Основы биохимии - Филиппович Ю. Б. 1999
Обмен белков
Регуляция белкового синтеза
Исходя из матричной гипотезы биосинтеза белка, Ф. Жакоб и Ж. Моно (1961) впервые предложили весьма многообещающую схему регуляции белкового синтеза. Следуя концепции один ген — одна мРНК — один белок, Ф. Жакоб и Ж. Моно связали узловой пункт регуляции белкового синтеза с ДНК, входящий в состав генетического аппарата клетки. По их мнению, в генетическом аппарате клетки существуют сообщества структурных генов, так называемых оперонов, каждый из которых ответствен за взаимосвязанный синтез ряда специфических белков. Деятельность оперона в качестве поставщика мРНК контролируется геном-оператором, который либо разрешает, либо запрещает запуск гомологической репликации серии мРНК на ДНК-матрице. В свою очередь, функция гена-оператора контролируется пространственно изолированным от него геном-регулятором, который продуцирует мРНК, необходимую для синтеза белка-репрессора. Именно белок-репрессор, будучи присоединен к гену-оператору, блокирует его функцию. Более того, сам белок-репрессор подвержен действию аллостерических эффекторов, которые, соединяясь с ним, так изменяют его третичную структуру, что либо стимулируют, либо ингибируют возникновение комплекса между репрессором и геном-оператором. В качестве аллостерических эффектов часто выступают субстраты (индуцированный синтез ферментов). Накапливаются данные об участии в контроле биосинтеза мРНК гормонов и ряда других соединений.
Сказанное иллюстрирует схема 4. В нее включены также информосомы, на уровне которых тоже осуществляется регуляция биосинтеза белков:
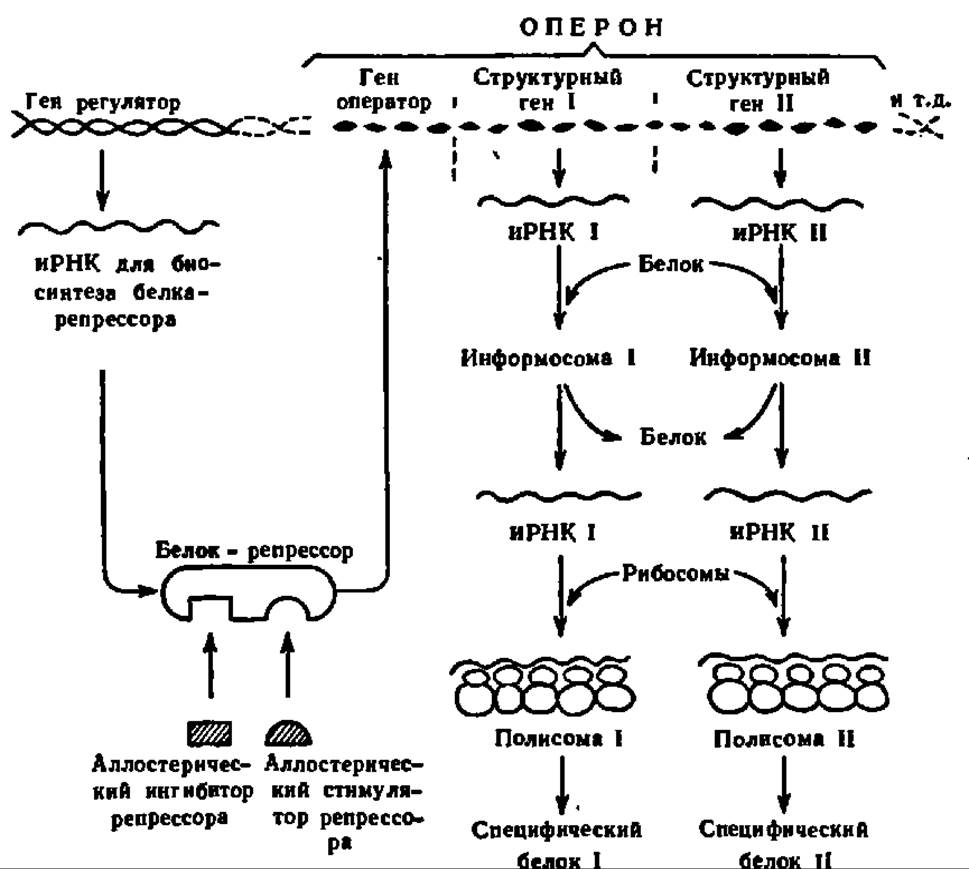
Схема 4. Регуляция биосинтеза белков
Данная схема отражает только часть тех факторов, которые принимают участие в регуляции биосинтеза белков. Эта регуляция осуществляется также на уровне метаболитов при активировании и переносе аминокислот; на уровне макромолекул при биосинтезе ДНК, различных видов РНК и рибосом; на уровне субклеточных структур (формирование полисом, роль белково-липидных мембран и т. п.), клетки (ядерноцитоплазменные взаимоотношения и др.), органа и организма (гормональная регуляция) и, наконец, на уровне среды (например, зависимость точности считывания кода белкового синтеза от температуры).
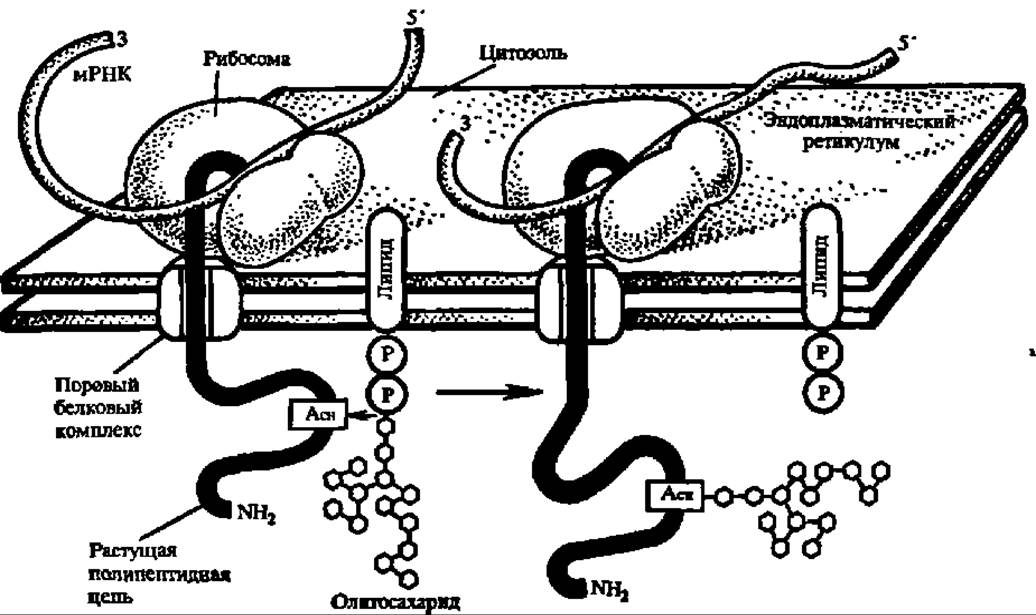
Рис. 102. Котрансляционное гликозилирование белка
Олигосахарид, связанный с липидом (долихолпирофосфатом — см. гл. IX). при участии гликозилтрансферазы переносится на остаток аспарагина синтезируемого а рибосоме белка, а результате чего возникает гликопротеин