афлотоксинів:
1 — афлотокснн В2; 2 — стеригматин; 3 — охратоксини A (R=H, R1=CI), В (R=H, R1=H), С (R=C2H5, R1=C1); 4—афлотоксин G1
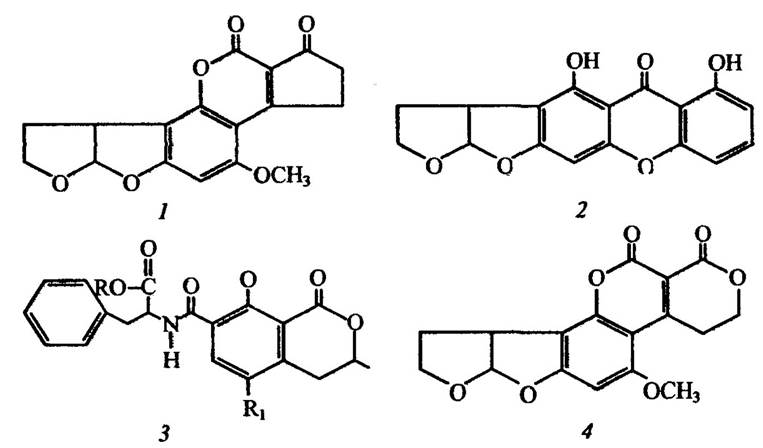
Рис. 2.10. Токсичні для хребетних речовини, які продукуються
та виділяються водоростями:
1 — рипоцефалин із зеленої водорості Rhipocephaius phoenix; 2 — бреветоксин Б водорості Ptychodiscus brevis; 3 — фосфорогідразидотіоат Ptychodiscus brevis; Ац — ацетил
Багаторічні бурі водорості Fucus vesiculosus і Ascophyllum nodosum продукують харчові детеренти, які відлякують молюсків Littorina littorea, що для водоростей є консументами. Ці захисні речовини є поліфітолами — низькомолекулярними флороглюцинольними полімерами.
Між водоростями й безхребетними тваринами можливі антогоністичні взаємодії, які опосередковані екзометаболітами. Так, водорозчинні речовини, які виділяються актинією Condilactis gigantea інгібують ріст зелених, червоних і бурих водоростей.
Водночас ряд водних безхребетних (молюсків, коралів, морських анемонів та ін.) утворюють симбіотичні угруповання з водоростями. Так, актинії Anthopleura elegantissima, певні тканини молюсків Tridacna crocea та інші організми містять симбіотичні водорості, що мають підвищену кількість супероксиддисмутази й каталази. Ці ферменти знешкоджують надлишковий оксиген, який продукується фотосинтезуючими водоростями.
Для життєдіяльності організмів істотне значення має якість і повноцінність середовища мешкання, яке формується під дією низки факторів.
У водних екосистемах гідробіонти виділяють найрізноманітніші органічні речовини в середовище мешкання, що викликає нагромадження їх розчинних форм. До складу таких речовин можуть входити такі, що порушують відношення окисних і відновних процесів у водних екосистемах. Це відбувається внаслідок того, що гідробіонти, зокрема водорості, можуть виділяти у водне середовище розчинні речовини, здатні до окиснення, а також антиоксиданти.
Одним із наслідків виділення у водне середовище екзометаболітів водоростей є підвищення концентрації пероксиду гідрогену. З’ясовано також, що ряд водоростей, зокрема Westella botryoides, виділяють ліпідні екстракти клітин, яким притаманна антибактеріальна дія. Аналогічну властивість мають також метаболіти різних водоростей (табл. 2.4), а також інших гідробіонтів.
Якість і повноцінність водного середовища залежить також від складу та кількості токсинів, що їх виділяють гідробіонти, зокрема водорості.
Мікроорганізми здатні виділяти ряд хелаторів, які можуть зв’язувати метали. З одного боку, це може призвести до дефіциту мікроелементів, а з іншого — знижувати токсичність антропогенного забруднення.
Для гідробіонтів велике значення мають також інші аспекти нагромадження метаболітів у середовищі мешкання, у тому числі ті, які пов’язані з існуванням ферментів, вітамінів і різних біологічно активних речовин.
Таблиця 2.4
ПРИКЛАДИ АНТИБАКТЕРІАЛЬНИХ РЕЧОВИН, ЯКІ ПРОДУКУЮТЬСЯ ТА ВИДІЛЯЮТЬСЯ ВОДОРОСТЯМИ (за Телитченко, Остроумовим, 1990)
Назва речовини |
Продуцент |
Циклічні полісахариди |
Бура водорість Chondryga californicus |
Пахидиктіол А |
Pachydictyon coriaceum |
Терпеноїди удотель; удотеафурон та її похідні |
Зелена водорість Udotea flabellum |
Терпеноїдний триальдегід |
Halimeda |
Неароматичні галогенвмісні антисептики: полігалометани, галооцтові кислоти, галоацетони, галоакрилові кислоти, галактенони |
Червоні водорості родини Bonnemaezoniacae |
Ліпідні екстракти клітин |
Зелена водорість Westella botiyoides |
Мікроорганізми ґрунту, як і води, теж здатні продукувати й виділяти бактерицидні речовини, токсини тощо. їхня життєдіяльність залежить також від якості та повноцінності середовища мешкання. Найрізноманітніші токсини можуть виділяти ціанобактерії (табл. 2.5).
Таблиця 2.5
ДЕЯКІ ТОКСИНИ ЦІАНОБАКТЕРІЙ
(за Телитченко, Остроумовим, 1990)
Назва ціанобактерії |
Токсини |
Структура |
Прісноводні |
||
Microcystis aeruginosa |
Мікроцистин |
Пептид |
Anabaena flosaquae |
Анатоксин a Анатоксин c |
Алкалоїд Пептид |
Aphanizomenon flosaquae |
Анатоксин, сакситоксин, неосакситоксин |
Алкалоїд |
Scy tonema hofmanni |
Ціанобактерій |
С23Н23О6Сl альдегід |
Морські |
||
Lyngbia majuscula |
Лінгбіятоксин А Дебромоаплізнатоксин |
Індольний алкалоїд Фенольний біслактон |
Oscillatoria nigroviridis |
Оксицилатоксин А |
Фенольний біслактон |
Schizothrix calcicola |
Дебромоаплізнатоксин |
Фенольний біслактон |
Tolypothrix conglutinata var. chlorata |
Толитоксин |
N-альдегід |
Зауважимо, що біохімічна взаємодія прокаріотів, грибів і нижчих рослин з іншими організмами сприяла, мабуть, адаптивній реакції цих організмів і, отже, зростанню загальної кількості видів у цих групах.