ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 8. СТІЙКІСТЬ ТА АДАПТАЦІЯ РОСЛИН І ТВАРИН
8.3. Стійкість та адаптація тварин до умов існування
8.3.3. Запобігання переохолодженню та перегріванню
Прояв активного життя у більшості тварин можливий тільки у відносно вузькому діапазоні температур — від кількох градусів точки замерзання чистої води (0 °С) до приблизно 50 °С. Температурні умови, придатні для мешкання тварин, існують у водному середовищі та більшій частині суходолу всюди принаймні в певний час року. Водночас гриби й велика кількість безхребетних Арктики і Антарктики здатні жити у воді, температура якої цілий рік становить близько -1,8 °С. Деякі організми мешкають у гарячих водних джерелах за температури 50 °С і навіть вище. Термофільні бактерії здатні жити за вищих температур — близьких до точки кипіння води (100 °С). Певні тварини, які опинилися в температурних умовах, що виходять за діапазон їхнього активного життя, здатні виживати — вони переходять у стан заклякнення. Деякі з них можуть переносити навіть температуру рідкого гелію (-269 °С). Стійкість тварин до високих температур не така велика, але деякі з них за спокою проявляють значну термофільність. Так, наприклад, спори бактерій здатні виживати за температури +120 °С і навіть вищої.
У ссавців і птахів температура тіла майже стала і мало змінюється навіть за значного коливання температури довкілля, на відміну від них у більшості інших тварин температура тіла близька до температури навколишнього середовища. Існують також деякі тварини (серед нижчих безхребетних і хребетних), які здатні певний час підтримувати значну різницю між температурою тіла й довкілля.
Ссавців і птахів зазвичай відносять до теплокровних, а всіх інших — до холоднокровних тварин. Проте холоднокровні тварини не завжди холодні (мають низьку температуру). Так, наприклад, температура тіла тропічних риб, ящірок у пустелі, комах, що перебувають на сонці, може бути вищою, ніж у ссавців. Водночас деякі ссавці та птахи здатні впадати в зимову сплячку або заклякати, і тоді температура їхнього тіла значно знижується.
Наявні терміни пойкілотермні (гр. poikilos — різноманітний і therme — теплота) замість «холоднокровні» і гомойотермні (гр. hemios — однаковий) замість «теплокровні» теж неточні. Так, пойкілотермні, за визначенням, змінюють температуру зі зміною її в довкіллі. Отже, наприклад, риби повинні мати таку саму температуру, як і різні шари води. Однак глибоководні риби мешкають у шарах води, де температура майже стала, і тому температура їхнього тіла не змінюється. Таких риб коректніше називати гомойотермними тваринами, але цей термін використовують тільки стосовно ссавців і птахів, температура тіла яких може коливатися в межах кількох градусів, а в тих, що впадають у зимову сплячку, — знижуватися майже до 0 °С.
Тварин, температура тіла яких підтримується на високому рівні, що забезпечує активне життя, тільки в певний час, а в інші періоди залежить від температури довкілля, називають гетеротермними (гр. heteros — різний).
Для визначення механізмів теплоутворення і підтримки температури тіла використовують терміни «екотермні» та «ендотермні» тварини. До ендотермних належать ті тварини (ссавці та птахи), у яких висока температура підтримується внутрішнім (метаболічним) теплом, а екзотермних — температура тіла яких залежить від зовнішніх джерел тепла й насамперед сонця. Ці терміни, як і попередні, неточні. Так, наприклад, у тунця температура м’язів може бути на 10—15 °С вищою, ніж навколишня вода за рахунок перебігу в них метаболічних процесів. Проте цих риб не можна вважати ендотермними тваринами, як ссавців і птахів. Більшість комах не є цілком екзотермними чи ендотермними тваринами. Так, наприклад, коли комаха гріється на сонці, то температура її тіла визначається як сонячною радіацією, так і інтенсивністю м’язових скорочень. Деякі ссавці та птахи іноді використовують тепло сонячних променів для зниження особистої теплопродукції.
ПРОЯВ ОСНОВНИХ ТЕМПЕРАТУРНИХ ЕФЕКТІВ У ТВАРИН НА БІОХІМІЧНОМУ РІВНІ. Атоми й молекули, які безперервно рухаються, володіють енергією. Саме вона визначає температуру системи та слугує мірою інтенсивності тепла. Зміна температури організму впливає, по-перше, на швидкість хімічних реакцій, які визначають обмін речовин і перетворення енергії, а по-друге, приводить до змін структури молекул, які визначаються так званими слабкими силами або слабкими взаємодіями — водневі зв’язки, сили Ван-дер-Ваальса, іонні зв’язки, гідрофобні взаємодії. Ці зв’язки малоенергетичні. Тому вони, на відміну від ковалентних («сильних») зв’язків, розриваються й утворюються під дією кінетичної енергії атомів та молекул, і це не потребує впливу ферментів. «Слабкі» зв’язки за фізіологічних температур здатні розриватися й утворюватися знову. З підвищенням температури кількість порушених «слабких» зв’язків збільшується й за досягнення верхньої межі перенесення температури твариною відбувається дезорганізація молекул, які визначають основні функції організму, що викликають його загибель. Основні структури і процеси, що залежать від «слабких» хімічних зв’язків: 1) структура води; 2) третинна і четвертинна структури білків; 3) комплекси ферментів з лігандами; 4) взаємодія між ліпідами; 5) взаємодія між ланцюгами нуклеїнових кислот; 6) взаємодія між нуклеїновими кислотами; 7) взаємодія нуклеїнових кислот із білками; 8) зв’язування гормонів білковими рецепторами тощо.
На відміну від «слабких» зв’язків, ковалентні зв’язки за фізіологічних температур стійкі й розриваються або утворюються під дією ферментів. Вони зменшують енергію активації (порогове значення енергії) хімічних реакцій. За звичайних фізіологічних температур швидкість ферментативних реакцій в 108— 1012 разів вища, ніж аналогічних реакцій без ферментів.
Функціонування ферментативних систем, у свою чергу, забезпечується значною мірою «слабкими» взаємодіями, оскільки саме вони формують утворення фермент-субстратних комплексів, а після перебігу реакції — повернення конформації білків-ферментів у висхідне положення. Приєднання регуляторів активності напряму перебігу ферментативної реакції теж зумовлюється «слабкими» силами. Саме ці сили визначають структурну «гнучкість» ферментів, що й робить ці каталізатори хімічних реакцій чутливими до змін температури.
За зниження температури метаболічні реакції можуть настільки загальмуватися, що життєво необхідні процеси не зможуть відбуватися з необхідною швидкістю. Навпаки, підвищення температури може викликати таке прискорення метаболічних процесів, що постачання організму достатньої кількості необхідних метаболітів у певний час стане неможливим.
У процесі еволюції більшість організмів виробили здатність підтримувати певний рівень метаболізму незалежно від температури свого тіла, а інші — згладжувати вплив зміни температури відповідною регуляцією біохімічних процесів.
Для визначення температурних ефектів широко застосовується показних Q10, який характеризує зміну швидкості якогось процесу за підвищення температури на 10 °С. Так, якщо швидкість збільшується вдвічі, то Q10= 2, втричі — Q10= 3 і т. д.
Відомо, що частота появи реакційноздатних молекул залежно від температури описується рівнянням Лрреніуса:
![]()
де K — константа швидкості; Т1 — початкова температура, Т2 — кінцева температура; Еа — енергія активації; R — універсальна газова стала.
Відношення двох швидкостей за різниці температури 10 °С (Q10), виходячи з рівняння Лрреніуса, визначається
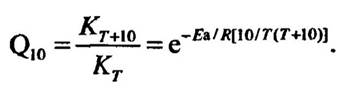
Отже, Q10 є функцією температури, і значення цього показника зменшується з її збільшенням.
Вплив температури на структуру білків. Основним фактором, який стабілізує третинну структуру (загальне розміщення, упакування ділянок, доменів і окремих амінокислот у поліпептидний ланцюг) і четвертинну (утворення агрегатів з двох і більше поліпептид- них ланцюгів) структуру білків (поряд з пептидними і дисульфід- ними зв’язками) є «слабка» взаємодія. Водневі зв’язки також відіграють вирішальну роль у стабілізації вторинної структури (наприклад, α-спіральних ланок) — просторовій конфігурації білкової молекули.
Зміни третинної структури білків під впливом температури мають, як правило, постійний характер, відбуваються в широкому
діапазоні температур і в більшості випадків не призводять до втрати основної функціональної здатності білків. Зміна четвертинної структури білків зазвичай супроводжується втратою функціональної активності білків. У разі низької температури тільки деякі ферменти холоднокровних тварин не втрачають своєї четвертинної структури. Серед усіх «слабких» зв’язків до низьких температур найчутливішими є гідрофобні взаємодії, які беруть безпосередню участь у стабілізації агрегатів з поліпептидних ланцюгів. У багатьох випадках порушенню четвертинної структури передує зміна третинної. Саме зміна третинної структури сприяє деградації білкових субодиниць.
Зміна температури впливає також на структурно-функціональні властивості макромолекулярних білкових комплексів, які утворюються в результаті агрегації різних білків, кожний з яких має всі чотири рівні структурної організації. Структура мембранних ліпідів, ліпід-білкові взаємодії також залежать від температури. Чутливою до температури є конформація поліпептидного ланцюга, що синтезується комплексом полірибосом. Поліпептид, який ще не набув своєї кінцевої конформації, може бути навіть чутливішим до зміни температури, ніж цілком сформований поліпептидний ланцюг.
Температурну залежність має також утворення комплексів білків-ферментів із субстратами та регуляторами їхньої активності, які зумовлені «слабкими» взаємодіями.
Вплив температури на структуру нуклеїнових кислот. Наслідком розриву «слабких» хімічних зв’язків нуклеїнових кислот може бути порушення їхньої вторинної структури. Прикладом слугує втрата структури молекули ДНК, у якої два ланцюги утримуються завдяки водневим зв’язкам між основами в парах гуанін— цитозин (Г — Ц) і аденін — тимін (А — Т), а також гідрофобним взаємодіям, завдяки яким кільцеві структури пуринів і пиримідинів відтискаються у внутрішню безводну ділянку молекули. За такої структури заряджені фосфатні групи контактують із водою.
Підвищення температури до рівня, що сприяє розриву «слабких» зв’язків, особливо водневих, може викликати втрату вторинної структури нуклеїнової кислоти. Порушення дволанцюгової структури ДНК унаслідок дії підвищеної температури, яку називають температурою плавлення ДНК, зростає також під впливом ультрафіолетового випромінювання внаслідок відкритого положення кільцевих структур пуринових і піримідинових основ.
Температура «плавлення» дволанцюгової ДНК пропорційна вмісту пар Г — Ц у молекулі. Кожну пару Г — Ц стабілізують три водневі зв’язки, а А — Т — два. Отже, дволанцюгові ДНК, які мають більший вміст пар Г — Ц, стабільніші до дії підвищених температур.
Температури «плавлення» дволанцюгових ДНК, як правило, значно вищі, ніж ті температури, за яких здатний виживати сам організм. Тому термічне порушення вторинної структури ДНК, очевидно, не має великого біологічного значення. Проте зміна третинної структури транспортної РНК (тРНК) за підвищення температури має істотне значення для виконання цією нуклеїновою кислотою своєї функції. За таких змін порушується процес приєднання амінокислоти до відповідної тРНК. Цей процес каталізується аміноацил-тРНК- синтетазами за участю АТФ. Для кожної амінокислоти є своя специфічна аміноацил-тРНК-синтетаза і специфічний тРНК-акцептор (іноді для однієї амінокислоти може бути кілька тРНК). На поверхні ферменту існують певні ділянки, які «впинають» певну амінокислоту, специфічну тРНК і АТФ. Втрата цієї здатності та зв’язування субстратів аміноацил-тРНК-синтетазою призводить до порушення синтезу білка. У дослідах із бактерією Micrococcus criophilus показано, що ферменти адаптованої до тепла бактерії здатні «впізнавати» конформацію тРНК, яка виникає за високої температури, а адаптовані до холоду — тільки «низькотемпературну» конформацію тРНК. Установлено також, що у термофільних бактерій зміна температури від оптимального рівня може призвести до зниження точності трансляції мРНК-залежної стадії синтезу білка.
Зауважимо, що всі компоненти білоксинтезуючої системи (молекули тРНК, мРНК, відповідні ферменти й рибосоми) здатні до температурних конфірмаційних змін, які можуть негативно впливати на життєдіяльність організмів.
Регуляція активності генів, яка визначається «слабкими» взаємодіями між нуклеїновими кислотами й білками, також залежить від температури. Її зміна може, наприклад, перешкоджати включенню або виключенню певного гена. Крім цього, можлива безпосередня активація генів у разі дії певних температур, які кодують саме ті білки або РНК, які необхідні за зміни температури.
Вплив температури на ліпіди мембран і активність мембранозв’язних ферментів. Важливу роль у визначенні структурно- функціональних властивостей біологічних мембран відіграють ліпіди. Вони не тільки підтримують структуру мембран, беруть участь у транспорті речовин, а й регулюють активність мембранозв’язаних білків, зокрема ферментів. В основу такої дії мембранних ліпідів покладено гідрофобні взаємодії між неполярними ланцюгами жирних кислот і залишками гідрофобних амінокислот, які експоновані на поверхні мембранозв’язаних білків. Білкові молекули, які прилягають до поверхні мембрани (периферійні білки), здатні утримуватись іонними взаємодіями між зарядженими групами на поверхні білків і зарядженими (полярними) «головками» фосфоліпідів мембран.
Як уже зазначалося, зміни температур порушують «слабкі» сили, які саме й визначають взаємодії між білками й ліпідами в мембранах.
За зміни температури відбувається порушення фізичного стану мембран, яке насамперед пов’язане з їхньою мікров’язкістю, що визначається, головним чином, ступенем насиченості ліпідних молекул. Що вища ненасиченість ліпідів, тобто збільшується кількість подвійних зв’язків між атомами карбону в жирнокислотних ланцюгах, то нижче точка «плавлення» ліпідів. Подвійні зв’язки створюють згини в аліфатичних ланцюгах й ускладнюють кристалізацію. Тільки за низьких температур «слабкі» зв’язки, які стабілізують взаємодію між молекулами ліпідів, стають достатньо міцними для того, щоб високоненасичені ліпіди могли затвердіти.
Установлено, що в організмах, які адаптувалися до хлору, зменшується насиченість жирних кислот. У свою чергу, ненасичені жирні кислоти слугують активаторами мембранозв’язаних ліпопротеї- нових ферментів. Отже, зменшення ступеня насиченості мембранних ліпідів за дії низьких температур може призводити до компенсаторної зміни активності мембранозв’язаних ферментів.
БІОХІМІЧНА СТІЙКІСТЬ ТВАРИН ДО ВИСОКИХ І НИЗЬКИХ ТЕМПЕРАТУР. Тварини, як уже зазначалося, значно розрізняються щодо оптимальних для них температур. Одні здатні існувати тільки у вузькому діапазоні температур довкілля, а інші — у набагато ширшому. Крім цього, можлива певна адаптація до екстремальних температур — після довготривалого перебування тварини в умовах, близьких до межі її температурного виживання, ця межа може бути зсунута. Варто зауважити, що летальна температура для даного організму залежить від часу дії — тварина може витримувати якусь температуру flfiz /fi- певного часу, але гине, коли її вплив триває довго.
Стійкість до високих температур. Тварини, як уже зазначалося, не здатні пройти весь життєвий цикл за температур понад 50 °С, але на окремих стадіях розвитку вони можуть переносити значно вищі температури. Так, наприклад, личинки мухи Polypedilum за зневоднення у висушеному стані виживають за температури 102 °С протягом хвилини, після чого здатні рости й зазнавати метаморфоз. Інший приклад — яйця прісноводного рачка Triops зберігають життєздатність у сухому мулі за температури 80 °С, а у воді за нормальних умов — за температури 99 °С (на 1 °С нижче температури кипіння води).
Хоча верхня межа активної життєдіяльності організмів близька до 50 °С, багато тварин гине за значно нижчих температур, особливо водні організми. Навіть у тропічних морях температура рідко сягає 30 °С, хоча в мілких закритих бухтах і лагунах вона буває й вищою.
Черевоногі молюски, які живуть у верхній смузі приливної (літоральної) зони і довше інших перебувають у повітряному середовищі, стійкіші до дії підвищеної температури, ніж ті, що живуть у нижній частині приливної зони, в якій температура близька до мінімальної. Так, наприклад, у приливній смузі мешкають Tecturius vilis, які протягом години здатні переносити температуру 48,5 °С, водночас у нижній частині цієї самої приливної смуги мешкають Tegula lisschkei, які за цей термін можуть переносити температуру не більш як 39 °С.
Серед риб найстійкішими до високих температур є, мабуть, коропозубики. Так, наприклад, Cyprinodon diabolus здатні мешкати у воді, температура якої 33,9 °С. Верхня температурна межа для дорослих особин цих рибок, маса яких близько 200 мг, становить приблизно 43 °С.
У порушенні життєдіяльності тварин за дії високих температур істотне місце належить: 1) денатурації білків, коагуляції їх під час нагрівання; 2) термічній інактивації ферментів, яка відбувається з більшою швидкістю, ніж їх синтез; 3) нестачі оксигену; 4) різниці в температурному коефіцієнті (зокрема, Qio) для взаємозв’язаних метаболічних процесів; 5) зміні структури мембран.
Разом з тим для вже розглянутих риб Tremantomus важко уявити, що летальна для них температура вище 6 °С може викликати термічну денатурацію білків, яка зазвичай відбувається за температури 45— 55 °С. Це ж стосується й температурної інактивації ферментів. Нестача оксигену в умовах, коли його використання підвищується зі збільшенням температури, не є визначальним фактором загибелі організмів. Так, наприклад, риби, які мешкають у холодній воді, гинули в теплій воді навіть тоді, коли вміст оксигену збільшували в кілька разів шляхом аерації води оксигеном.
Основними причинами загибелі тварин за дії підвищеної температури є, очевидно, різниця в температурній чутливості великої кількості ферментів, які беруть участь в обміні речовин та перетворенні енергії, а також зміна структури мембран, що призводить до численних порушень конформації білків, взаємодій між білками та ліпідами тощо.
Стійкість до низьких температур. Тварини, які мешкають у помірному й холодному кліматі, витримують довготривалу дію температур, які набагато нижчі за точку замерзання чистої води (0 °С). Вони запобігають холодовому ушкодженню за допомогою двох основних механізмів: переохолодження (зниження температури рідин в організмі нижче точки її замерзання без утворення льоду і толерантності (лат. tolerantia — терпимість) до заморожування (здатність переносити замерзання води й утворення льоду в організмі). Імовірність замерзання переохолоджуваної води залежить від температури, наявності центрів утворення льоду та часу. За відсутності чужорідних частинок, які слугують центрами кристалізації, точку замерзання чистої води можна знизити до -20 °С, а іноді навіть -40 °С.
Здатність тварин до переохолодження має велике значення для їхнього виживання, особливо в умовах раптового похолодання. Деякі плазуни, у яких температура замерзання рідин тіла близько -0,6 °С, здатні витримувати переохолодження до -8 °С без замерзання.
Внутрішньоклітинне замерзання води призводить до загибелі клітин у результаті ушкодження їхньої ультраструктури. Водночас багато організмів здатні витримувати позаклітинне утворення льоду. З перетворенням позаклітинної води у лід у ній концентруються розчинені речовини. Це створює великий осмотичний градієнт, який відсмоктує воду з клітин, що може викликати їх зневоднення. У більшості організмів клітини гинуть після переходу приблизно 2/3 клітинної води в позаклітинний простір.
Однак деякі тварини здатні виживати за утворення льоду в позаклітинному просторі. Так, личинки комара Chironomus витримують численні заморожування за температури -25 °С і відтавання. У більшості тварин існують адаптаційні механізми, які дозволяють уникнути руйнівних наслідків утворення льоду. Основні з них: 1) зниження точки замерзання рідин тіла; 2) підвищення здатності цих рідин до переохолодження без утворення льоду; 3) якщо неможливо запобігти утворенню води в позаклітинній рідині, то ушкодження клітин можна уникнути за рахунок зменшення осмотичного переходу води в позаклітинний простір.
Зауважимо, що тривалість дії низької температури теж впливає на здатність рідини тіла організмів до замерзання. Так, наприклад, у личинок хлібного пильщика Cephus cinctus за температури -15 °С лід не утворюється, а за нижчих температур ці личинки замерзають. Так, за температури -30 °С це відбувається лише за 1,2 с, а за -17 °С 50 % особин промерзають протягом року.
Безхребетні тварини приливної зони субполярних областей здатні переносити численні зміни температури нижче й вище точки замерзання води. Під час відливів ці тварини промерзають так, що більша частина води в їхньому організмі перетворюється в лід. Вони протягом 6 год і більше можуть перебувати при температурі -30 °С, за якої 90 % води в організмі замерзає, а в тій, що залишилася, концентрація розчинених речовин збільшується в багато разів, що попереджує її замерзання. Клітини таких тварин утрачають частину води і зморщуються, але в них вода не замерзає завдяки концентрації в ній розчинених речовин.
Мідія Mytilus edulis здатна переносити температуру до -10 °С. Це зумовлено частково осмотичними властивостями її внутрішньоклітинних рідин. У цього молюска близько 20% клітинної води не може бути вилучено з клітин за умов утворення внутрішньоклітинного льоду. Напевно, у клітинах таких організмів містяться речовини, осмотична дія яких проявляється значно сильніше, ніж це могло б бути зумовлено розчинними неорганічними й невеликими за розмірами органічними речовинами (амінокислотами, цукрами тощо). До таких речовин належать, зокрема, гліцерол, а також глікопротеїни антарктичних і арктичних риб.
Гліцерол знижує точку замерзання й межу переохолодження рідин тіла тварин унаслідок взаємодії його гідроксильних груп з полярними молекулами води завдяки утворенню водневих зв’язків. Ця сполука міститься у відносно високій концентрації в деяких комах, що сприяє зменшенню утворення льоду за переохолодження, а також захисту ушкоджених льодом тканин. У деяких комах концентрація гліцеролу зростає перед настанням зими. Так, наприклад, в їздця Bracon cephi кількість гліцеролу перед зимою збільшується майже на 30 % і становить 5 молей на 1 кг води. Завдяки цьому температура замерзання гемолімфи знижується до -17,5 °С. Личинка цієї комахи може бути переохолоджена (без утворення льоду) до - 47,2 °С.
Толерантність комах до холоду пояснюється не тільки збільшенням вмісту в їхній гемолімфі гліцеролу. Існують комахи, у яких концентрація цієї сполуки не перевищує 3 % або гліцеролу зовсім немає. Проте ці комахи захищені від ушкоджень під час промерзання. Крім цього у більшості арктичних комах, наприклад у туруна Pterostichus brevicomis, точка граничного переохолодження знижується наприкінці літа ще до того, як вміст гліцеролу в організмі збільшиться. Більше того, усередині зими вміст гліцеролу зменшується, а ступінь толерантності до замерзання в цей період не знижується.
Отже, крім гліцеролу існують також інші компоненти гемолімфи, які захищають комах від ушкоджуючої дії промерзання.
У деяких антарктичних і арктичних риб виявлено глікопротеїни, основна функція яких полягає у зниженні точки замерзання біологічних рідин. Яків гліцеролі, у таких глікопротеїнах, що їх називають ще «антифризними» білками, гідроксильні групи утворюють водневі зв’язки з молекулами води. Блокування цих груп, наприклад
шляхом їх ацетилювання, призводить до повної втрати «антифризної» здатності, і в цьому разі вони знижують точку замерзання води у цьому випадку не більше, ніж звичайні розчинені речовини (амінокислоти, глюкоза, сахароза тощо).
Риби родини Nototheniidae містять глікопротеїни-антифризи. Структуру одного з них, молекулярна маса якого близько 10 500 Да, наведено на рис. 8.3.
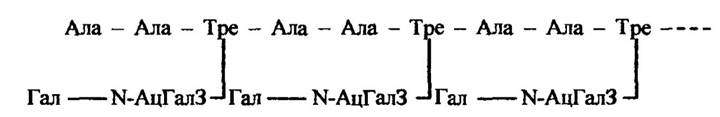
Рис. 8.3. Будова основної повторювальної одиниці глікопротеїнів-антифризів у риб Nototheniidae
Примітка. Ала — аланін, Тре — треонін, Гал — галактоза, N-АцГалЗ- N-ацилгалактозамін.
У риб даної родини знайдено також інші глікопротеїни-антифризи, зокрема з молекулярною масою 3500 і 2600 Да, у яких на відміну від наведеного на рис. 8.3 глікопротеїну залишки аланіну в певних ланках поліпептидного ланцюга заміщені залишками проліну.
У крові антарктичних риб роду Trematomus концентрація глікопротеїнів-антифризів прямо пропорційна її здатності до замерзання. Найвища вона в Trematomus borchgrevinki — пелагічної риби, яка мешкає в зонах з плавучим льодом, що утворюється під суцільним морським льодом, і яка спускається нижче тільки для виловлювання планктонових організмів.
У крові Trematomus глікопротеїн-антифриз міститься у трьох формах з молекулярною масою 10 500, 17 000 і 21 500 Да. Він, як і глікопротеїн-антифриз Nototheniidae, складається з повторювальної одиниці (рис. 8.3), яка містить дві амінокислоти — аланін (23 %) і треонін (16 %) — та приєднану до треоніну похідну галактози.
За механізмом дії глікопротеїни-антифризи не тільки структурують воду в результаті утворення водневих зв’язків із гідроксильними групами цих глікопротеїнів, а й гальмують ріст утворених кристаликів льоду за рахунок адсорбції на їхній поверхні, що попереджує приєднання нових молекул води до кристалічної гратки. Утворена глікопротеїнова оболонка навколо дрібних льодових кристалів може настільки послабити структуру льоду, що він тане.
Під час обговорення проблеми стійкості й толерантності до замерзання основну увагу приділено питанням утворення льоду та наслідкам цього для організму. Однак утворення льоду — це тільки один із можливих проявів дії низьких температур. Існують також інші негативні наслідки для організмів, зокрема зміни третинної та четвертинної структури білків унаслідок порушення гідрофобних взаємодій. Деякі речовини, зокрема гліцерол, здатні на тільки знижувати точку замерзання води, а й стабілізувати гідрофобні взаємодії, а саме білків, у тому числі й ферментів.
Зауважимо, що крім біохімічних способів адаптації до низьких температур деякі тварини здатні запобігати замерзанню за допомогою реакцій поведінки, анатомічних і фізіологічних засобів. Багато тварин мігрують перед настанням зими з місць, де їм загрожує замерзання. Це можуть бути міграції довжиною в тисячі кілометрів або ж у межах звичайної для них географічної зони. Так, наприклад, тварини, які впадають у зимову сплячку, можуть зариватися в землю і в такий спосіб уникати дії низьких температур. Деякі морські риби, наприклад слиж Salvelinus alpinus, на зиму мігрують із морської води, де температура може впасти нижче точки замерзання для рідин тіла, у прісноводні річки та озера, в яких температура у природних шарах не знижується менш як до 4 °С. Певні риби, які мешкають на мілководних ділянках моря, зимують у природних шарах із метою запобігання контакту з плаваючим і поверхневим льодом.
Нерідко межі температурної стійкості тварин неоднакові взимку і влітку. Взимку такі тварини залишаються активними за такої температури, яка влітку для них смертельна, і, навпаки, стійкість до високих температур взимку нижча, ніж улітку. Отже, тварини здатні акліматизуватися до температурних змін. Існують як географічна, так і сезонна адаптація й акліматизація. Так, наприклад, для жаби Rana sylvatica оптимальна температура води в період розмноження — 10 °С, нижня межа температури води для розвитку ікри — 2,5 °С, верхня — 24 °С.
Нижня летальна температура також змінюється залежно від пори року. Наприклад, якщо жуків Pterosticus brevicomis, які мешкають на Алясці, заморозити влітку (температура -6,6 °С), то вони гинуть. Взимку ці жуки виживають за температури, нижчої —35 °С.
Тож у тварин існують генетично зумовлена стійкість, яка зв’язана з їхнім геоірафічним поширенням і сезонними змінами.
РЕГУЛЯЦІЯ ТЕМПЕРАТУРИ ТІЛА. Температура будь-якого тіла залишається незмінною лише тоді, коли приплив тепла та його втрата (віддавання) рівні між собою. Як уже зазначалося, за умов дії низьких температур тепловий баланс тварин стабілізується як зменшенням віддавання тепла, так і збільшенням його припливу (виробітку). Більшість ссавців і птахів здатні підтримувати необхідну для життєдіяльності стабільну температуру. Однак у деяких із них за певних умов температура тіла різко знижується і вони заклякають або впадають у зимову сплячку. Проте й у цьому разі регуляція температури тіла цілком не припиняється.
За підвищення температури довкілля для підтримування температури тіла необхідно, щоб організм цьому перешкоджав.
Теплообмін між організмом і довкіллям відбувається трьома основними шляхами: 1) теплопровідність (пов’язана з прямим передаванням кінетичної енергії молекул); 2) випромінювання; 3) випаровування. Вони залежать від низки зовнішніх факторів, серед яких найважливішим є температура. За зниження температури довкілля тепловіддача зростає, а за її підвищення — зменшується. Якщо зовнішня температура вище температури поверхні тіла, то передавання тепла шляхом теплопровідності і випромінювання іде в напрямку з довкілля в організм. При цьому загальний притік тепла в організм буде складатися з тепла, що надходить з довкілля, та з метаболічної теплопродукції. Для підтримання сталої температури тіла у цьому разі необхідно збільшити випаровування, щоб воно віднімало все надлишкове тепло.
Регуляція температури тіла ссавцями і птахами. Тепло, яке виробляється організмом тварини, може віддаватись у довкілля, але воно спочатку має перенестися до поверхні тіла. Тому температура поверхні тіла повинна бути нижчою, ніж внутрішніх органів, оскільки інакше перенесення тепла не буде.
В організмі ссавців продукування тепла різними органами й системами органів неоднакове. Так, наприклад, у людини внутрішні органи грудної та черевної порожнини становлять за масою лише 6 % маси тіла, а виробляють до 56 % усієї теплопродукції організму. Головний мозок продукує до 16 % теплоти організму людини. Шкіра і м’язи продукують не більш як 18 % усього тепла. Під час фізичної роботи теплопродукція за рахунок інтенсифікації метаболізму різко зростає. Для того щоб за фізіологічного навантаження внутрішня температура залишалася сталою, перенесення тепла до поверхні тіла має зрости порівняно зі станом спокою.
Органи ссавців, які виробляють більше тепла, можуть бути теплішими за інших, хоча вони охолоджуються кров’ю (венозна кров, яка відтікає від органів, тепліша, ніж артеріальна). Різниця в температурі може сягати 0,5 °С.
У людини за збалансованого теплообміну температура покривів тіла нижче внутрішньої температури. Це означає, що венозна кров, яка тече від поверхневих тканин, холодніша, ніж артеріальна. Це є одним з основних шляхів охолодження внутрішніх органів. Залежно від температурних умов довкілля температура шкіри схильна до значних коливань.
Внутрішня температура людини та інших ссавців, а також птахів піддається добовим коливанням, зазвичай у межах 1—2 °С. У денних тварин вона максимальна вдень і мінімальна вночі, а в нічних — навпаки. Цей добовий ритм не пов’язаний безпосередньо зі зміною стану активності та спокою, оскільки він зберігається й тоді, коли тварина постійно перебуває в повному спокої. У багатьох ссавців і птахів добові коливання температури тіла відповідають світловим і темновим періодам доби.
Отже, для того щоб вимірювання температури тіла ссавців і птахів було точним, необхідно мати інформацію про активний стан тварини, час доби, цикл температури тіла тощо.
Якщо знехтувати різницею 1—2 °С, то можна вважати, що кожній групі теплокровних хребетних тварин притаманна стала, «нормальна» для них температура. Як правило, температура тіла більшості птахів становить 40 ± 2 °С, у плацентарних ссавців — 38 ± 2°С, у сумчастих — 36 ± 2 °С, вищих ссавців — 36 ± 2 °С. Летальна температура приблизно на 6 °С вища, ніж «нормальна» температура тіла.
Зауважимо, що ссавці і птахи, які мешкають в умовах низької температури Арктики й Антарктики, підтримують температуру свого тіла на тому самому рівні, який притаманний кожному з класів у цілому, незважаючи на існування в холодних районах.
Терморегуляція за дії низьких температур у ссавців. Для підтримування сталої температури тіла тварин за дії холоду продукування тепла в процесі метаболізму, як уже зазначалося, має врівноважуватися тепловіддачею і може бути вирахуване за формулою:
![]()
де Н — кількість тепла, яка продукується в метаболічних процесах; Q — тепловіддача, С — коефіцієнт пропорційності, який слугує показником теплопровідності, Тт — температура тіла, Tg — температура довкілля.
У наведеному рівнянні Tg (температура довкілля) не залежить від тварини, якщо вона не переміщується в інше середовище. Для пристосування до несприятливої зовнішньої температури тварина має змінювати інші величини — особисту теплопродукцію (Н), теплопровідність (Q або температуру тіла (ТУ). Оскільки ТУ має підтримуватися сталою, то тварині залишається можливість змінювати власну теплопродукцію або теплопровідність. Зміну температури тіла тварин, які впадають у зимову сплячку або заціпеніння, буде розглянуто нижче.
Підвищення теплопродукції (рівня метаболізму) може використовуватися в широких межах. Основні способи збільшення теплопродукції: 1) м’язова активність; 2) мимовільне скорочення м’язів, за якого розщеплення АТФ актин-міозиновою АТФазною системою відбувається без виконання роботи (тремтіння); 3) підвищення метаболізму без скорочення м’язів (термогенез без тремтіння). Один з механізмів останнього процесу, який активується гормонами щитовидної залози, полягає в тому, що за участі Na+, K+-АТФазної системи калоригенних (які продукують тепло) тканин — печінки, скелетних м’язів і нирок — відбувається перетворення енергії АТФ у тепло.
За низьких температур довкілля має бути збільшена інтенсивність метаболізму, щоб температура тіла залишалася сталою. Установлено, що для більшості ссавців за температури нижче певної точки, яка називається нижчою критичною температурою (Тнк), інтенсивність метаболізму зростає лінійно зі зниженням температури. Вище цієї критичної температури продукування тепла залишається незмінним, оскільки воно не може бути нижчим, ніж рівень у стані спокою. Для більшості тропічних ссавців нижня критична температура лежить у межах 20 — 30 °С і за температур, нижчих цієї критичної, інтенсивність метаболізму швидко зростає зі зниженням температури. Для цих тварин характерна висока теплопровідність. У них метаболізм різко зростає навіть за помірних знижень температури.
Для ссавців Арктики й Антарктики характерна низька критична температура. Так, наприклад, у песця рівень метаболізму помірно зростає тільки за температури повітря, нижчої -40 °С. Арктичні тварини характеризуються низькою теплопровідністю. У ссавців Арктики набагато ширша термонейтральна зона (діапазон температур, в якому інтенсивність метаболізму не залежить від температури), а також підвищення рівня метаболізму за дії холоду значно менше, ніж у тропічних тварин.
Теплопровідність (потік тепла за одиницю часу через одиницю площі в розрахунку на 1 °С різниці температур) у широкому розумінні є мірою потоку тепла від тварини в довкілля за дії низьких температур. Це охоплює також перенесення тепла від внутрішніх органів до поверхні шкіри й від шкіри через хутрові покриви, якщо вони є, у довкілля. Низька теплопровідність відповідає високій теплоізоляції і навпаки.
Один із засобів підвищення теплорегуляції — наявність хутра. Теплоізоляційні властивості хутра зростають зі збільшенням його товщини, досягають максимуму у відносно великих за розмірами тварин, наприклад таких, як песець. У дрібних тварин спостерігається кореляція між товщиною хутра (його теплоізоляційними властивостями) і розмірами тварин. Для можливості швидкого пересування дрібні тварини повинні мати тонке й легке хутро. Це повною мірою стосується її найдрібніших ссавців, зокрема мишоподібних гризунів і землерийок. Цим тваринам через відносно погану теплоізоляцію доводиться відшукувати необхідні їм кліматичні умови (наприклад, жити в норах) або за низьких температур упадати в сплячку, щоб не підтримувати високу температуру.
Відомо, що густина хутра у тварин залежить від пори року: взимку хутро густіше і краще зберігає тепло, ніж легше хутро влітку. Сезонні зміни найзначніші у великих тварин, а в дрібних гризунів вони виражені меншою мірою. У чорного ведмедя, наприклад, теплоізоляційні властивості хутра влітку приблизно на 52 % нижче, ніж узимку.
У тюленів, які мають відносно тонке хутро, теплоізоляцію створює також товстий шар підшкірного жиру. Тому різниця в теплоізоляційних властивостях шкіри тюленя в повітрі й у воді невелика.
Тюлені й кити подібні до інших теплокровних тварин — температура їхнього тіла зазвичай 36 — 38 °С. У більшості цих тварин рівень метаболізму майже не змінюється за охолодження води аж до точки замерзання. Тому основний спосіб пристосування до низьких температур — ефективна теплоізоляція за рахунок товстого шару підшкірного жиру, який перешкоджає втраті тепла. Температура поверхні шкіри цих тварин майже дорівнює температурі холодної води. У тюленів на глибині шару підшкірного жиру близько 50 мм температура відповідає внутрішній температурі тіла.
Більшість наземних арктичних ссавців мають інший градієнт температури: поверхня шкіри під хутром у них лише на кілька градусів холодніша, ніж внутрішні ділянки тіла, тобто шкіра зовні захищена теплоізоляційним шаром. У білого ведмедя, як уже зазначалося, хутро має низьку теплоізоляцію і тому підшкірний жир є основним теплоізоляційним матеріалом.
За потепління або прискорення метаболізму під час швидкого плавання тюлені, кити й подібні до них тварини уникають перегрівання завдяки тому, що за таких умов підсилюється прилив крові через підшкірний жир до поверхневого шару шкіри через систему кровоносних судин. Вона дає змогу відносно довго регулювати кількість тепла, яке потрапляє до поверхні шкіри та відводиться в довкілля.
Особливістю водних ссавців є те, що в них теплоізоляційний шар (підшкірний жир) розміщений глибше тієї поверхні, через яку відбувається тепловіддача. Завдяки цьому кров може потрапляти до цієї поверхні в обхід теплоізоляційного шару, і в теплій воді або за великих фізичних навантажень тепло відводиться поза цим шаром. Наземні ссавці, у яких теплоізоляційний шар міститься зовні поверхні шкіри, не здатні так ефективно регулювати втрату тепла з усієї поверхні шкіри, і їм необхідно використовувати також інші способи тепловіддачі.
Укриті хутром тварини Арктики й Антарктики неоднаковою мірою захищають від холоду всі частини тіла, оскільки мають залишатися певні відкриті ділянки поверхні для віддавання тепла, особливо за фізичного навантаження. Саме тому, мабуть, у наземних тварин існують ділянки тіла, які мають слабку теплоізоляцію (морда, лапи тощо). Так, наприклад, у південноамериканського гуанако Lama guanicoe приблизно 40 % тіла вкрито товстим шаром хутра, а 20% майже не мають хутрового покриву, що дозволяє цій тварині в широких межах регулювати тепловіддачу.
Деякі тварини регулюють свою тепловіддачу зміною пози. Гуанако, коли вони випрямляють лапи, здатні через їхню поверхню віддавати майже 50 % надлишкового тепла в довкілля. Коли ж гуанако лежить чи стоїть зі стуленими лапами, то площа, через яку відбувається віддача тепла, зменшується майже вдвічі. Якщо вона лежить і згортається так, щоб по можливості вкрити всі незахищені хутром місця, то тепловіддача в них зменшується майже в 5 разів порівняно з тепловіддачею тварини, яка розпрямляється.
Ласти та хвостові плавці тюленів і китів не мають підшкірного шару жиру. Водночас, у них існує розвинута сітка кровоносних судин, через які за участю крові можлива втрата великої кількості тепла. Проте завдяки особливостям структури кровоносної сітки в цих органах, що діє як теплообмінник, цього не відбувається. У ластах і плавниках тюленів і китів кожна артерія оточена венами. Артеріальна кров охолоджується венозною кров’ю, яка її оточує. Наслідком цього є те, що артеріальна кров потрапляє в периферійні ділянки в охолодженому стані і вони віддають холодній воді мало тепла. Разом з тим венозна кров нагрівається ще до того, як вона повертається у внутрішні органи.
Коли китоподібні тварини плавають у теплій воді, то виникає необхідність віддавати тепло. У цьому разі за збільшення приливу крові й артеріального тиску центральна артерія розширюється та стискає навколишні вени. Венозна кров за цих умов тече по інших венах і повертається у внутрішні ділянки тіла без попереднього підігрівання, що сприяє охолодженню тіла.
Теплообмінники існують не тільки в китоподібних. Так, наприклад, вони є в ламантинів, які мешкають у тропіках і субтропіках. Ці тварини мають відносно низьку інтенсивність метаболізму, тому для них важливе зменшення теплових втрат. Такі самі пристосування
притаманні деяким тропічним тваринам, зокрема лінивцям. У них артерія, розміщена паралельно лапам, розділяється на тонкі артерії, що стикаються з такою самою кількістю вен. У дощову й вітряну погоду у сплячих лінивців це дає змогу зменшити витрати тепла внутрішніх органів.
У людини в руках і ногах відбувається певний теплообмін між головними артеріями, які прилягають до вен, що розміщені в глибині тканин. Під дією холоду більша частина венозної крові повертається по цих венах, тимчасом як під впливом тепла венозна кров тече поверхневими венами, що містяться під шкірою, й завдяки цьому уникають теплообміну з артеріальною кров’ю.
Один з механізмів підвищення температури тіла ссавців і птахів, а також риб і земноводних — лихоманка. Вона викликається бактеріальною або вірусною інфекцією і пов’язана з продукуванням речовин, які називають пірогенами. За лихоманки терморегуляція зберігається цілком, але температура підтримується на вищому рівні. За певних умов, зокрема переохолодження, лихоманка, очевидно, має позитивний ефект.
Теплоізоляція в разі дії низьких температур у птахів. У більшості птахів існують механізми регуляції тепла, притаманні ссавцям, — подібна залежність рівня метаболізму від зовнішньої температури. Однак у птахів є певні відмінності. Зокрема, теплопровідність у них зменшується зі зниженням зовнішньої температури.
Як відомо, пір’я у птахів є запобіжником утрати тепла. Один із можливих способів зменшення теплопровідності — скуйовджування пір’я, утягування лапок і настовбурчення. Інший спосіб — охолодження периферійних тканин за підтримання сталої внутрішньої температури. У цьому разі знижується кровотік. Зменшення товщини периферійного шару призводить до зниження продукування в ньому тепла. Кожен із цих процесів сприяє як зниженню тепловіддачі, так і зменшенню інтенсивності метаболізму, але дає змогу в сукупності підтримувати внутрішню температуру тіла на сталому рівні.
Певне значення в збереженні температури тіла птахів має теплообмін у лапках, особливо в тих птахів, які стоять чи плавають у холодній воді. Цей теплообмін такий самий, як і у ссавців, — тепла артеріальна кров охолоджується венозною кров’ю, яка оточує її з усіх боків. Якби кров, яка тече до покритих тонкою шкірою поверхні лапок, не проходила через теплообмінник, утрати тепла були б значними, а так вони зводяться до мінімуму. Наприклад, у чайки, лапки якої перебували 2 год у льодяній воді, втрата тепла через шкіру становила приблизно 1,5 % загальної теплопродукції організму. У птахів доти, доки не виникає загроза обмороження, втрата тепла в лапках невелика, але на морозі ця втрата зростає пропорційно зниженню температури довкілля. Підвищена тепловіддача викликає необхідність посилення вироблення тепла за рахунок інтенсифікації метаболічних процесів.
Один зі способів зменшення тепловіддачі у птахів — скупчування (взаємне обігрівання). Він найхарактерніший для пінгвінів Антарктики, особливо для великих імператорських пінгвінів Aptenodytes forsteri, які мешкають у найхолодніших умовах і виводять пташенят узимку.
З наближенням зими ці пінгвіни покидають відкриту воду і прямують до своїх постійних місць розмноження на льодовому шельфі, який розміщений на відстані 50-100 км від відкритої води. Самка відкладає яйце, а самець поміщає його на лапки й обігріває, постійно залишаючись на одному місці до виведення пташеняти. Самка повертається на годування до відкритої води. Самець стоїть на льоду понад два місяці на морозі за температури, яка може сягати -30 °С і навіть -40 °С, за сильного вітру. Самка повертається з повним шлунком їжі в момент вилуплення пташеняти, годує його, а самець відправляється до відкритої води на годівлю.
Під час довготривалого голодування пінгвіни зберігають нормальну температуру тіла 38 °С. Вони підтримують свою життєдіяльність завдяки великим запасам підшкірного жиру. Великий самець імператорського пінгвіна перед переходом до місця розмноження має масу приблизно 35 кг, а під час голодування він втрачає близько 15 кг.
За теплопровідністю пінгвіни не відрізняються від інших птахів, у них нижня критична температура дорівнює -10 °С, що набагато вище температури для антарктичної зими.
Стоячим пінгвінам-самцям за низьких температур дає змогу виживати те, що вони стоять щільною юрбою. Коли тіла тварин, що стикаються, мають однакову температуру, то передавання тепла не відбувається. Отже, скупчення пінгвінів допомагає економити енергію й не дає їм можливості замерзнути.
Крім пінгвінів, скупчуватися можуть і інші тварини. Це зменшує величину відкритої поверхні і тим самим знижує дію холоду й затрати енергії на вироблення тепла. Це має особливе значення для пташенят і новонароджених ссавців, які мешкають разом, — вони здатні зберігати тепло за тимчасової відсутності батьків. Крім цього, зменшення витрат енергії на тепло сприяє прискореному росту.
Зимова сплячка й заціпеніння. Деякі ссавці і птахи впадають у сплячку, або гібернацію (лат. giberna — зима), узимку. Це дозволяє їм у цей період значно зменшити затрати енергетичних ресурсів. Під час зимової сплячки температура тіла тварин падає майже до рівня навколишнього середовища, що супроводжується зниженням інтенсивності метаболічних процесів, дихання, частоти скорочення серця тощо. Тварина в цей час перебуває в заціпенілому стані й слабко реагує на зовнішні подразники. Приготування до зимової сплячки супроводжується нагромадженням жиру, який є запасом енергетичного «палива».
У зимову сплячку впадають в основному дрібні тварини, які за притаманного їм метаболізму потребують інтенсивного харчування, зокрема гризуни (наприклад хом’яки, ховрахи, вовчки), комахоїдні (приміром, їжаки), кажани, певні сумчасті тощо, а також деякі птахи — колібрі, стрижі, дрімлюги тощо.
Деякі тварини, зокрема ведмеді, взимку впадають у сплячку, яка супроводжується Зниженням температури тіла лише на кілька градусів, рівень метаболізму й фізіологічних показників у них знижується незначно, самки нерідко народжують у цей період дитинчат. Цей стан не є повною зимовою сплячкою (істинною гібернацією).
Крім зимової сплячки в деяких тварин (наприклад кажанів, єхидни) можливий стан заціпеніння — зниження температури тіла та інтенсивності метаболізму короткочасно, на кілька годин. Ті самі кажани здатні впадати в зимову сплячку, а також і у стан заціпеніння.
Перехід у стан заціпеніння викликається припиненням терморегуляторного підвищення рівня метаболізму за зменшення температури повітря нижче критичної точки. Обмеження метаболізму до рівня, притаманного стану спокою, приводить до падіння температури тіла. У свою чергу, тільки-но температура тіла починає знижуватися, інтенсивність метаболізму гальмується і тварина впадає в заціпеніння.
Зимова сплячка й заціпеніння — процеси, які контролюються не тільки необхідністю економії енергії та кількістю їжі. Річні цикли координуються певним часом року, а добові — світловим часом доби. За цих станів деякі фізіологічні функції (наприклад, дихання) залишаються координованими, хоча й гальмуються. Якщо температура довкілля стає меншою, ніж це необхідно для координування подібних функцій, то тварина, яка впала в сплячку, гине. Однією з можливостей запобігання цьому явищу є вихід зі стану сплячки або припинення зниження температури тіла в результаті підвищення теплопродукції.
Підвищення температури тіла під час пробудження тварин, які впали в зимову сплячку або заціпеніння, пов’язане з найбільшими затратами енергії порівняно з іншими етапами життєдіяльності таких тварин. У цей період тварина дуже тремтить (скоротливий м’язовий термогенез).
У процесі розігрівання важливу роль відіграє особливий вид жирової тканини — бурий жир. Основна біологічна роль цієї тканини — генерація тепла. Вона має істотне значення як джерело тепла не тільки для тварин під час пробудження, а й за акліматизації до холоду в ранній постнатальний період. Ця тканина оточує життєво важливі органи й лежить на шляху крові, яка надходить до них.
Бурий жир становить не більше 1—2 % маси тіла. Однак він здатний підвищувати теплопродукцію організму до такого ступеня, що вона може сягнути третини всього утвореного в організмі тепла. За умов стимуляції цієї тканини симпатичною нервовою системою бурий жир здатний виділяти до 400 Вт тепла на 1 кг маси, що на кілька порядків більше звичайної термогенної здатності тканин ссавців (людина в стані спокою продукує приблизно 1 Вт тепла на 1 кг маси).
Основна кількість бурого жиру міститься у верхній частині спини, ближче до шиї. Вона оточує кровоносні судини, які живлять головний мозок кров’ю.
Своїм кольором бурий жир зобов’язаний наявності великої кількості кровоносних судин. Клітини цієї тканини містять велику кількість мітохондрій, у внутрішній мембрані яких зосереджені дихальні ланцюги, основною біологічною функцією яких є генерування енергії у формі градієнта електрохімічних потенціалів іонів водню (Дрн+) в окисно-відновних реакціях. У мітохондріях інших тканин енергія, запасена у формі Дрн+, використовується в основному Н+-АТФ-синтетазою для синтезу АТФ з АДФ інеорганічного фосфату (Фн):
![]()
У внутрішній мембрані мітохондрій бурого жиру вміст ферментів дихального ланцюга набагато перевищує вміст Н+-АТФ-синтетази. Крім того, у цій мембрані наявний особливий білок термогенін, який є переносником Н (протонофор). Його вміст становить 10—15 % загального білка мітохондрій клітин бурого жиру. Основна його функція — роз’єднання процесів окиснення в дихальному ланцюгу, що генерує енергію у формі Дрн+ з утворенням АТФ. В активованій тканині бурого жиру енергія окисно-відновних реакцій дихального ланцюга мітохондрій затрачається в основному не на синтез АТФ з АДФ і Фн, а розсіюється в тепло. Цей механізм є короткочасним і триває приблизно 15 хв. Функціонування тканин бурого жиру супроводжується посиленим поглинанням оксигену.
Під час виходу зі сплячки на початкових етапах пробудження спочатку розігрівається верхня частина тіла, де розміщені такі важливі органи, як серце і головний мозок, а потім задня. Підвищення температури серця має бути першим етапом пробудження, оскільки воно забезпечує циркуляцію крові та її надходження у всі інші органи. Саме тому основна маса бурого жиру міститься в передній частині тіла.
Тканина бурого жиру характерна для ссавців, які впадають у зимову сплячку. Водночас її немає у птахів, які регулярно впадають у сплячку, зокрема колібрі, стрижів, дрімлюг. У них пробудження відбувається за інтенсифікації інших метаболічних процесів, які координуються температурою, довжиною світлового часу доби тощо.
Регуляція температури тіла у водних холоднокровних тварин. Температура тіла більшості холоднокровних водних тварин, як і наземних, залежить в основному від температури довкілля. Однак у деяких з них вона може підтримуватися на вищому рівні, ніж температура води. Це може відбуватися за рахунок збільшення теплопродукції або зниження віддачі тіла.
Вода має велику, порівняно з повітрям, теплопровідність і теплоємність. Тому дрібні тварини швидко втрачають тепло й не можуть підняти температуру тіла істотно вище навколишньої води. Для того щоб значно збільшити теплопродукцію, таким тваринам необхідно підвищити споживання оксигену. Однак для цього необхідна велика поверхня зябер. Водночас кров, протікаючи крізь зябра, охолоджуватиметься до температури води, оскільки зяброва мембрана достатньо тонка для забезпечення дифузії оксигену й майже не перешкоджає віддаванню тепла.
У риб, які швидко плавають (наприклад, тунців, акули), між зябрами і тканинами тіла існує теплообмін, подібний до такого в ластах тюленів і хвостовому плавці китів. По боках тіла під шкірою цих риб розміщені судини, які постачають кров’ю м’язи, що забезпечують швидке плавання. Від головних судин відходять багато тонких артерій і вен, які утворюють щільне сплетіння й несуть кров у протилежних напрямах, — формується теплообмінник. Артеріальна кров, яка тече від зябер, має температуру води. Ця кров, що транспортується по оточених венами артеріях, поглинає тепло венозної крові від м’язів. Венозна кров, яка збирається у великі судини під шкірою, уже охолоджена, оскільки її тепло повернулося до м’язів через зігріту артеріальну кров. Наслідком цього може бути різниця температури м’язів тунця й води, яка здатна досягати 14 °С.
Підвищення температури м’язів дає таку перевагу, що тунець може плавати з великою швидкістю незалежно від температури води (у певному діапазоні). Крім цього, завдяки наявності теплообмінника, у тунця підтримується висока температура не тільки м’язів, а й інших органів, зокрема печінки та шлунково-кишкового тракту. Це прискорює перетравлення та всмоктування їжі. Деякі тунці підтримують також підвищену температуру головного мозку.
У деяких акул є теплообмінник, схожий на теплообмінник тунця, що забезпечує підтримання високої температури м’язів.
Регуляція температури тіла плазунами. Для підвищення температури тіла наземним тваринам необхідно зменшити випарювання і втрату тепла теплопровідністю, а також підвищити власну теплопродукцію та поглинання випромінювання. Як правило, тваринам вигідніше отримувати тепло з довкілля, ніж використовувати ресурси організму.
Сонячне тепло інтенсивно використовується плазунами, а також комахами. Поглинання сонячних променів сприяє забарвленню тіла тварин та їхній орієнтації стосовно Сонця. Багато плазунів здатні змінювати своє забарвлення шляхом концентрування або диспергування пігментів в особливих клітинах шкіри. Чорний колір найбільше зменшує відбиття сонячного світла у видимі області спектра.
Для збільшення освітленої поверхні тварини розміщують своє тіло під прямим кутом до сонячних променів, розпластуються та витягують кінцівки. Плазуни можуть зігріватися також, коли притискаються до теплих предметів оточення, наприклад нагрітого каміння.
Ящірки Liolaemus залишаються активними навіть за температури 2 °С. Якщо таку ящірку помістити на сонце, наприклад за температури 1,5 °С, то за 1 год температура її тіла, яка вимірювалася в клоаці, з 2,5 °С може піднятися до 33 °С і залишається такою, навіть коли температура повітря підвищується.
Плазунам притаманно наступне: при зігріванні частота скорочень серця висока і зростає з підвищенням температури тіла, а за охолодження зменшується. Це змінює циркуляцію крові й дає змогу пристосовуватися до різних температур довкілля.
На сонці гріються не тільки ящірки, а й інші плазуни — черепахи, змії, крокодили тощо.
Плазуни здатні для підвищення температури також посилювати власну теплопродукцію. Так, до цього способу вдаються самки деяких змій у період, коли вони зігрівають яйця, згортаючись навколо них. Під час висиджування яєць самки споживають більше оксигену за зниження температури, що сприяє посиленню метаболізму та підвищенню температури на 4—5 °С вище температури довкілля.
Регуляція температури комахами, що літають. Більшість комах за низьких температур стають в’ялими й не здатні літати. Проте деякі з них можуть розігрівати літальні м’язи та зберігати активність за холодної температури повітря. Ці м’язи, розміщені в грудному відділі тіла, здатні продукувати тепло в процесі, який подібний до тремтіння у ссавців. Таке розігрівання м’язів перед польотом характерне в основному для великих комах — наприклад, сарани, нічних метеликів, джмелів, ос, бджіл, які здатні до швидкого й довготривалого польоту. Так, приміром, джміль Bombus vagus завдяки безперервному тремтінню грудного відділу здатний зберігати температуру 32-33 °С, коли температура повітря становить від 9 °С до 24 °С.
У різних видів джмелів у літальних м’язах виявлено високу активність фруктозодифосфатази та фосфофруктокінази. Фруктозодифо- сфатаза зазвичай інгібується АМФ, а фосфофруктокіназа — активується. Однак фруктозодифосфатаза джмелів, на відміну від інших, не чутлива до АМФ. Тому в цих комах можливе одночасне функціонування цих ферментів, що приводить до утворення збільшеної кількості АТФ, результатом розщеплення якої є продукування тепла:
![]()
Джмелі підвищують свою температуру не тільки перед польотом, а й обігріваючи розплід. У цьому разі матка держить черевце притиснутим до стільника, оскільки температура черевця підтримується на рівні 31—35 °С. Навіть за температури повітря 5 °С матка джмеля зберігає температуру в стільниках близько 20 °С.
Медоносні бджоли Apis mellifera регулюють не тільки власну температуру, а й температуру всієї колонії. Оптимальна температура у вулику для розвитку розплоду становить приблизно 35 °С з невеликими відхиленнями. Якщо в літній період у вулику стає занадто тепло, бджоли розбризкують над стільниками воду, випарювання якої сприяє потокам повітря, що бджоли, котрі розмістилися на деякій відстані одна від іншої, створюють тріпотінням крил.
За низької температури бджоли зазвичай скупчуються, що сприяє зменшенню віддачі власного тепла. У великому діапазоні температури повітря від -17 °С до 11 °С температура у вулику підтримується на рівні 18 — 32 °С. У цьому аспекті родина бджіл веде себе як єдиний організм і підтримує свою температуру на високому рівні за умов, за яких окрема особина загинула б.
Ще один приклад пристосування комах до низьких температур — продукування тепла літальними м’язами на прикладі метелика- бражника Manduca sexta. Ця комаха веде нічний спосіб життя й нерідко літає за низьких температур. Вона подібно до колібрі харчується нектаром і зависає перед квіткою. Перед польотом бражники розігрівають тремтінням літальні м’язи грудного відділу, який укритий довгими й пухнастими лусочками, що допомагають зберігати тепло під час розігрівання м’язів. У процесі польоту температура грудного відділу підтримується на рівні 40 — 41 °С у широкому діапазоні зовнішньої температури.
Бражник, який летить, для запобігання перегрівання грудей посилює тепловіддачу обміном крові між грудним відділом і черевцем. Цій віддачі тепла сприяють відносно велика поверхня черевця та його слабка теплоізоляція.
Терморегуляція у тварин у разі дії високих температур. За надлишку тепла в організмі тварин його необхідно відводити в довкілля. Це можливо такими способами: 1) збільшенням надходження крові до шкіри, що прискорює перенесення тепла з внутрішніх ділянок тіла до поверхні; 2) відкриванням поверхонь тіла, особливо тих, де мало або немає покривів.
Коли температура довкілля вища, ніж тіла тварин, що буває, наприклад, у пустелях, то в організм переходить тепло з навколишнього середовища — від нагрітого повітря шляхом теплопровідності, нагрітої поверхні землі й Сонця поглинанням випромінювання. За цих умов надлишок тепла, що виробляється організмом у процесі метаболізму та поглинається з довкілля, має відводитися. Прилив тепла ззовні відбувається через поверхню тіла, тому теплове навантаження пропорційне площі цієї поверхні.
Дрібні за розміром тварини мають більшу стосовно об’єму поверхню. У разі дії підвищених температур це надає тваринам, які мають великі розміри, певну перевагу перед дрібними, оскільки вони на одиницю маси здатні поглинати менше зовнішнього тепла від випромінювання, ніж малі тварини. Однак у достатньо великих тварин подальше збільшення розмірів тіла не дає істотної переваги.
Розглянемо теплорегуляцію за дії високих температур у великої тварини на прикладі верблюда. У цієї тварини за достатньої кількості питної води кожної доби температура тіла коливається в межах 36-38 °С. Проте за нестачі води добові зміни температури тіла можуть значно коливатися. Уранці температура може сягати 34 °С, а до кінця дня — майже 41 °С. У прохолодну ніч нагромаджене тепло може розсіюватися випромінюванням без затрати води. Висока температура тіла протягом дня не тільки дає змогу акумулювати тепло, а й зменшує температурний градієнт між організмом і довкіллям. Це зниження надходження тепла в організм і також акумулювання тепла сприяють збереженню води, яка могла б затрачатися під час випаровування на охолодження тіла тварини.
Наявність у верблюда товстого шару шерсті, який має високі теплоізоляційні властивості, знижує надходження тепла з довкілля в організм. Ще одне із пристосувань верблюда до високих температур — здатність переносити значно більшою мірою, ніж багато інших тварин і людина, зневоднення організму. Так, наприклад, утрата води в кількості 10 — 12 % маси тіла людини може спричинити летальний кінець, а верблюд здатний переносити вдвічі більшу втрату води.
Дрібні тварини через малі розміри й відносно велику поверхню тіла стосовно маси мають бути нездатними вести активний спосіб життя в денний час доби за високих температур. Однак деякі з таких тварин живуть за цих умов. Прикладом є південоамериканські ховрахи. Ці дрібні тварини масою 100 — 200 г у денний час здатні перебігати з місця на місце, ховатися й вибігати з нори. Вони, як і верблюди, можуть акумулювати тепло, а також переносити високі температури. Так, ховрахи гинуть за температури 43 °С, але добре переносять температуру 42,4 °С. Тільки-но ховрах починає перегріватися на сонці, він одразу повертається у відносно прохолодну нору, де охолоджується. Часто забігаючи у прохолодну нору та виходячи з неї, ховрах швидко акумулює тепло й також швидко його позбавляється без випаровування води. Це й дозволяє ховраху бути активним упродовж дня.
Випаровування води, як уже зазначалося, є одним з основних способів позбавлення від надлишкового тепла в разі високих температур. Кількість тепла, необхідна для випарювання 1 л води, становить 2426 кДж. Основні шляхи підвищення випаровування води у тварин: 1) потовиділення; 2) прискорення дихання; 3) випаровування слини під час облизування хутра або лап.
У людини підвищення випаровування вологи потовиділенням є основним способом підтримування температури тіла в разі перегрівання. Деякі тварини, навіть вкриті шерстю та великі за розмірами (наприклад, бики, антилопи, верблюди), теж мають потові залози. У верблюда хутро дуже густе, але це не перешкоджає випаровуванню поту, оскільки повітря в пустелі дуже сухе.
Водночас деякі дрібніші тварини (вівці, кози тощо) майже не мають потових залоз і випаровування вологи в них підсилюється завдяки частотному диханню — вони починають дихати швидко й поверхнево. Це стосується й собак. У них, крім цього, посилюється випаровування вологи ще й з поверхні висунутого язика.
За частого дихання тварина сама створює потік повітря над вологою поверхнею на відміну від тварини, яка потіє. Ще одна відмінність частого дихання і потовиділення полягає в тому, що піт містить багато солей, а за сильного потовиділення втрата солей може бути такою, що викличе їх дефіцит. Водночас, за частого дихання тварини не втрачають солі (якщо тільки слина не капає з рота). Одна з переваг частого дихання — запобігання перегріванню мозку за раптового теплового стресу. Він може виникнути, наприклад, коли тварина спасається втечею від хижака, що призводить до сильного
підвищення метаболізму й відповідно температури тіла. У копитних тварин більша частина крові надходить до мозку по внутрішній сонній артерії, яка розділяється біля основи черепа на багато дрібних артерій, але перед тим, як увійти в мозок, вони знову сполучаються в загальний стовбур. Ці дрібні артерії проходять крізь великий венозний синус, через який тече кров від стінок носової порожнини. Вона охолоджена, особливо за частого дихання. Тож кров у дрібних артеріях охолоджується й тільки після цього досягає мозку. Так, наприклад, коли африканська газель біжить зі швидкістю 40 км/год протягом 5 хв, то температура її артеріальної крові з 39 °С піднімається до 44 °С, але температура мозку не досягає 41 % і залишається на безпечному рівні.
Зауважимо, що посилення м’язової роботи за частого дихання могло б призвести до продукування додаткової кількості тепла, але цього не відбувається в результаті того, що м’язова робота, а отже й теплопродукція, сильно зменшуються завдяки еластичним властивостям дихальної системи. Під час вдихання значна частина м’язової роботи втрачається на розтягування пружних елементів дихальної системи, які у процесі видихання самі собою стискуються. Подібно до інших еластичних систем, дихальна має власну частоту коливань. Для підтримання її коливань у власному ритмі (з резонансною частотою) необхідні мінімальні зусилля. Відповідно й вироблення тепла буде незначним.
Ще один спосіб підвищення випаровування вологи — змочування слиною та облизування. Воно притаманне багатьом сумчастим, у тому числі великим кенгуру, а також ряду гризунів. Цей спосіб не дуже ефективний і використовується, очевидно, як терміновий засіб тоді, коли температура тіла наближається до летальної.
Деякі плазуни, які потрапляють в умови перегрівання, починають дихати частіше відкритим ротом, що нагадує часте дихання у ссавців. Такий спосіб дихання збільшує випаровування вологи в дихальних шляхах. Крім цього, він, як у ссавців, може сприяти охолодженню мозку. Так, наприклад, у пустельної ящірки температуpa тіла не на багато холодніша 44,1 °С, але температура мозку залишається на рівні 42,3 °С. У цієї ящірки сонні артерії проходять дуже близько до поверхні глотки. Артеріальна кров у них охолоджується перед тим, як потрапити в мозок.
Птахи, на відміну від більшості ссавців, не мають потових залоз. Посилене випаровування вологи в них досягається завдяки частому диханню або швидким коливанням ротової порожнини й верхньої частини глотки — так зване гулярне дихання. Вони з метою охолодження можуть переходити на резонансну частоту дихання, як і ссавці. Очевидно, ці два шляхи підвищення випаровування вологи слугують для птахів ефективними способами охолодження.