ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
8. НУКЛЕОТИДЫ И НУКЛЕИНОВЫЕ КИСЛОТЫ
8.2. Структура нуклеиновых кислот
Открытие структуры ДНК Уотсоном и Криком в 1953 г. стало важнейшим научным событием, которое положило начало развитию ряда совершенно новых научных направлений и повлияло на уже существующие. В этом разделе мы основное внимание уделяем структуре ДНК, некоторым событиям, связанным с ее открытием, а также некоторым более современным уточнениям, дополняющим наши знания о ДНК. Здесь также будет рассмотрено строение молекулы РНК.
Как и при изучении строения белка (гл. 4), полезно описать строение нуклеиновых кислот, исходя из иерархии организации ее структуры (первичная, вторичная, третичная). Первичная структура нуклеиновых кислот образуется за счет ковалентных взаимодействий и представляет собой последовательность нуклеотидов. Любая регулярная стабильная форма, образованная из некоторых или всех типов нуклеотидов, может быть названа вторичной структурой нуклеиновой кислоты. Все виды пространственной организации, которые рассмотрены далее в данной главе, являются элементами вторичной структуры. Сложные комплексы, образующиеся при сборке больших хромосом из эукариотического хроматина или бактериальных нуклеоидов, рассматриваются в основном как третичные структуры; это будет обсуждаться в гл. 24.
ДНК — двойная спираль, хранящая генетическую информацию
ДНК была впервые выделена и описана Фридрихом Мишером в 1868 г. Он назвал фосфатсодержащее вещество «нуклеином». Но до 1940-х гг., до работ Освальда Т. Эйвери, Колина Маклеода и Маклина МакКарти не было исчерпывающих доказательств, что ДНК несет генетическую информацию. Эйвери с коллегами обнаружили, что ДНК, выделенная из болезнетворного штамма бактерии Streptococcus pneumoniae (пневмококк; вызывает у человека тяжелую болезнь), генетически трансформирует безопасный штамм этого организма в вирулентную форму. Был сделан вывод, что ДНК, выделенная из болезнетворного штамма, несет наследственный генетический признак вирулентности. В 1952 г. Эксперимент Альфреда Д. Херши и Марты Чэйз, в котором было изучено заражение бактериальной клетки вирусом (бактериофагом) с радиоактивно меченной ДНК или белком, ясно доказал, что именно ДНК, а не белок, несет наследственную информацию.
Еще один важный ключ к разгадке строения ДНК был получен в работе Эрвина Чаргаффа и его коллег в конце 1940-х гг. Они обнаружили, что четыре нуклеотидных основания ДНК встречаются в разных соотношениях в молекулах ДНК разных организмов, но содержание определенных пар оснований практически одинаково. Эти данные, собранные для огромного количества организмов, принадлежащих к разным видам, привели Чаргаффа к следующим выводам:
1. Содержание оснований в ДНК разных видов, как правило, различается.
2. Образцы ДНК, выделенные из разных тканей организмов одного вида, имеют одинаковый состав оснований.
3. Состав оснований ДНК конкретного вида не зависит от возраста, питания, окружающей среды.
4. Во всех клеточных ДНК вне зависимости от вида, к которому принадлежит организм, число остатков аденозина равно числу остатков тимидина (А = Т), а число остатков гуанозина равно числу остатков цитидина (G = С). Отсюда следует, что сумма пуриновых оснований равна сумме пиримидиновых оснований; т. е. А + G = Т + С.
Эти количественные отношения, называемые иногда «правилами Чаргаффа», были подтверждены многими исследователями. Эти правила помогли разгадать трехмерную структуру ДНК и пролили свет на то, как генетическая информация кодируется в ДНК и передается из поколения в поколение.
Розалинд Франклин и Морис Уилкинс использовали мощный метод рентгеноструктурного анализа (доп. 4-5) для исследования нитей ДНК. В начале 1950-х гг. они показали, что при дифракции рентгеновских лучей на ДНК образуется характерная дифракционная картина (рис. 8-12). Из таких фотографий следовало, что молекулы ДНК представляют собой периодическую спиральную структуру с двумя периодами вдоль длинной оси — один 3,4 А, другой — 34 А. После этого встала проблема подобрать такую трехмерную модель молекулы ДНК. чтобы она отвечала не только всем данным рентгеноструктурного анализа, но и правилу Чаргаффа о равенстве числа оснований в парах А = Т и G = С и другим известным химическим свойствам ДНК.

Рис. 8-12. Рентгенограмма ДНК. Пятна образуют крест в центре различимой спиральной структуры. Темные полосы по бокам образованы повторяющимися основаниями.

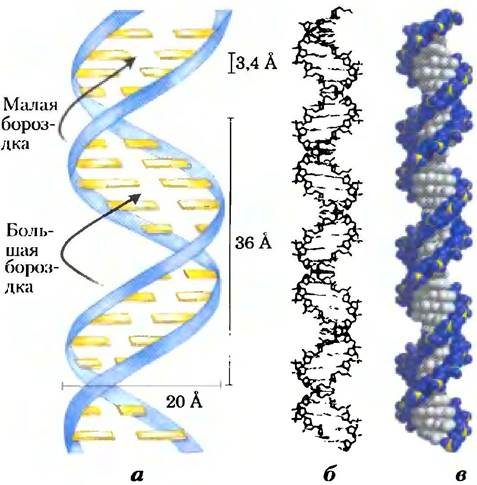
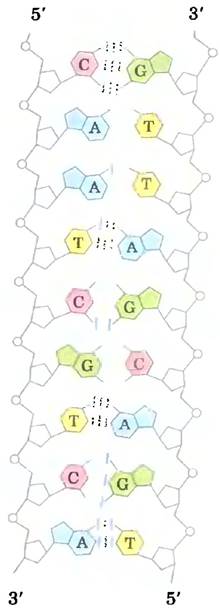
Джеймс Уотсон и Френсис Крик, проанализировав уже полученную информацию о ДНК, смогли понять, как она построена. В 1953 г. Уотсон и Крик представили трехмерную модель структуры ДНК, которая удовлетворяла всем известным данным. Она состояла из двух спиральных цепочек ДНК, закрученных вокруг одной и той же оси с образованием двойной правозакрученной спирали (см. доп. 4-1 для объяснения способов определения право- или левозакрученной спиральной структуры). Гидрофильные остовы чередующихся дезоксирибозных и фосфатных групп находятся снаружи двойной спирали и по соседству с водным окружением. Циклы фуранозы каждой дезоксирибозы находятся в С-2' эндо-конформации. Пуриновые и пиримидиновые основания обоих тяжей уложены очень близко друг к другу внутри двойной спирали. Это гидрофобные, почти плоские структуры, перпендикулярные главной оси ДНК. При таком соединении на поверхности парной структуры образуются большая бороздка и малая бороздки (рис. 8-13). Каждое нуклеотидное основание одной цепи образует пару с основанием другой цепи в одной плоскости. Уотсон и Крик показали, что связанные водородными связями пары оснований С с G и А с Т, показанные на рис. 8-11, наилучшим образом согласуются с их структурой и объясняют правило Чаргаффа о том, что в любой молекуле ДНК G = С и А = Т. Надо подчеркнуть, что между G и С могут образоваться три водородные связи, что обозначается как G = С, а между A и Т могут образоваться только две водородные связи, что обозначается как A = Т. Это одна из причин, что разделение спаренных цепей ДНК затруднено, если число парбольше, чем А = Т. Образование пар между другими основаниями в той или иной степени дестабилизирует структуру двойной спирали.
Рис. 8-13. Модель структуры ДНК, предложенная Уотсоном и Криком. Исходная модель, предложенная Уотсоном и Криком, содержала 10 пар оснований, имела длину 34 А (или 3,4 нм) на один оборот спирали; последующие измерения показали, что на один оборот приходится 10,5 пар оснований, и он имеет длину 36 А (или 3,6 нм), а) Схематическое изображение, показывающее размеры спирали. б) Схематическая модель в виде палочек, изображающая остов ДНК и стопку оснований, в) Объемная модель.
Уотсон и Крик, конструируя свою модель, должны были принять одну из двух гипотез о расположении цепей двойной спирали ДНК — параллельны или антипараллельны эти цепи, т. е. направлены ли их 5',3'-фосфодиэфирные связи в одну или в разные стороны. Модель с антипараллельной ориентацией цепей была более убедительной, и в более поздних работах с ДНК-полимеразой (гл. 25) были получены экспериментальные доказательства того, что цепи действительно антипараллельны, а в дальнейшем это полностью подтвердилось рентгеноструктурным анализом.
Для учета периодичности, обнаруженной при рентгеноструктурном анализе цепей ДНК, Уотсон и Крик создали молекулярную модель, в которой соседние основания одной цепи внутри двойной спирали находились бы на расстоянии 3,4 А друг от друга; и вся эта структура повторялась через 34 А, совершая полный оборот двойной спирали за 10 пар оснований. В водном растворе структура ДНК слегка отличается от той, что была получена в кристалле, и имеет 10,5 пар оснований на каждый полный оборот спирали (рис. 8-13).
Как показано на рис. 8-14, две антипараллельные полинуклеотидные цепи ДНК не идентичны ни по последовательности пар оснований, ни по составу. Но они комплементарныдруг другу. Если в одной цепи встречается аденин, в другой на этом месте расположен тимин; аналогично, гуанину в одной цепи соответствует цитозин в другой.
Рис. 8-14. Комплементарность цепей в двойной спирали ДНК. Комплементарность антипараллельных цепей ДНК следует из правил образования пар, предположенных Уотсоном и Криком. Комплементарные антипараллельные цепи отличаются друг от друга по составу: если левая цепь имеет формулу А3Т2G1С3, то правая — А2Т3G3С1. Они также различаются и по последовательности, если читать ее в направлении 5' —> 3’. Обратите внимание, что в двойной спирали число парных оснований совпадает, а именно А = Т и G = С.
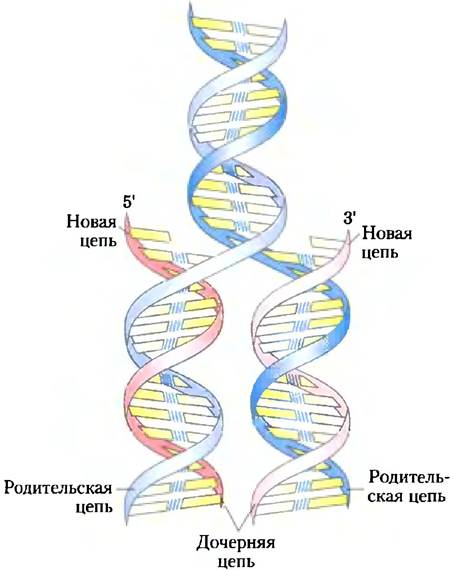
ДНК представляет собой двойную спираль (дуплекс), которая удерживается за счет двух типов сил, описанных ранее: водородных связей между парами оснований (рис. 8-11) и стекинговых взаимодействий между параллельными плоскостями соседних оснований. Комплементарность между цепями ДНК обеспечивается за счет образования водородных связей между парами оснований. Главный вклад в стабильность двойной спирали вносят стекинговые взаимодействия между соседними основаниями одной цепи, которые менее специфичны в отношении типов оснований. Важные особенности модели двойной спирали ДНК подтверждаются большим количеством химических и биологических данных. Более того, эта модель также объясняет механизм, по которому возможен перенос генетической информации. Существенной чертой модели является комплементарность двух цепей ДНК. Уотсон и Крик сумели выявить эту особенность логически, еще до того, как были получены подтверждающие данные, что такая структура может быть реплицирована (1) разделением двух цепей и (2) достройкой комплементарной цепи каждой спирали. Так как нуклеотиды каждой новой цепи подбираются в последовательности, удовлетворяющей правилу спаривания оснований, каждая существующая цепь играет роль образца, по которому синтезируется комплементарная цепь (рис. 8-15). В дальнейшем эти объяснения были подтверждены экспериментально, что вызвало революционные изменения нашего понимания биологической наследственности.
Рис. 8-15. Репликация цепей ДНК, предположенная Уотсоном и Криком. Уже существующие («родительские») цепи ДНК расходятся, и каждая из них служит шаблоном для синтеза комплементарной («дочерней») цепи (красная).
Пример 8-1. Пары оснований в ДНК
В образцах ДНК, выделенных из двух неизвестных видов бактерий (X и Y), аденин составляет соответственно 32 и 17% от общего состава оснований. Каково относительное содержание гуанина, тимина и цитозина в двух образцах ДНК? Какие допущения вам пришлось сделать, чтобы ответить на этот вопрос? Один из видов бактерий был выделен из горячего источника с температурой 64 °С. Какой из двух видов с наибольшей вероятностью выделен из горячего источника и почему?
Решение. В любой двойной спирали ДНК А = Т и G = С. ДНК из бактерии X содержит 32% А, следовательно, она содержит 32% Т. В сумме это дает 64%, следовательно, суммарное содержание пар составляет оставшиеся 36%: 18% приходится на в и 18% на С. В образце из бактерии Y содержится 17% А, следовательно, 17% Т, в сумме 34%. Оставшиеся 66% распределяются поровну между G и С (по 33%). Данный расчет сделан на основании предположения, что обе молекулы ДНК являются двунитевыми.
Чем выше содержание G + С пар в молекуле ДНК, тем выше температура плавления ДНК. Более вероятно, что из горячего источника была выделена бактерия У, в которой содержится больше в G+С пар. Ее ДНК имеет более высокую температуру плавления и, следовательно, обладает большей стабильностью при температуре воды в горячем источнике.
ДНК может иметь разные пространственные конфигурации
ДНК — мобильная молекула. Возможно вращение вокруг некоторых связей сахарофосфатного (фосфодезоксирибозного) скелета, и из-за теплового движения может происходить сгибание, растягивание и распаривание (плавление) цепей. В клеточной ДНК было найдено довольно большое количество типов структур, производных модели ДНК Уотсона и Крика, причем многие из них могут играть важную роль в метаболизме ДНК. Эти структурные вариации обычно не влияют на ключевые свойства ДНК, определенные Уотсоном и Криком: на комнлементарность цепей и их антипараллельность и на выполнение требования А = Т и G = С.
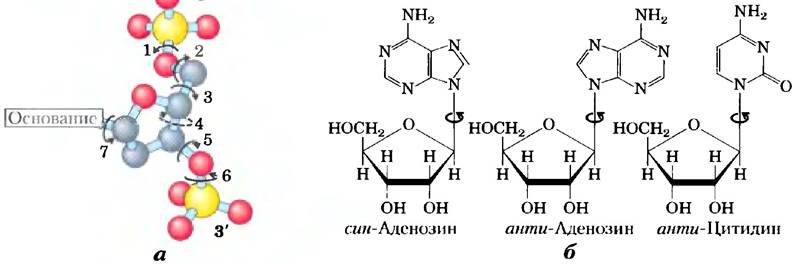
Изменение пространственной структуры ДНК связано с тремя возможностями: разные конформации дезоксирибозы, вращение вокруг смежных связей, из которых состоит фосфодезоксирибозный остов (рис. 8-16, а), и свободное вращение вокруг С-1'-N-гликозидной связи (рис. 8-16, б). Из-за стерических ограничений пурины и пуриновые нуклеотиды могут находиться только в двух стабильных конформациях по отношению к дезоксирибозе, их называют син- и анти- (рис. 8-16, б). Пиримидины могут существовать только в одной анти-конформации из-за стерического взаимодействия между сахаром и карбонильным кислородом при атоме С-2 пиримидинового цикла.
Рис. 8-16. Вариации в структуре ДНК. а) Конформация нуклеотидов в ДНК зависит от значений углов вращения вокруг семи разных связей. Вокруг шести из них возможно свободное вращение. Из-за ограниченного вращения вокруг четвертой связи меняется конформация пентозного цикла: один из атомов в пятичленном фуранозном кольце находится вне плоскости, образованной остальными четырьмя. В результате получается эндо- или экзо-конформация, в зависимости оттого, находится ли атом вне плоскости с той же стороны от нее, что и С-5' атом, или с противоположной (рис. 8-3, б). б) Для пуриновых нуклеотидов по отношению к рибозе возможно только две стерических конформации — анти- или син-. Пиримидины встречаются обычно в онто-конформации.
Пространственная организация ДНК, предложенная Уотсоном и Криком, также называется В-формой ДНК (В-ДНК). Образование В-формы наиболее вероятно для случайной последовательности молекулы ДНК при физиологических условиях, таким образом, В-ДНК служит стандартом, на который ссылаются при исследовании любых свойств ДНК. Два других варианта, хорошо охарактеризованных в кристалле, называются А- и Z-формами. Эти три конформации ДНК показаны на рис. 8-17 с кратким перечислением их основных свойств. А-форма образуется в средах с низким содержанием воды. А-ДНК представляет собой правозакрученную двойную спираль, на один оборот которой приходится 11 пар оснований (в отличие от 10,5 в В-ДНК). Плоскость пар оснований в А-ДНК находится под углом приблизительно 20° к оси спирали. В результате этих структурных изменений углубляется большая бороздка, в то время как малая бороздка становится почти незаметной. Реагенты, используемые при кристаллизации ДНК, как правило, дегидрируют ее, поэтому большинство коротких молекул ДНК кристаллизуются именно в А-форме.
Рис. 8-17. Сравнение А-, В- и Z-форм ДНК. Каждая молекула, показанная здесь, состоит из 36 пар оснований. Основания показаны серым, атомы фосфора желтым, а рибоза и атомы кислорода фосфатных групп — синим. В последующих главах синий цвет используется при изображении цепей ДНК. В таблице суммированы свойства трех форм ДНК.
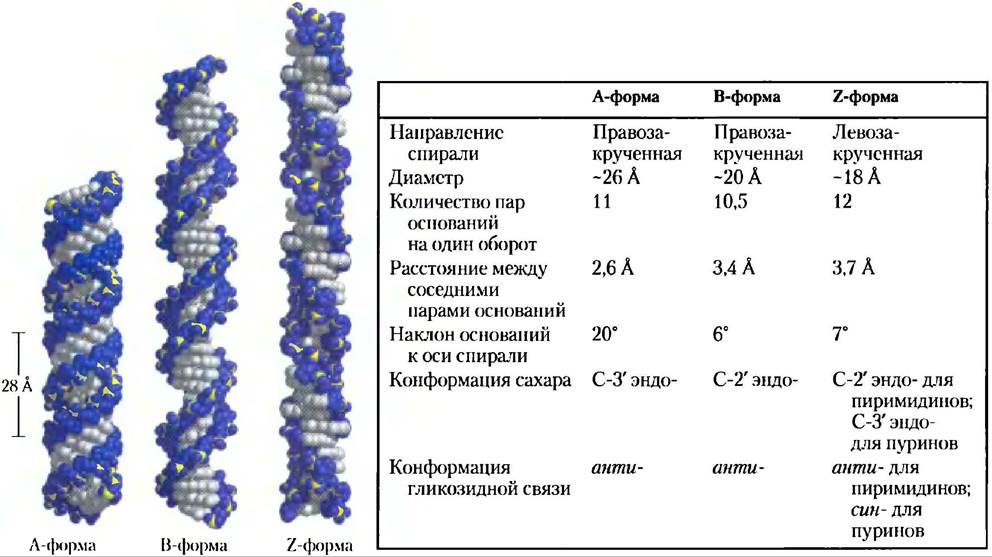
Z-форма ДНК очень сильно отличается от В-структуры; самое главное отличие заключается в том, что это — левозакрученная спираль. На один ее оборот приходится 12 оснований, сама молекула более узкая и удлиненная. Остов ДНК принимает зигзагообразную форму. Левозакрученную спираль образует чаще всего вполне определенная последовательность нуклеотидов. Наиболее известные примеры таких последовательностей — это чередующиеся пиримидиновые и пуриновые основания, особенно чередующиеся остатки С и G или 5-метил-С и G. При образовании левозакрученной спирали в Z-ДНК пуриновые остатки принимают син-конформацию и чередуются с пиримидинами в анти-конформации. Большая бороздка в Z-ДНК едва заметна, а малая бороздка узкая и глубокая.
Встречается ли А-ДНК в клетках — неизвестно, но по некоторым данным, короткие фрагменты Z-ДНК могут быть и у прокариот, и у эукариот. Эти фрагменты Z-ДНК могут играть роль (пока неизвестно какую) в регуляции экспрессии некоторых генов или в генетической рекомбинации.
Определенные последовательности ДНК могут иметь необычное пространственное строение
В больших хромосомах был найден ряд других вариантов трехмерной организации ДНК, форма которых зависит от последовательности. Это может влиять на функцию и метаболизм сегментов ДНК, которые находятся в непосредственной близости друг от друга. Например, изгибы встречаются в спиралях ДНК везде, где расположены подряд четыре или более остатков аденозина. Шесть аденозинов подряд образуют изгиб под углом -18°. Такая кривизна, образуемая этой или другой последовательностью, может играть важную роль при связывании ДНК с некоторыми белками.
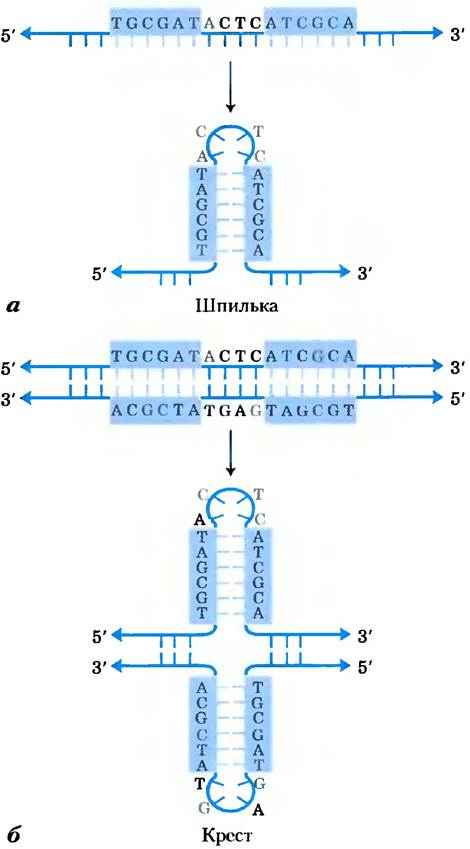
Достаточно распространенный тип последовательности ДНК — это так называемый палиндром. Палиндром — это слово, фраза или предложение, которое звучит одинаково при чтении слева направо и справа налево, например, потоп, мед ждем. Этот термин применяется для обозначения участков ДНК с инвертированными повторамипоследовательностей оснований, которые имеют симметрию второго порядка для двух цепей ДНК (рис. 8-18). Такая последовательность комплементарна самой себе, и поэтому может образовывать структуры типа шпильки или креста (две симметричные шпильки) (рис. 8-19). Если инвертированные повторы встречаются в одной цепи ДНК, такие последовательности называются зеркальными повторами. Зеркальные повторы не являются комплементарными последовательностями для самих себя и не способны образовывать шпильки и крестообразные структуры. Последовательности такого типа найдены практически во всех больших молекулах ДНК и могут включать в себя от нескольких оснований до нескольких тысяч оснований. Частота, с которой палиндромы встречаются в виде крестообразных структур в клетке, неизвестна, хотя в некоторых случаях такие формы организации ДНК были найдены in vivo у Escherichia coli. Отдельные цепи самокомплементарных последовательностей ДНК (или РНК) в растворе выделяются в виде сложных структур, содержащих большое количество шпилек.
Рис. 8-18. Палиндромы и зеркальные повторы. Палиндромами называются такие последовательности в двойной спирали нуклеиновых кислот, которые обладают симметрией второго порядка. При повороте на угол 180° вокруг горизонтальной оси, а затем на угол 180° вокруг вертикальной оси такая структура накладывается сама на себя, что показано цветными стрелками. Зеркальные повторы — симметричные повторы в каждой цепи. Для того чтобы наложить эту структуру саму на себя, необходимо всего лишь один раз повернуть последовательность вокруг вертикальной оси на 180°.
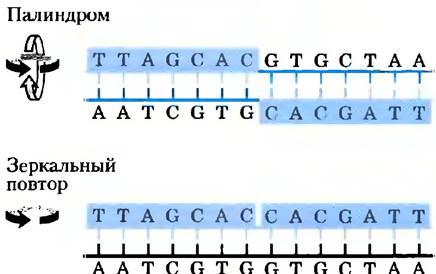
Рис. 8-19. Шпильки и кресты. Палиндромные последовательности ДНК (или РНК) могут по-разному сворачиваться с образованием пар между основаниями внутри одной цепи, а) Когда в образовании структуры участвует только одна последовательность ДНК (или РНК), полученная форма называется шпилькой. б) Крестом называется вторичная структура из палиндромных последовательностей, образованная при участии двух нитей ДНК. Синим закрашены асимметричные участки ДНК, которые комплементарны последовательностям в той же или в другой цепи.
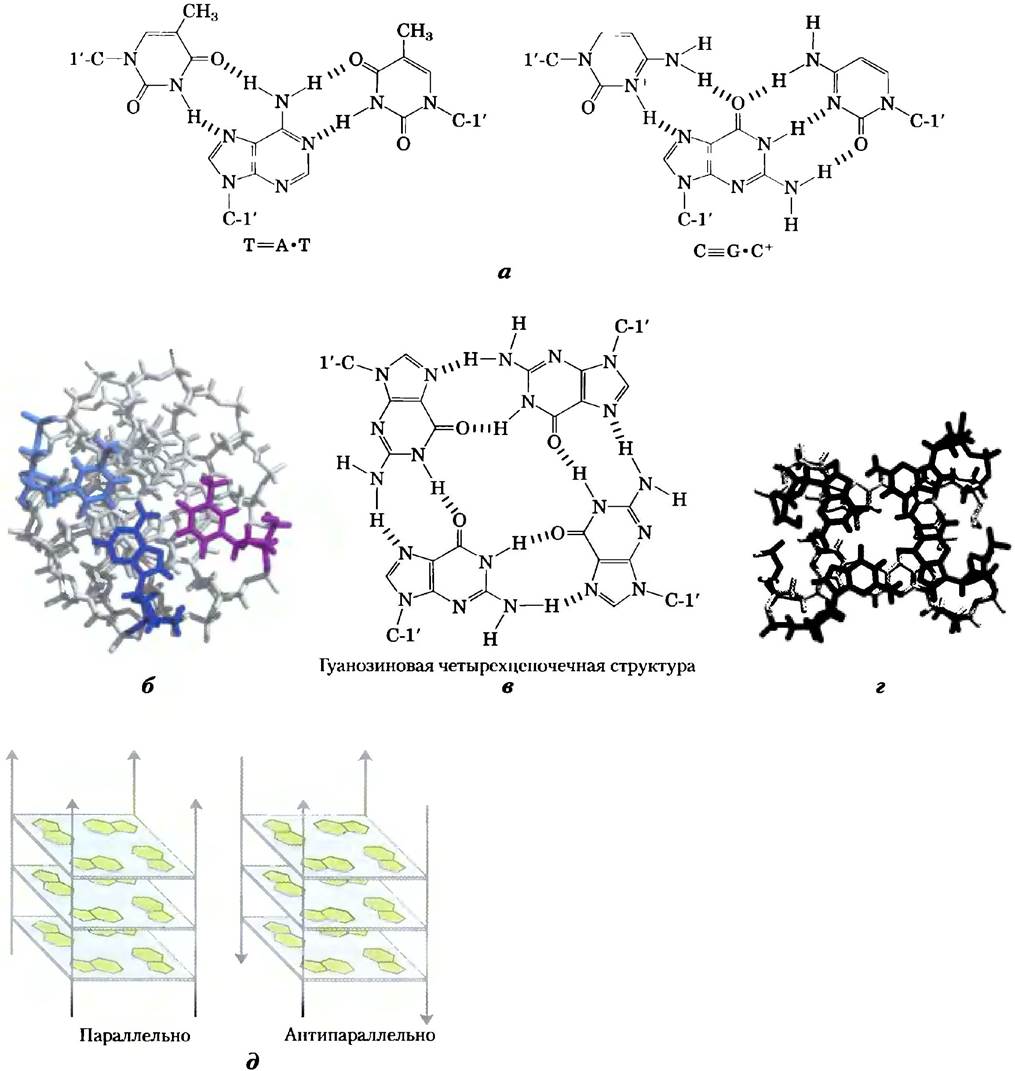
Некоторые необычные структуры ДНК образуются из трех или более цепей ДНК. Нуклеотиды, участвующие в образовании уотсон- криковских пар (рис. 8-11), могут иметь ряд дополнительных водородных связей, особенно с функциональными группами, расположенными в большой бороздке. Например, остаток цитидина (протонированный) может связываться с остатком гуанозина, участвующим в образовании нуклеотидной пары G=C, а тимидин — с аденозином А=Т пары (рис. 8-20). Атомы N7, О6 и N6 пуринов могут участвовать в образовании водородных связей в триплексе ДНК; эти атомы иногда называются хугстиновскими положениями, а не-уотсон-криковские пары — хугстиновскими парами, по имени Карста Хугстина, который в 1963 г. первым указал на возможность существования такого необычного способа образования пар. Благодаря этим нетипичным водородным связям формируется трехцепочечная ДНК (триплекс). Трехцепочечные фрагменты, показанные на рис. 8-20 (а, б), наиболее стабильны при низких значениях pH, так как для образования триплета G=C • С+ необходимо протонирование цитозина. В трехцепочечной структуре рКа цитозина >7,5, что отличается от обычного значения 4,2. Эти структуры также более охотно формируются в длинных последовательностях, содержащих только пиримидины или пурины в одной из цепей. Некоторые трехцепочечные структуры ДНК содержат две пиримидиновые цепи и одну пуриновую; другие состоят из двух пуриновых и одной пиримидиновой.
Четыре цепи ДНК также могут связываться друг с другом, образуя четырехцепочечную структуру (тетраплекс или квадруплекс), но это происходит только между цепочками ДНК с высоким содержанием гуанозиновых остатков (рис. 8-20, в, г). Гуанозиновая четырехцепочечная структура, или G-тетраплекс, довольно стабильна в широком диапазоне условий. Ориентация цепей в тетраплексе может быть различной, как показано на рис. 8-20, д.
Рис. 8-20. Строение ДНК, состоящей из трех или четырех цепей, а) Схема соединения оснований в одной из хорошо изученных форм трехцепочечной ДНК. Хугстиновские пары на каждом изображении отмечены красным, б) Тройная спираль ДНК, состоящая из двух пиримидиновых цепей (поли(Т)) и одной пуриновой (поли(А)) (взято из PDP ID 1ВСЕ). Темно-синяя и голубая цепи направлены антипараллельно и образуют типичные уотсон- криковские пары. Третья (полностью пиримидиновая) цепь (сиреневый цвет) направлена параллельно пуриновой цепи и связана с другими цепями водородными связями, которые не участвуют в образовании уотсон-криковских пар. Вид на трехцепочечную структуру снизу, показана стопка из пяти плоскостей оснований. Ближайший триплет изображен разными цветами, в) Фрагмент четырехцепочечной структуры из гуанозинов. г) Два последовательных G-тетраплекса (взято из PDB ID 1QDG), вид снизу, ближайший тетраплекс изображен разными цветами, д) Возможные варианты ориентации цепей, содержащих G-тетраплексы.
В ДНК живой клетки сайты узнавания многих специфических ДНК-связывающих белков (гл. 28) представлены палиндромами, а полипиримидины и полипурины, которые могут образовывать тройные спирали или даже Н-ДНК, найдены в регионах, участвующих в регуляции или экспрессии некоторых эукариотических генов. В принципе, искусственные цепочки ДНК, синтезированные таким образом, чтобы связываться с этими последовательностями с образованием трехцепочечной ДНК, могут нарушить экспрессию генов. Это важно отметить в связи с большим коммерческим интересом к возможности регуляции экспрессии генов для решения различных задач в области медицины и сельского хозяйства.
Матричные РНК содержат информацию о полипептидных цепях
После краткого рассмотрения структуры ДНК обратимся к обсуждению той генетической информации, которую она содержит. РНК, второй важный тип нуклеиновых кислот в клетке, выполняют множество функций. В процессе экспрессии генов РНК участвуют в качестве посредников, использующих информацию, закодированную в ДНК, для формирования аминокислотной последовательности функционального белка.
Учитывая то, что ДНК эукариот находится в ядре, а синтез белка происходит в цитоплазме на рибосомах, генетическую информацию из ядра в цитоплазму должны переносить не ДНК, а другие молекулы. С начата 1950-х гг. РНК рассматривалась в качестве наиболее вероятного кандидата: эти молекулы были найдены и в ядре, и в цитоплазме, а увеличение синтеза белка сопровождалось увеличением содержания РНК в цитоплазме и увеличением скорости се оборота. Эти и другие сведения привели некоторых исследователей к мысли о том, что РНК переносит генетическую информацию от ДНК к аппарату для биосинтеза белка — рибосоме. В 1961 г. Франсуа Жакоб и Жак Моно предложили целостную (и по существу верную) картину этого процесса и термин матричная РНК, мРНК (messenger RNA) для той части клеточной РНК, которая переносит генетическую информацию от ДНК к рибосомам, где становится шаблоном, по которому собирается специфическая последовательность аминокислот. Хотя длина мРНК разных генов может сильно варьировать, для конкретного гена она имеет определенный размер. Процесс образования мРНК по матрице ДНК называется транскрипцией.
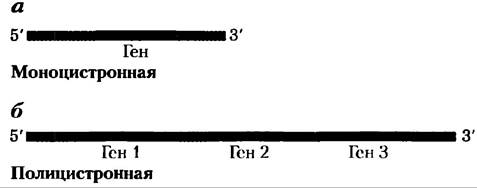
У бактерий и архей одна молекула мРНК может кодировать одну или несколько полипептидных цепей. Если в мРНК закодирован только один полипептид, то она называется моноцистронной; если два или более разных полипептидов — полицистронной. У эукариот большинство мРНК моноцистронные. (Для простоты цистрон здесь называется геном. Термин сам по себе имеет исторические корни в генетике, и его формальное определение выходит за рамки данного изложения.) Минимальная длина мРНК определяется длиной полипептидной цепи, которую она кодирует. Например, для синтеза полипептидной цепи из 100 аминокислотных остатков необходима кодирующая последовательность РНК длиной, по крайней мере, 300 нуклеотидов, гак как каждая аминокислота кодируется триплетом нуклеотидов (эта и другие детали синтеза белка рассматриваются в гл. 27). Однако длина мРНК, синтезируемой по матрице ДНК, всегда немного больше, чем необходимо просто для кодирования полипептидной последовательности (или последовательностей). Дополнительная, некодирующая последовательность РНК содержит участки, которые регулируют белковый синтез. На рис. 8-21 изображена общая структура бактериальной мРНК.
Рис. 8-21. Бактериальная мРНК. Здесь представлена схематическая диаграмма, на которой показаны а) моноцистронная и б) полицистронная молекулы мРНК бактерий. Красные сегменты представляют участок РНК, в котором закодирован отдельный ген; серые сегменты — некодирующие участки РНК. В полицистронном транскрипте некодирующие участки РНК разделяют три гена.
Многие молекулы РНК имеют очень сложную трехмерную структуру
Матричная РНК — это только один из нескольких классов клеточной РНК. Транспортная РНК служит вспомогательной молекулой при синтезе белка; она ковалентно связана с аминокислотой и при этом может комплементарно соединяться с матричной РНК таким образом, чтобы аминокислота встраивалась в растущую цепь в правильном порядке. Рибосомная РНК функционирует в качестве одного из компонентов рибосом. Существует также много разных типов РНК со специализированными функциями, в том числе такие, которые обладают ферментативной активностью (рибозимы).
Все типы РНК детально рассматриваются в гл. 26. РНК выполняют много разных функций, часто несколько сразу, и это отражает тот факт, что их пространственное строение более разнообразно, чем у молекулы ДНК.
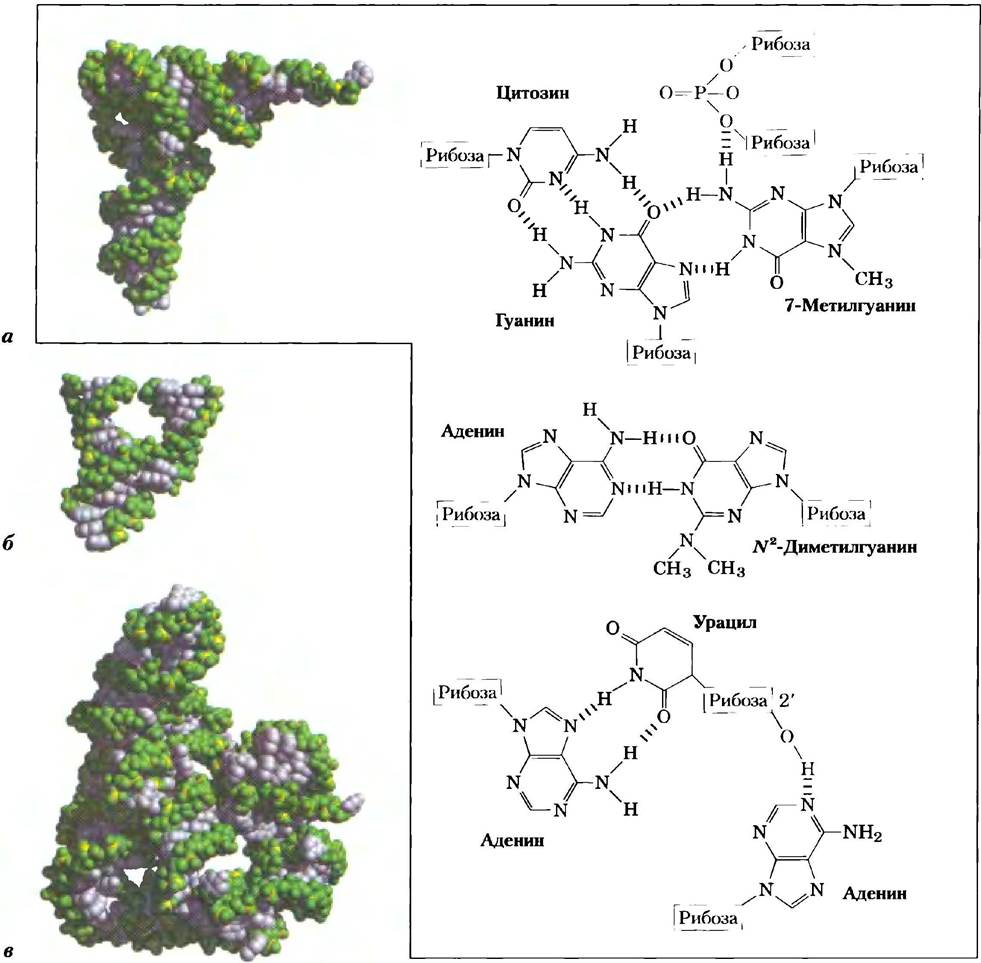
Продуктом транскрипции ДНК всегда является одноцепочечная РНК. Единственная нить принимает, как правило, правозакрученную конформацию благодаря стекинговым взаимодействиям между основаниями (рис. 8-22), причем они намного сильнее между двумя пуринами, чем между пурином и пиримидином или между двумя пиримидинами. Пурин-пуриновое взаимодействие настолько сильно, что пиримидин, разделяющий два пурина, может быть вытеснен из области между пуринами для обеспечения контакта между пуринами. Любая комплементарная самой себе последовательность формирует более сложные структуры. РНК может образовывать пары оснований либо с ДНК, либо с РНК. Пары практически такие же, как и в ДНК: G связывается с С, А с U (или с редко встречающимся остатком Т в некоторых РНК). Единственное различие состоит в том, что возможно образование пары между остатками G и U — это необычно для ДНК, но довольно распространено в РНК (см. рис. 8-24). Спаренные цепочки в двуцепочечных РНК или РНК-ДНК антипараллельны, как и в ДНК.
Рис. 8-22. Типичный правозакрученный фрагмент одноцепочечной РНК. Основания показаны серым, атомы фосфора желтым, остатки рибозы и атомы кислорода фосфатных групп — зеленым цветом. В последующих главах зеленый цвет используется для изображения цепей РНК, а синий — для обозначения цепей ДНК.
РНК не имеет регулярной вторичной структуры, которая могла бы послужить стандартом, как двойная спираль ДНК. Трехмерные структуры многих РНК, как и белков, сложны и уникальны. Слабые взаимодействия, особенно стекинговые, играют огромную роль в стабилизации структур РНК, так же, как и в ДНК. В участках, где присутствуют комплементарные последовательности, преобладающая двухнитевая структура — это А-форма правозакрученной двойной спирали. Спирали Z-формы были получены в лабораторных условиях (при высокой температуре и очень высокой солености). В-форма РНК не была обнаружена. Разрывы в регулярной спирали А-формы, которые появляются из-за пропущенных или неподходящих оснований в одной или обеих нитях, встречаются довольно часто, и в результате образуются выступы или внутренние петли (рис. 8-23). Расположенные поблизости комплементарные участки РНК образуют шпильки. В структуре РНК заложены широкие возможности для образования двойных спиралей (рис. 8-24), поэтому шпильки — это наиболее часто встречающаяся вторичная структура. На конце шпилек РНК часто обнаруживаются специфические короткие последовательности оснований (такие как UUCG); известно, что они могут образовывать прочные и стабильные петли. Такие последовательности могут быть отправными пунктами при сворачивании молекулы РНК в трехмерную структуру. Важный дополнительный вклад в стабилизацию молекулы вносят водородные связи, которые не участвуют в образовании стандартных уотсон-криковских пар. Например, водородные связи может образовывать 2'-гидроксильная группа рибозы. Некоторые из этих особенностей хорошо изучены в структуре фенилаланиновой транспортной РНК дрожжей (отвечает за включение остатка Рhе в полипептиды) и в двух РНК-ферментах, или рибозимах, чьи функции, как и белковых ферментов, зависят от их трехмерной структуры (рис. 8-25).
Рис. 8-23. Вторичная структура молекул РНК. а) Выступы, внутренние петли и шпильки. б) Как показано здесь для структуры типа «шпилька», участки двойной спирали РНК обычно находятся в А-форме.
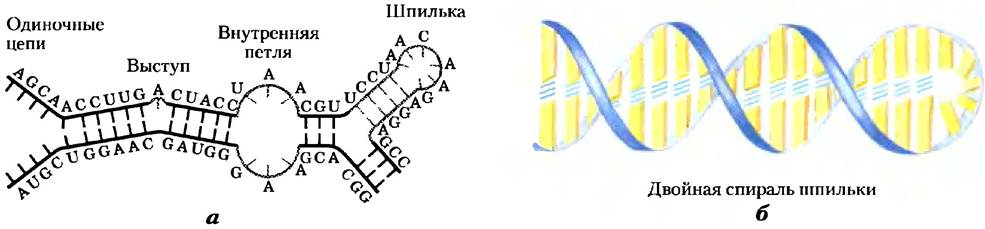
Рис. 8-24. Структуры двойных спиралей в РНК. Здесь показана возможная вторичная структура РНК для Ml-компонента фермента РНКазы Р из Е. coli; в этой молекуле РНК большое количество шпилек. РНКаза Р, которая также содержит и белок (здесь не показан), участвует в созревании молекул транспортных РНК (рис. 26-27). Две квадратные скобки выделяют две другие комплементарные последовательности, которые могут связываться при образовании пространственной структуры. Синие точки показывают образование не уотсон-криковской пары оснований G = U (вставка). Обратите внимание на то, что образование пары G = C возможно только при сворачивании уже синтезированных цепей РНК или при отжиге их друг на друге. Не существует такой РНК-полимеразы (фермент, который синтезирует молекулы РНК по матрице ДНК), которая в процессе синтеза новой молекулы РНК вставляет U напротив G или наоборот.
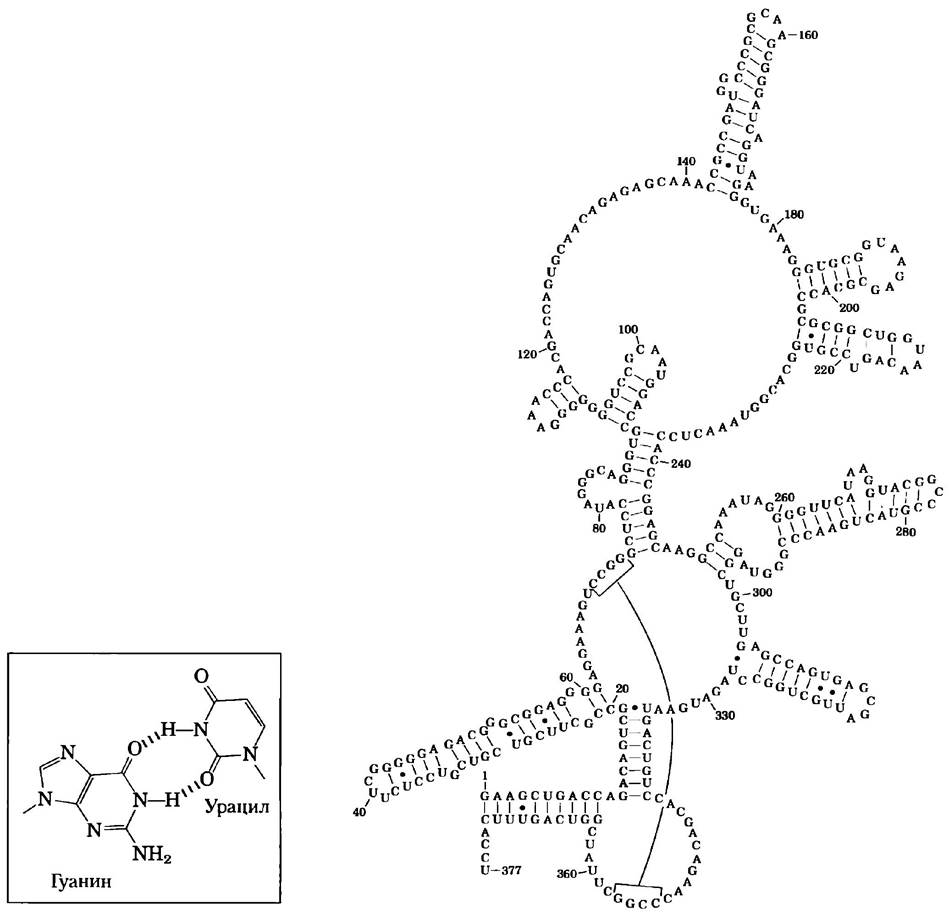
Рис. 8-25. Трехмерная структура РНК. а) Пространственное строение фенилаланиновой тРНК дрожжей (PDB ID 1TRA). Показаны некоторые необычные пары оснований, найденные в этой тРНК. Обратите внимание на водородные связи (показаны красным) между кислородом рибозы, участвующим в образовании фосфодиэфирной связи, и 2'-гидроксильной группой другой рибозы. б) Рибозим, выделенный из определенного вируса растений, вторичная структура которого похожа на головку молотка (взято из PDB ID 1MME). Рибозимы, или РНК-ферменты, катализируют разнообразные реакции, в первую очередь реакции метаболизма РНК и синтеза белков. Сложная трехмерная организация этих РНК отражает их участие в катализе, как это было описано для белков в гл. б. в) Сегмент мРНК, считающийся интроном (некодирующая последовательность), выделенный из реснитчатого простейшего Tetrahymena thermophila (взято из PDB ID 1GRZ). Этот интрон (рибозим) катализирует вырезание самого себя из участка цепи РНК, расположенного между двумя экзонами (кодирующая последовательность) (описано в гл. 26).
Анализ строения РНК и выявление взаимосвязи между структурой и функцией — новая исследовательская задача, где имеются те же сложности, что и при изучении пространственной организации белка. Важность понимания строения РНК растет по мере выявления функций молекул РНК.
Краткое содержание раздела 8.2 Структура нуклеиновых кислот
■ Большое количество разнообразных данных свидетельствует в пользу того, что ДНК служит носителем генетической информации. В частности, эксперимент Эйвери, Маклсода и МакКарти показал, что ДНК, выделенная из одного бактериального штамма, может попадать в клетки другого штамма и трансформировать их в клетки первого штамма, наделяя некоторыми наследственными характеристиками донора. Эксперимент Херши и Чейз показал, что ДНК бактериального вируса, а не его оболочка, переносит в клетку хозяина генетическую информацию для репликации вируса.
■ Обобщив множество опубликованных данных, Уотсон и Крик постулировали, что природная ДНК состоит из двух антипараллельных цепей, находящихся в форме правозакрученной спирали. Между комплементарными парами оснований, А = Т и G = С, образуются водородные связи внутри спирали. Пары оснований находятся в плоскости, перпендикулярной длинной оси двойной спирали, расстояние между соседними основаниями составляет 3,4 А, на один оборот спирали приходится 10,5 пар оснований.
■ ДНК может существовать в нескольких формах, которые отличаются друг от друга пространственным строением. Два варианта уотсон-криковской формы, или В-ДНК, — это А-ДНК и Z-ДНК. Некоторые структурные варианты зависят от последовательности нуклеотидов, в молекуле ДНК образуются изгибы. Цепи ДНК с определенными последовательностями могут образовывать шпильки, крестообразные структуры, а также участки трехцепочечной и четырехцепочечной ДНК.
■ Матричная РНК переносит генетическую информацию от ДНК к рибосомам для синтеза белков. Транспортная РНК и рибосомная РНК также участвуют в белковом синтезе. Пространственная структура РНК довольно сложная; отдельные цени могут сворачиваться в шпильки, принимать форму двойной спирали или образовывать сложные петли.