ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
Структура этого просто обязана существовать.
Джеймс Уотсон, Двойная спираль, 1968
8. НУКЛЕОТИДЫ И НУКЛЕИНОВЫЕ КИСЛОТЫ
В клеточном метаболизме нуклеотиды выполняют множество функций. Они переносят энергию для биохимических превращений, играют роль важных химических посредников (вторичных мессенджеров) в ответе клеток на действие гормонов и других внеклеточных стимулов, входят как структурные элементы в состав ряда кофакторов ферментов и метаболических интермедиатов. Наконец, из нуклеотидов построены нуклеиновые кислоты: дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК), в которых хранится на молекулярном уровне генетическая информация. Структура каждого белка и, в конечном итоге, всех биомолекул и компонентов клетки определяется данными, закодированными в последовательности нуклеотидов молекул нуклеиновых кислот. Способность хранить и передавать генетическую информацию от одного поколения другому входит в число основных условий жизни.
Эта глава посвящена обзору химических свойств нуклеотидов и нуклеиновых кислот, обнаруженных в большинстве типов клеток; детальное описание функций нуклеиновых кислот дано в части III.
8.1. Основные понятия
Нуклеотиды — блоки, из которых построены нуклеиновые кислоты.Аминокислотная последовательность каждого белка клетки и нуклеотидная последовательность каждой молекулы РНК определены последовательностью нуклеотидов соответствующего участка клеточной ДНК. Участок молекулы ДНК, содержащий необходимую информацию для синтеза биологического продукта с определенной функцией, будь это белок или РНК, называется ген. Обычно в клетке содержится много тысяч генов, и неудивительно, что молекулы ДНК очень велики. Хранение и передача биологической информации — единственная известная функция молекул ДНК.
У РНК более широкий набор функций; в клетке существует несколько разновидностей этих молекул. Рибосомная РНК (рРНК) входит в состав рибосом — сложных комплексов, где происходит синтез белков. Матричная РНК (мРНК) служит посредником, переносящим генетическую информацию от одного или нескольких генов к рибосоме, в которой могут синтезироваться соответствующие белки. Транспортная РНК (тРНК) точно преобразует информацию мРНК в соответствующую последовательность аминокислот. В дополнение к этим основным трем классам существует множество типов РНК со специальными функциями, что более подробно описано в части III.
В состав нуклеотидов и нуклеиновых кислот входят определенные основания и пентозы
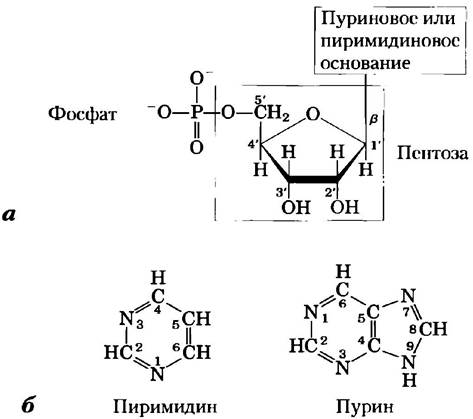
Нуклеотиды состоят из трех характерных компонентов: (1) азотистого (азотсодержащего) основания, (2) сахара пентозы и (3) фосфата (рис. 8-1). Молекулу без фосфатной группы называют нуклеозидом. Азотистыми основаниями являются производные двух веществ: пиримидина и пурина. В типичных нуклеотидах основания и пентозы— гетероциклические соединения.
Рис. 8-1. Структура нуклеотидов, а) Принятая нумерация атомов пентозы. Это рибонуклеотид. В дезоксирибонуклеотидах ОН-группа при 2'-атоме углерода (обозначена красным) замещена на -Н. б) Принятая нумерация атомов циклов пурина и пиримидина — соединений, которые служат основой для пуриновых и пиримидиновых оснований нуклеотидов и нуклеиновых кислот.
Ключевые договоренности.
Атомы углерода и азота в исходных структурах пронумерованы общепринятым способом, чтобы облегчить наименование и идентификацию многочисленных производных. Нумерация атомов в пентозном кольце основана на правилах, приведенных в гл. 7, но в пентозах нуклеотидов к номерам атомов углерода добавляется еще знак штриха ('), чтобы отличать их от атомов азотистых оснований. ■
Основание нуклеотида ковалентно соединено (через атом N-1 пиримидинов и атом N-9 пуринов) N-β-гликозидной связью с 1'-углеродом пентозы. 5'-углеродный атом пентозы, в свою очередь, соединяется эфирной связью с фосфатом. N-β-гликозидная связь образуется при удалении воды (гидроксильной группы пентозы и водорода основания), как и О-гликозидная связь (рис. 7-29).
И ДНК, и РНК содержат два главных пуриновых основания: аденин (А) и гуанин (G) и два главных пиримидиновых основания: в ДНК и в РНК — это цитозин (С), но второе пиримидиновое основание различается, в ДНК — это тимин (Т), а в РНК — урацил (U). Изредка тимин встречается в РНК, а урацил в ДНК. Структуры пяти главных оснований показаны на рис. 8-2, а номенклатура соответствующих им нуклеотидов и нуклеозидов представлена в табл. 8-1.
Рис. 8-2. Основные пуриновые и пиримидиновые основания нуклеиновых кислот. Некоторые из этих названий отражают историю их открытия. Гуанин, например, изначально был выделен из гуано (птичьего помета), а тимин — из тимуса.
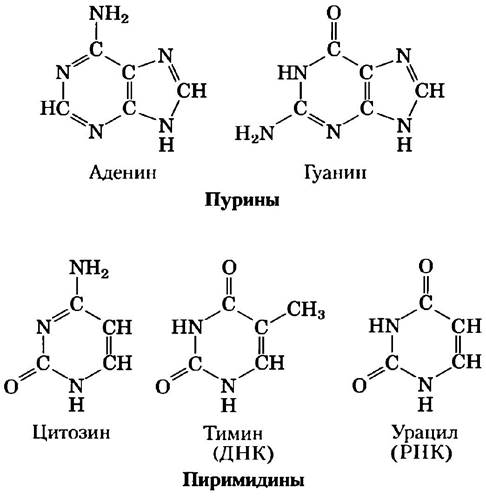
Таблица 8-1. Нуклеотиды и номенклатура нуклеиновых кислот
Основание |
Нуклеозид |
Нуклеотид |
Нуклеиновая кислота |
Пурины |
|||
Аденин |
Аденозин Дезоксиаденозин |
Аденилат Дезоксиаденилат |
РНК ДНК |
Гуанин |
Гуанозин Дезоксигуанозин |
Гуанилат Дезоксигуанилат |
РНК ДНК |
Пиримидины |
|||
Цитозин |
Цитидин Дезоксицитидин |
Цитидилат Дезоксицитидилат |
РНК ДНК |
Тимин |
Тимидин или дезокситимидин |
Тимидилат или дезокситимидилат |
ДНК |
Урацил |
Уридин |
Уридилат |
РНК |
Обратите внимание на то, что «нуклеозид» и «нуклеотид» — общие названия, которые включают в себя и рибо-, и деэоксирибоформы. Кроме того, рибонуклеозиды и рибонуклеотиды здесь просто названы нуклеозидами и нуклеотидами (например, рибоаденозин — отоаденозин), а дезоксирибонуклеозиды и дезоксирибонуклеотиды — дезоксинуклеозидами и дезоксинуклеотидами (например, дезоксирибоаденозин — дезоксиаденозин). Оба способа названий приняты, но укороченные варианты используются чаще. Исключением является тимин; термин «риботимидин» используется только в тех случаях, когда тимин включается в структуру РНК.
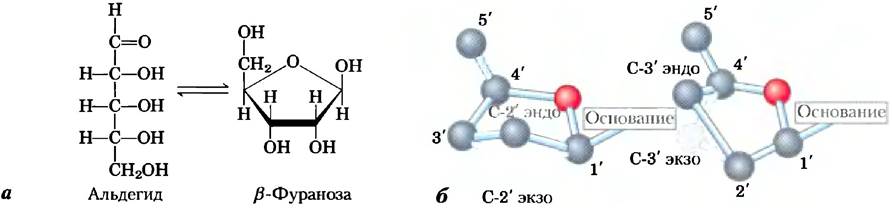
В состав нуклеиновых кислот входит два типа пентоз. Повторяющиеся дезоксирибонуклеотидные единицы ДНК содержат 2'-дезокси- D-рибозу, а рибонуклеотидные единицы РНК — D-рибозу. В нуклеотидах оба типа пентоз находятся в β-фуранозной форме (замкнутое пятичленное кольцо). Как показано на рис. 8-3, атомы пентозного цикла расположены не в одной плоскости, а находятся в одной из складчатых конформаций.
Рис. 8-3. Конформация рибозы. а) В растворе свободная рибоза принимает две формы: развернутая цепь (альдегид) и замкнутый цикл (β-фураноза), которые находятся в равновесии. В РНК рибоза содержится только в виде цикла, β-D-рибофуранозы. Дезоксирибоза подвергается похожим изменениям в растворе, но в состав ДНК входит только в виде β-2'-дезокси-D-фуранозы. б) Рибофурановый цикл, соединенный с нуклеотидом, может существовать в одной из четырех складчатых конформаций. Во всех случаях четыре из пяти атомов находятся в одной плоскости. Пятый атом (С-2' или С-3') располагается с той же (эндо-) или с другой (экзо-) стороны относительно С-5' атома.
Ключевые договоренности.
Хотя между ДНК и РНК есть два основных отличия — разные пентозы и присутствие урацила в РНК и тимидина в ДНК — именно пентозы определяют особенности этих двух типов кислот. Если нуклеиновая кислота содержит 2'-дезокси-D-рибозу, она по определению является ДНК, даже если в ней есть сколько-то остатков урацила. Аналогичным образом, если нуклеиновая кислота содержит D-рибозу, она представляет собой РНК вне зависимости от нуклеотидного состава. ■
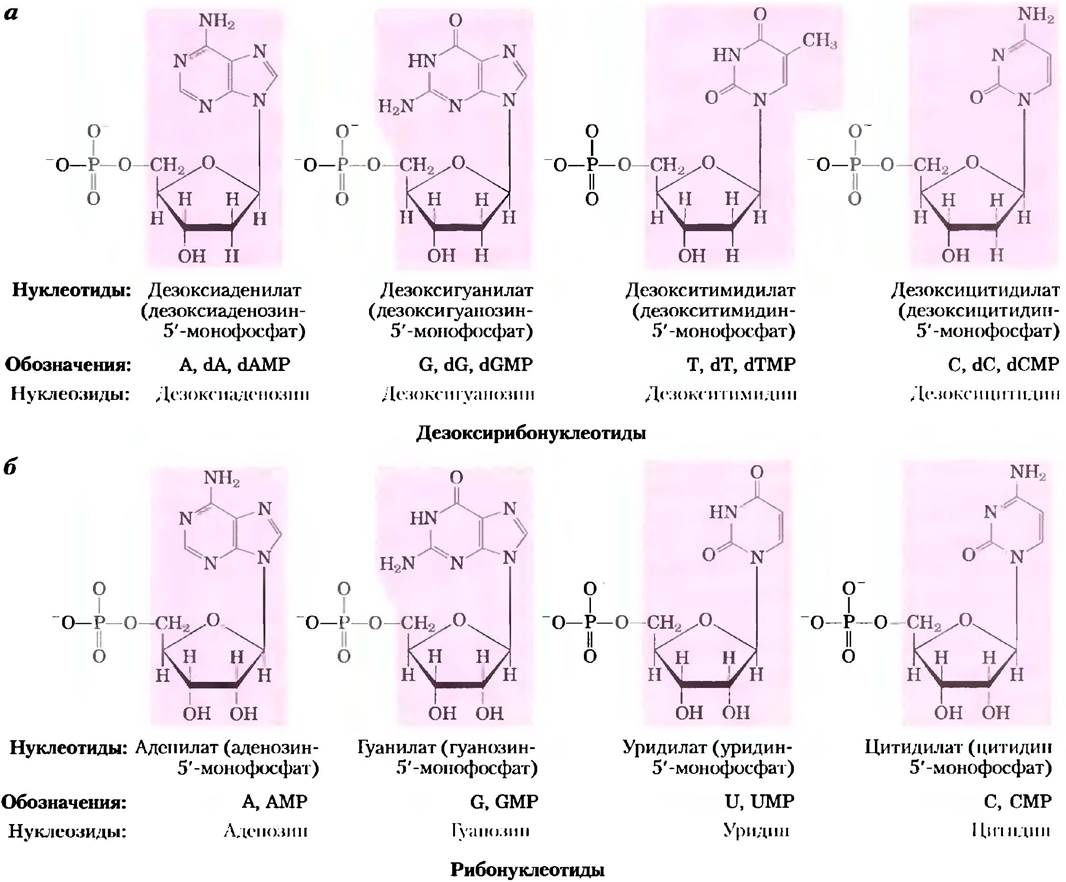
На рис. 8-4. даны структуры и названия четырех главных дезоксирибонуклеотидов (рибонуклеозид-5'-монофосфатов), структурных единиц ДНК, и четырех главных рибонуклеотидов (рибонуклеозид-5'-монофосфатов), структурных единиц РНК.
Рис. 8-4. Дезоксирибонуклеотиды и рибонуклеотиды нуклеиновых кислот. Все нуклеотиды показаны в свободной форме при pH = 7,0. Нуклеотиды ДНК (о) обычно обозначают как А, G, Т и С, а иногда как dA, dG, dT и dC; а РНК — (б) A, G, U и С. В свободной форме нуклеотиды обычно обозначаются как dAMP, dGMP, dTMP и dCMP; а рибонуклеотиды — AMP, GMP, UMP СМР. Для каждого нуклеотида наиболее часто встречающееся название указано перед полным названием в круглых скобках. Во всех сокращениях подразумевается, что фосфатная группа находится в 5'-положении. Нуклеозидная часть каждой молекулы выделена розовой рамкой. На этом и следующих рисунках атомы углерода углеводного кольца не показаны.
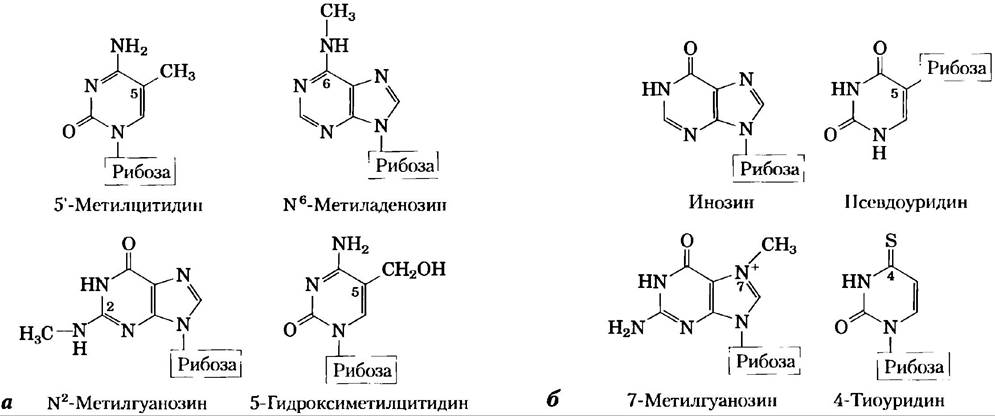
Хотя в состав нуклеотидов входят в основном главные пурины и пиримидины, и в ДНК, и в РНК встречаются некоторые минорные основания (рис. 8-5). В ДНК это чаще всего метилированные формы главных оснований, также в некоторых вирусных ДНК определенные основания могут быть гидроксиметилированы или гликозилированы. Модифицированные или нестандартные основания в молекуле ДНК часто участвуют в регуляции или защите генетической информации. Различные минорные основания найдены также в РНК, особенно в тРНК (см. рис. 8-25 и 26-23).
Ключевые договоренности.
Номенклатура минорных оснований в некотором смысле запутанная. Как и главные основания, большинство из минорных имеют тривиальные названия, например, на рис. 8-5 показана структура гипоксантина в составе нуклеозида инозина. При замещении атома в пуриновом или пиримидиновом цикле в названии нового соединения просто указывается заместитель и номер атома, к которому он присоединен, например, 5-метилцитозин, 7-мстилгуанин и 5-гидроксиметилцитозин (показаны в виде нуклеозидов на рис. 8-5). Элемент, к которому присоединен заместитель (N, С, О), не указывается. Номенклатура меняется, если замещенный атом находится вне циклической структуры; в таком случае указывается тип атома, к которому присоединен заместитель, а также номер атома в кольце в виде верхнего индекса. Атом азота аминогруппы, присоединенной к С-6 атому аденина, называется N6; аналогичным образом, карбонильный кислород и азот аминогруппы, присоединенные к С-6 и С-2 атомам гуанина, называются О6 и N2соответственно. Примерами могут служить N6- метиладенозин и N2-метилгуанозин (рис. 8-5). ■
Рис. 8-5. Некоторые минорные пуриновые и пиримидиновые основания (изредка встречающиеся в нуклеиновых кислотах основания) показаны в виде нуклеозидов. а) Минорные основания ДНК. 5-Метилцитидин встречается в ДНК животных и высших растений, N6-метиладенозин в бактериальной ДНК, а 5-гидроксиметилцитидин в ДНК бактерии, инфицированной определенными бактериофагами. б) Некоторые минорные основания тРНК. В состав инозина входит основание гипоксантин. Обратите внимание на то, что псевдоуридин, как и уридин, содержит урацил; эти соединения различаются между собой местом соединения основания с рибозой — в уридине рибоза связана с N-1 атомом урацила, как и в других пиримидинсодержащих нуклеозидах; в псевдоуридине — с С-5 атомом.
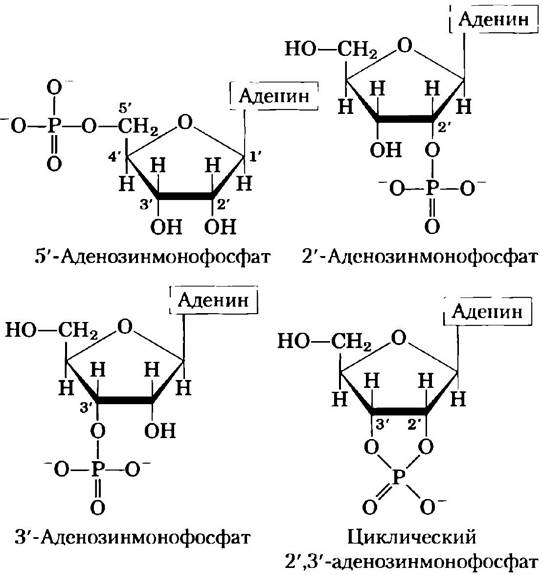
Клетки также содержат нуклеотиды, в которых фосфатные группы соединены не с 5'-углеродом (рис. 8-6). Циклические 2',3'- рибонуклеозидмонофосфаты — это стабильные интермедиаты, которые можно выделить, а 3'- рибонуклеозидмонофосфаты в качестве конечных продуктов образуются при гидролизе РНК определенными рибонуклеазами в качестве конечных продуктов. Другие случаи, такие как циклический 3',5'-аденозинмонофосфат (сАМР) и циклический 3',5'- гуанозинмонофосфат (сGМР), рассматриваются в конце этой главы.
Рис. 8-6. Некоторые монофосфаты аденозина. 2'-Аденозинмонофосфат, 3'-аденозинмонофосфат и циклический 2',3'-аденозинмонофосфат образуются под действием специфических ферментов и при щелочном гидролизе РНК.
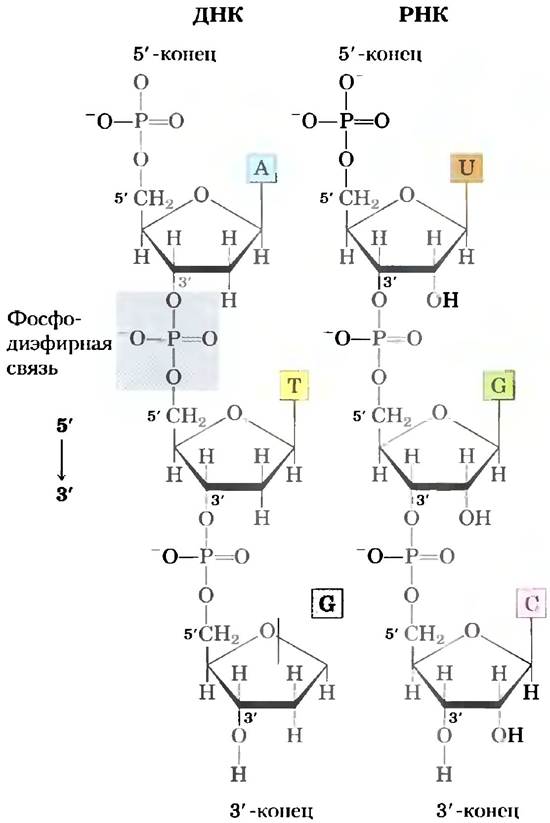
Нуклеотиды в нуклеиновых кислотах последовательно связываются фосфодиэфирными связями
Нуклеотиды в ДНК и в РНК ковалентно связаны между собой фосфодиэфирными мостиками, в которых 5'-фосфатная группа одного нуклеотида соединяется с 3'-гидроксильной группой следующего нуклеотида с образованием фосфодиэфирной связи (рис. 8-7). Таким образом, ковалентный остов нуклеиновых кислот состоит из чередующихся остатков фосфата и пентозы, а азотистые основания можно рассматривать как боковые группы, присоединенные к остову регулярно через одинаковые интервалы. Остовы ДНК и РНК гидрофильны. Гидроксильные группы остатков сахара образуют водородные связи с водой. Фосфатные группы имеют рКa - 0 и полностью ионизированы при pH 7, их отрицательные заряды нейтрализуются в основном за счет ионных взаимодействий с положительными зарядами белков, ионов металлов и полиаминов.
Ключевые договоренности.
Все фосфодиэфирные связи в ДНК и РНК направлены одинаково (рис. 8-7), благодаря чему каждая цепочка нуклеиновых кислот ориентирована и имеет разные 3'- и 5'-концы. По определению, у 5'-конца отсутствует нуклеотид в 5'-положении, а у 3'-конца отсутствует нуклеотид в 3'-положении. На одном или обоих концах могут присутствовать другие группы (чаще один или несколько фосфатов). Обозначение ориентации последовательности нуклеиновой кислоты от 5' к 3' относится к концам последовательности, а не к ориентации каждой отдельной фосфодиэфирной связи, соединяющей между собой нуклеотиды. ■
Рис. 8-7. Фосфодиэфирные связи в ковалентном остове ДНК и РНК. Фосфодиэфирные связи (одна из которых выделена в молекуле ДНК) последовательно соединяют нуклеотиды между собой. Остов из чередующихся пентозных и фосфатных групп обладает ярковыраженной полярностью и в ДНК, и в РНК. На 5'-конце макромолекулы отсутствует нуклеотид в 5'-положении, а на 3'-конце отсутствует нуклеотид в 3'-положении.
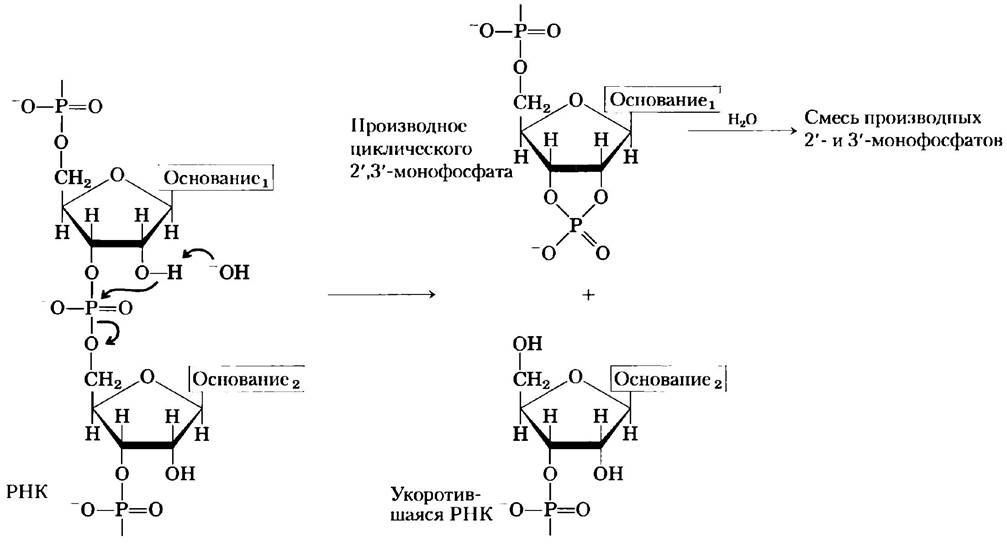
Фосфодиэфирные связи в молекулах ДНК и РНК подвергаются медленному неферментативному гидролизу. В пробирке РНК гидролизуется довольно быстро в щелочной среде, а ДНК — нет; в этом процессе принимают участие непосредственно 2'-гидроксильные группы РНК (которых нет в ДНК). Первыми продуктами действия щелочи на РНК являются циклические 2',3'- пуклеотидмонофосфаты, которые быстро гидролизуются с образованием смеси 2'-нуклеозид- и 3'-нуклеозидмонофосфатов (рис. 8-8).
Рис. 8-8. Гидролиз РНК в щелочных условиях. 2'-Гидроксильная группа играет роль нуклеофила при внутримолекулярном смещении электронной плотности. 2',3'-Производное циклического монофосфата гидролизуется до смеси 2'- и 3'-монофосфатов. ДНК, в которой отсутствует 2'-гидроксильная группа, в аналогичных условиях стабильна.
Последовательность нуклеотидов в нуклеиновых кислотах можно представить схематически, как показано на следующей странице на примере участка ДНК из пяти нуклеотидов. Фосфатные группы изображены как![]() каждая дезоксирибоза обозначена вертикальной линией от атома С-1' сверху до атома С-5' внизу (но помните, что сахар в нуклеиновых кислотах всегда находится в замкнутой (3-фуранозной форме). Связи между нуклеотидами (которые проходят через
каждая дезоксирибоза обозначена вертикальной линией от атома С-1' сверху до атома С-5' внизу (но помните, что сахар в нуклеиновых кислотах всегда находится в замкнутой (3-фуранозной форме). Связи между нуклеотидами (которые проходят через![]() ) изображены диагональю от середины (С-3') дезоксирибазы одного нуклеотида до низа (С-5') следующего.
) изображены диагональю от середины (С-3') дезоксирибазы одного нуклеотида до низа (С-5') следующего.
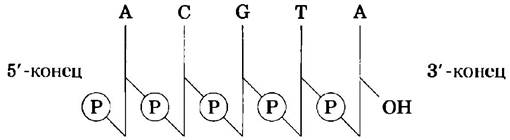
Вот примеры других упрощенных обозначений: рА-С-G-Т-Аон, рАрСрGрТрА и рАСGТА.
Ключевые договоренности.
Исторически сложилось, что последовательность одной цепи нуклеиновой кислоты всегда изображают, начиная с 5'-конца и заканчивая 3'-концом слева направо в направлении 5' —> 3'. ■
Короткие нуклеиновые кислоты называются олигонуклеотидами. Определение «короткие» в некотором смысле произвольное, но полимеры, содержащие 50 и менее нуклеотидов, обычно называются олигонуклеотидами. Болес длинные цепочки нуклеиновых кислот называются полинуклеотидами.
Свойства оснований нуклеотидов влияют на трехмерную структуру нуклеиновых кислот
Свободные пиримидины и пурины обладают слабовыраженными основными свойствами и поэтому называются основаниями. Пурины и пиримидины, встречающиеся в ДНК и в РНК, представляют собой молекулы с сопряженными связями (рис. 8-2), что оказывает большое влияние на структуру, распределение электронов и спектр поглощения света нуклеиновыми кислотами. Делокализация электронов атомов кольца придает большей части связей характер полуторных. Вследствие этого молекулы пиримидинов плоские, пуринов — почти плоские (с одним изгибом). Свободные основания пуринов и пиримидинов могут существовать в двух или более таутомерных формах в зависимости от pH. Урацил, например, может существовать в лактамной, лактимной и дважды лактимной формах (рис. 8-9). На рис. 8-2 показаны структуры, преобладающие в растворе при pH 7,0. Из-за резонанса все азотистые основания поглощают УФ-свет, нуклеиновые кислоты характеризуются максимумом поглощения в районе 260 нм (рис. 8-10).
Рис. 8-9. Таутомерные формы урацила. Лактамная форма преобладает при pH 7,0; другие формы становятся более заметными при понижении pH. Другие пиримидины и пурины в свободном состоянии также могут принимать различные таутомерные формы, но они встречаются намного реже.
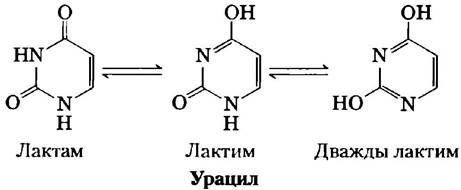
Рис. 8-10. Спектр поглощения основных нуклеотидов. Спектр показан как зависимость коэффициента молярной экстинкции от длины волны. Коэффициенты молярной экстинкции при 260 нм и pH 7,0 (ε260) приведены в таблице. Спектры соответствующих рибонуклеотидов и дезок- сирибонуклеотидов, так же, как и нуклеозидов, практически идентичны. Для измерения поглощения смеси нуклеотидов используется длина волны 260 нм (пунктирная вертикальная линия).
Пуриновые и пиримидиновые основания гидрофобии и плохо растворимы в воде при близких к нейтральным значениях pH в клетке. В кислых и щелочных средах основания становятся заряженными, и увеличивается их растворимость в воде. Гидрофобные взаимодействия между соседними основаниями, плоскости колец которых расположены параллельно друг другу (как стопка монет), представляют собой один из двух наиважнейших типов нековалентных связей между основаниями в нуклеиновых кислотах. Связь между параллельными плоскостями оснований (так называемые стекинговые взаимодействия, от англ, stacking interactions) осуществляется также за счет ван-дер-ваальсовых сил и диполь-дипольных взаимодействий. Упаковка оснований в виде стопки монет приводит к уменьшению количества контактов оснований с водой, а взаимодействия между параллельными основаниями очень важны для стабилизации трехмерной структуры нуклеиновых кислот, как будет описано ниже.
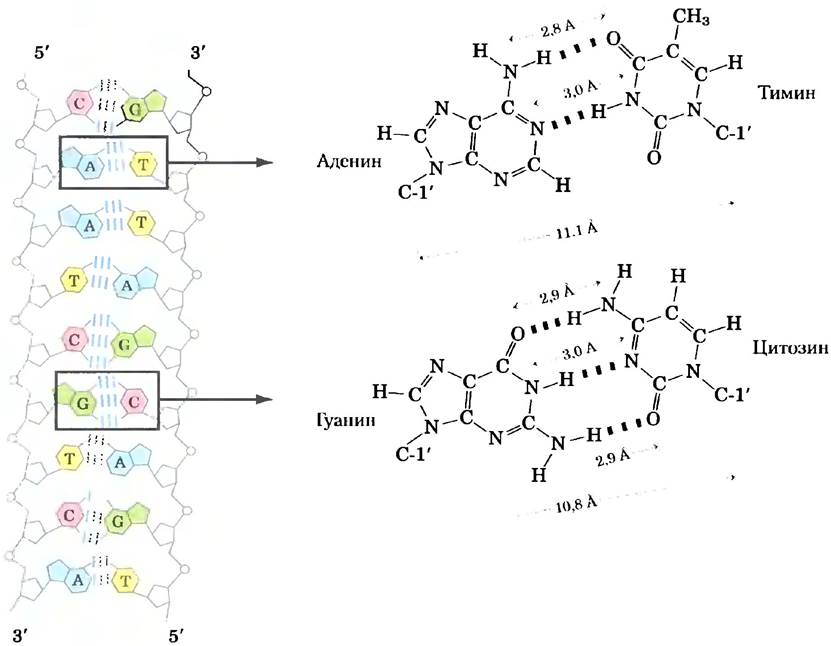
Наиболее важные функциональные группы пиримидинов и пуринов — это атомы азота кольца, карбонильные группы и аминогруппы, не участвующие в образовании цикла. Водородные связи между амино- и карбонильными группами служат вторым важнейшим типом взаимодействий между основаниями в нуклеиновых кислотах. Водородные связи между основаниями обеспечивают комплементарную ассоциацию между двумя (и изредка тремя или четырьмя) спиралями нуклеиновых кислот. Атомы, участвующие в образовании водородных связей, были определены Джеймсом Д. Уотсоном и Френсисом Криком в 1953 г. Они выяснили, что аденин А специфически связывается с тимином Т (или U), а гуанин G — с цитозином С (рис. 8-11). Эти два типа пар оснований преобладают в структуре двойных спиралей ДНК и РНК, за такой тип организации нуклеиновых кислот ответственны таутомеры, показанные на рис. 8-2. Это специфическое образование пар оснований обеспечивает стабильность процесса удвоения генетической информации, как мы увидим позже в этой главе.

Рис. 8-11. Водородные связи в парах оснований, определенные Уотсоном и Криком. Здесь, как и везде, водородные связи изображены тремя голубыми линиями.
Краткое содержание раздела 8.1 Основные понятия
■ Нуклеотиды содержат азотистые основания (пурины или пиримидины), пентозу и одну или несколько фосфатных групп. Нуклеиновые кислоты представляют собой полимеры нуклеотидов, соединенных друг с другом фосфодиэфирными связями между 5'-гидроксильной группой пентозы одного нуклеотида и 3'-гидроксильной группой другого.
■ Существуют два типа нуклеиновых кислот: РНК и ДНК. Нуклеотиды в РНК содержат рибозу и главные пиримидиновые основания уранил и цитозин. В ДНК нуклеотиды содержат 2'-дезоксирибозу и главные пиримидиновые основания тимин и цитозин. Типичными пуринами и в ДНК, и в РНК являются аденин и гуанин.