ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
ЧАСТЬ I. СТРОЕНИЕ И КАТАЛИЗ
Из всех природных систем живая материя является единственной, которая, несмотря на огромные преобразования, хранит в своей структуре самое большое количество собственного прошлого.
Эмиль Цукеркандл и Лайнус Полинг, статья в Journal of Theoretical Biology, 1965
9. ТЕХНОЛОГИЯ НА ОСНОВЕ ИНФОРМАЦИИ ИЗ ДНК
Обратимся к технологиям, которые лежат в основе прогресса современных биологических наук, определяют нынешние и будущие рубежи биохимии и иллюстрируют многие ее важные принципы. Объяснение законов, определяющих ферментативный катализ, структуру макромолекул, клеточный метаболизм и передачу информации, позволяет исследовать все более сложные биохимические процессы. Клеточное деление, иммунитет, эмбриогенез, зрение, вкус, онкогенез, познавательная способность — все это гармонично сочетается в искусно организованном оркестре молекулярных и макромолекулярных взаимодействий, которые мы теперь начинаем понимать все более ясно. Реальным результатом биохимического путешествия, начатого в XIX в., является постоянно растущая способность исследовать и изменять живые системы.
Для того чтобы попять сложный биохимический процесс, биохимик выделяет его и исследует отдельные компоненты in vitro, а затем соединяет их вместе, получая согласованную картину всего процесса в целом. Основным источником понимания его молекулярной сути является собственный информационный архив клетки, ее ДНК. Однако истинный размер хромосом создает гигантскую проблему: как найти и изучить отдельный ген среди десятка тысяч генов, включающих миллиарды пар оснований генома млекопитающих? Решения стали появляться в 1970-х гг.
Достижения десятилетий работы тысяч ученых — генетиков, биохимиков, клеточных биологов и физхимиков — были объединены в лабораториях Пола Берга, Герберта Бойера и Стенли Коэна, разработавших методы обнаружения, очистки, приготовления и изучения малых сегментов ДНК, полученных из намного превышающих их по размерам хромосом. Методы клонирования ДНК проложили дорогу в такие современные области, как геномика и протеомика, исследования генов и белков в масштабах целых клеток и организмов. Эти новые методы видоизменяют фундаментальные исследования, сельское хозяйство, медицину, экологию, судебную медицину и многие другие области, иногда ставя общество перед трудным выбором и этическими дилеммами.

Начнем эту главу с наиболее общего изложения фундаментальных биохимических принципов теперь уже классической методики клонирования ДНК, затем, после создания основы для обсуждения геномики, проиллюстрируем ряд применений и возможностей этих технологий, с основным акцентом на современные достижения в геномике и протеомике.
9.1. Клонирование ДНК: основные понятия
Клан — это идентичная копия. Этот термин первоначально употреблялся для клеток одного типа, выделенных и способных к воспроизводству с целью создания популяции идентичных клеток. Клонирование ДНК включает выделение специфического гена или сегмента ДНК из хромосомы, присоединение его к малой молекуле-носителю ДНК, а затем копирование этой модифицированной ДНК тысячи или миллионы раз, посредством как увеличения числа клеток, так и создания множественных копий клонируемой ДНК в каждой клетке. Результатом является избирательная амплификация данного гена или сегмента ДНК. Клонирование ДНК из любого организма включает пять основных процедур.
1. Вырезание ДНК в точных позициях. Сайт- специфичные эндонуклеазы (эндонуклеазы рестрикции) обеспечивают необходимые молекулярные «ножницы».
2. Выбор малой молекулы ДНК, способной к саморепликации. Эти ДНК называются клонирующими векторами (вектор — фактор доставки); обычно это плазмиды или вирусные ДНК.
3. Ковалентное соединение двух фрагментов ДНК. Фермент ДНК-лигаза сшивает клонирующий вектор с ДНК, которую надо клонировать. Смешанные молекулы ДНК, содержащие ковалентно связанные сегменты из двух или более источников, называются рекомбинантными ДНК.
4. Перемещение рекомбинантной ДНК из пробирки в клетку-хозяина, предоставляющую ферментативный аппарат репликации ДНК.
5. Отбор или идентификация клеток-хозяев, содержащих рекомбинантную ДНК.
Набор методов, используемых для выполнения этих и подобных процедур, называют технологией рекомбинантных ДНК или, более неформально, генной инженерией.
Большая часть нашего начального изложения будет сфокусирована на клонировании ДНК в бактерии Escherichia coli, первом организме, использованном для работы с рекомбинантными ДНК и до сих пор наиболее распространенной клеткой-хозяином. У Е. coli есть много преимуществ: метаболизм ее ДНК (как и многих других се биохимических процессов) хорошо изучен; многие имеющиеся в природе клонирующие векторы, связанные с Е. coli, такие как плазмиды и бактериофаги (бактериальные вирусы; называемые также фагами), хорошо охарактеризованы; имеются методики быстрого переноса ДНК из одной бактериальной клетки в другую. Мы также обратимся к клонированию ДНК в других организмах; эта тема более полно обсуждается ниже.
Эндонуклеазы рестрикции и ДНК-лигаза создают рекомбинантную ДНК
Очень важным для технологии рекомбинантной ДНК является набор ферментов (табл. 9-1), ставших доступными после десятилетий исследований метаболизма нуклеиновых кислот. Два класса ферментов лежат в основе основного подхода к созданию и «размножению» молекулы рекомбинантной ДНК (рис. 9-1). Во-первых, эндонуклеазы рестрикции (рестриктазы) распознают и разрезают ДНК в местах специфических нуклеотидных последовательностей (нуклеотидные последовательности распознавания или сайты рестрикции), создавая набор более маленьких фрагментов. Во- вторых, клонируемый фрагмент ДНК может быть встроен в подходящий клонирующий вектор с использованием ДНК-лигазы для соединения молекул ДНК. Затем рекомбинантный вектор вводится в клетку-хозяина, которая размножает фрагмент в ходе множества стадий клеточного деления.
Таблица 9-1. Некоторые ферменты, используемые в технологии рекомбинантных ДНК
Фермент(ы) |
Функция |
Эндонуклеазы рестрикции типа II |
Разрезают ДНК в местах специфической последовательности основании |
ДНК-лигаза |
Соединяет две молекулы или фрагмента ДНК |
ДНК-полимераза I (Е. coli) |
Заполняет пробелы в дуплексах, поэтапно добавляя нуклеотиды к 3'-концам |
Обратная транскриптаза |
Создает ДПК-копию молекулы РНК |
Полинуклеотидкиназа |
Присоединяет фосфат к 5'-ОН концу полинуклеотида, помечая его или осуществляя сшивание |
Терминальная трансфераза |
Добавляет гомонополимерные «хвосты» к 3'-ОН концам линейного дуплекса |
Экзонуклеаза III |
Отщепляет нуклеотидные остатки с 3'-концов цепочки ДНК |
Экзонуклеаза бактериофага λ |
Отщепляет нуклеотиды с 5'-копцов дуплекса, выставляя одноцепочечные 3'-концы |
Щелочная фосфатаза |
Отщепляет герминальные фосфаты иди с 5'-, и. и с 3'-конца (или с обоих) |
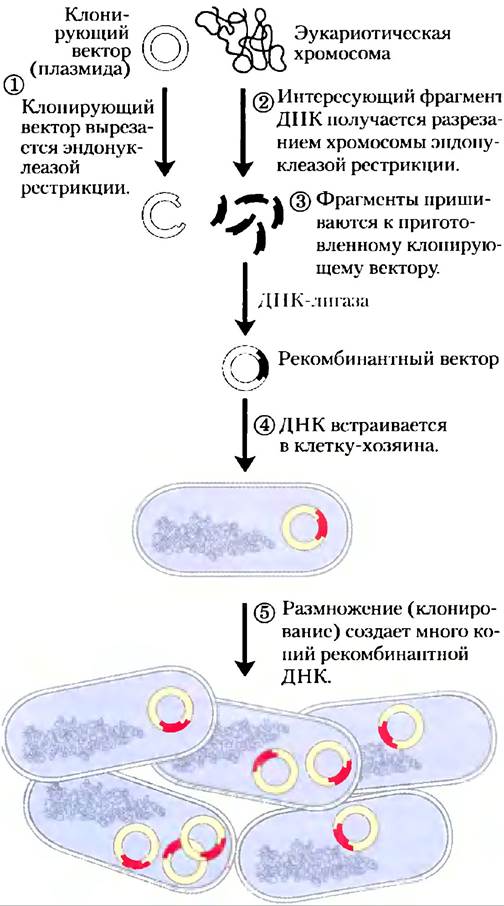
Рис. 9-1. Схема клонирования ДНК. Клонирующий вектор и эукариотические хромосомы независимо друг от друга разрезаются одной и той же эндонуклеазой рестрикции. Затем фрагменты, которые надо клонировать, встраиваются в клонирующий вектор. Получившаяся рекомбинантная ДНК (показан только один рекомбинантный вектор) встраивается в клетку-хозяина, где она размножается (клонируется). Внимание! Масштаб не сохранен: размер хромосомы Е. coli по сравнению с размером типичного клонирующего вектора (такого как плазмида) на самом деле намного больше.
Эндонуклеазы рестрикции обнаружены у самых разнообразных бактерий. В начале 1960-х гг. Вернер Арбер открыл, что биологическая функция рестриктаз состоит в распознавании и разрезании чужеродных ДНК (например, ДНК инфицирующих вирусов), при этом говорят, что такие ДНК рестриктируются. В ДНК клетки-хозяина последовательность оснований, которая распознается ее собственной эндонуклеазой рестрикции, защищена от расщепления метилированием ДНК, катализируемом специфичной ДНК-метилазой. Эндонуклеаза рестрикции и соответствующая метил аза иногда называются системой рестрикции-модификации.
Известно три типа эндонуклеаз рестрикции: I, II и III. Рестриктазы I и III типа обычно представляют собой большие, мультисубъединичные комплексы, обладающие как эндонуклеазной, так и метилазной активностью. Эндонуклеазы рестрикции I типа разрезают ДНК в произвольном месте, которое может быть удалено более чем на 1000 пар оснований (1000 п. н.) от распознаваемой последовательности нуклеотидов. Эндонуклеазы рестрикции III типа разрезают ДНК примерно на расстоянии 25 п. н. от распознаваемой нуклеотидной последовательности. Рестриктазы обоих типов двигаются вдоль ДНК благодаря реакции, которая требует энергии АТР. Эндонуклеазы рестрикции II тина, впервые полученные Хамилтоном Смитом в 1970 г., удобнее, поскольку не требуют АТР и разрезают ДНК в пределах самой распознаваемой нуклеотидной последовательности. Исключительное удобство этой группы эндонуклеаз рестрикции было продемонстрировано Даниэлем Натансоном, который впервые использовал их для разработки новых методов картирования и исследования генов и геномов.
У различных видов бактерий были открыты тысячи эндонуклеаз рестрикции, причем один или несколько таких ферментов распознают более 100 различных последовательностей ДНК. Распознаваемые нуклеотидные последовательности обычно от 4 до 6 п. н. в длину и представляют собой палиндромы (см. рис. 8-18). В табл. 9-2 приведены последовательности оснований, распознаваемые некоторыми эндонуклеазами рестрикции типа II. В некоторых случаях взаимодействие между эндонуклеазой рестрикции и ее распознаваемой нуклеотидной последовательностью подробно объяснено на молекулярном уровне; например, в табл. 9-2 изображены последовательности, узнаваемые несколькими эндонуклеазами рестрикции типа II.
Таблица 9-2. Нуклеотидные последовательности, распознаваемые некоторыми эндонуклеазами рестрикции II типа

Стрелками указаны фосфодиэфирные связи, расщепляемые каждой эндонуклеазой рестрикции; звездочки — основания, которые метилируются соответствующей метилазой (где это известно); N — произвольное основание. Обратите внимание, что название каждого фермента состоит из трехбуквенной аббревиатуры (написано курсивом) бактериального вида, из которого он получен, за которой иногда следует обозначение штамма и римские цифры (для того чтобы отличить различные эндонуклеазы рестрикции, выделенные из одних и тех же бактериальных видов). Так, надпись BamIII означает, что это первая (I) эндонуклеаза рестрикции, полученная из штамма Н бактерии Bacillusamyloliquefaciens.
Некоторые эндонуклеазы рестрикции создают в двух цепях ДНК разрывы с уступом, оставляя от двух до четырех нуклеотидов одной цепи неспаренными в каждом получающемся конце. Эти неспаренные цепи называются «липкими» концами (рис. 9-2, а), потому что они могут комплементарно связаться друг с другом или с комплементарными «липкими» концами фрагментов других ДНК. Другие эндонуклеазы рестрикции разрезают обе цепочки ДНК в противоположных фосфодиэфирных связях, не оставляя неспаренных оснований на концах, такие концы часто называют «тупыми» (рис. 9-2, б).
Рис. 9-2. Разрезание молекул ДНК эндонуклеазами рестрикции. Эндонуклеазы рестрикции распознают и разрезают только специфические нуклеотидные последовательности, оставляя или (а) «липкие» концы (с торчащими одиночными цепями), или (б) «тупые» концы. Фрагменты могут быть присоединены к другим ДНК, таким как разрезанный клонирующий вектор (плазмида), показанный здесь. Этой реакции способствует гибридизация комплементарных «липких» концов. Присоединение (лигирование) менее эффективно для фрагментов ДНК с «тупыми» концами, чем с комплементарными «липкими», а фрагменты ДНК с разными (некомплементарными) «липкими» концами обычно вообще не лигируются. (в) В плазмиду, разрезанную рестриктазой, может быть встроен синтетический фрагмент ДНК с последовательностями, которые узнают несколько эндонуклеаз рестрикции. Вставка с одним сайтом рестрикции называется линкером, а с несколькими участками — полилинкером, в эндонуклеазы рестрикции

Средний размер фрагментов ДНК, полученных в результате разрезания геномной ДНК эндонуклеазой рестрикции, зависит от частоты, с которой специфический участок рестрикции встречается в молекуле ДНК, что, в свою очередь, сильно зависит от размера последовательности распознавания. В молекуле ДНК с произвольной нуклеотидной последовательностью, в которой все четыре основания присутствуют в равных количествах, последовательность, состоящая из 6 п. н« распознаваемая такой эндонуклеазой рестрикции как BamHI, встречалась бы в среднем один раз на каждые 46 (4096) п. н., если предположить, что ДНК содержит 50% G=С-пар. Ферменты, которые распознают последовательности длиной в 4 п. н., создавали бы более короткие фрагменты из молекулы ДНК с произвольной последовательностью, при этом последовательность распознавания такого размера встречалась бы около одного раза на каждые 44 (256) п. н. В природных молекулах ДНК специфические последовательности распознавания встречаются реже, потому что нуклеотидные последовательности в ДНК не произвольны, а четыре вида нуклеотидов содержатся не в равных количествах. В лабораторных экспериментах средний размер фрагментов, получаемых разрезанием большой молекулы ДНК эндонуклеазой рестрикции, может быть увеличен простой остановкой реакции до ее завершения, а результат называется неполной нарезкой. Размер фрагмента можно также увеличить при использовании особого класса эндонуклеаз, называемых хоуминг-эндонуклеазами (см. рис. 26-38). Они распознают и разрезают гораздо более длинные последовательности ДНК (от 14 до 20 п. п.).
После того как молекула ДНК была расщеплена на фрагменты, специфический фрагмент заданного размера можно получить электрофорезом в агарозном или полиакриламидном геле или с помощью ВЭЖХ (с. 135). Однако в случае типичного генома млекопитающих разрезание эндонуклеазой рестрикции обычно приводит к образованию большого числа различных фрагментов ДНК, а это затрудняет выделение специфического фрагмента электрофорезом или ВЭЖХ. Распространенным промежуточным шагом в клонировании специфического гена или сегмента ДНК является создание библиотеки ДНК (описывается в разд. 9.2).
После выделения необходимого фрагмента ДНК можно использовать ДНК-лигазу для присоединения его к расщепленному таким же образом клонирующему вектору, т. е. к вектору, полученному с помощью той же эндонуклеазы рестрикции. Например, фрагмент, полученный с помощью EcoRI, как правило, не связывается с фрагментом, полученным с помощью ВаmHI. Как это более подробно описано в гл. 25 (см. рис. 25-17), ДНК-лигаза катализирует образование новых фосфодиэфирных связей в реакции, использующей АТР или сходный с ним кофактор. Спаривание оснований комплементарных «липких» концов сильно облегчает лигирование (рис. 9-2, а). «Тупые» концы тоже можно лигировать, хотя и менее эффективно. Исследователи могут создавать новые последовательности ДНК, вставляя синтетические фрагменты ДНК (называемые линкерами) между лигируемыми концами. Встраиваемые фрагменты ДНК с множественными последовательностями распознавания эндонуклеазами рестрикции (обычно используемыми далее в качестве участков для встраивания дополнительных ДНК разрезанием и лигированием) называются полилинкерами (рис. 9-2, в).
Эффективность «липких» концов при избирательном соединении двух фрагментов ДНК стала очевидной в самых ранних экспериментах с рекомбинантными ДНК. Еще до того, как эндонуклеазы рестрикции стали широко распространенными, некоторые исследователи обнаружили, что возможно создавать «липкие» концы за счет комбинированного действия эндонуклеазы бактериофага λ и терминальной трансферазы (табл. 9-1). Соединяемые фрагменты содержали комплементарные гомополимерные «хвосты». Петер Лобан и Дэйл Кайзер использовали этот метод в 1971 г. в первых экспериментах по соединению фрагментов ДНК природного происхождения. Вскоре после этого подобные методы стали использоваться в лаборатории Пола Берга при соединении сегментов ДНК вируса обезьяны (SV40) с ДНК, полученной из бактериофага λ; таким образом была создана первая молекула рекомбинантной ДНК с сегментами, полученными от разных видов.
Клонирующие векторы позволяют амплифицировать встроенные сегменты ДНК
Принципы, которые определяют доставку рекомбинантной ДНК в клонируемой форме в клетку-хозяина и ее дальнейшую амплификацию, можно проиллюстрировать на примере трех распространенных клонирующих векторов, используемых обычно в экспериментах с Е. coli, — плазмид, бактериофагов и искусственных бактериальных хромосом — и вектора, используемого при клонировании больших сегментов ДНК в дрожжах.
Плазмиды. Плазмиды представляют собой кольцевые молекулы ДНК, которые реплицируются отдельно от хромосомы клетки-хозяина. В природе встречаются бактериальные плазмиды длиной от 5000 до 400 000 п. н. Они могут быть введены в клетки бактерий в ходе процесса, который называется трансформацией. Клетки (обычно это Е. coli) и плазмидные ДНК инкубируются вместе при температуре 0 °С в растворе хлорида кальция, а затем подвергаются шоку при быстром сдвиге температуры до 37-40 °С. По причине, до конца непонятной, некоторые клетки при таком воздействии поглощают плазмидную ДНК. Некоторые виды бактерий по своей природе способны к поглощению ДНК и не требуют воздействия хлорида кальция. В альтернативном методе инкубируемые с плазмидной ДНК клетки подвергаются действию высоковольтного импульса. При таком подходе, называемом электропорацией, бактериальные мембраны кратковременно делаются проницаемыми для больших молекул.
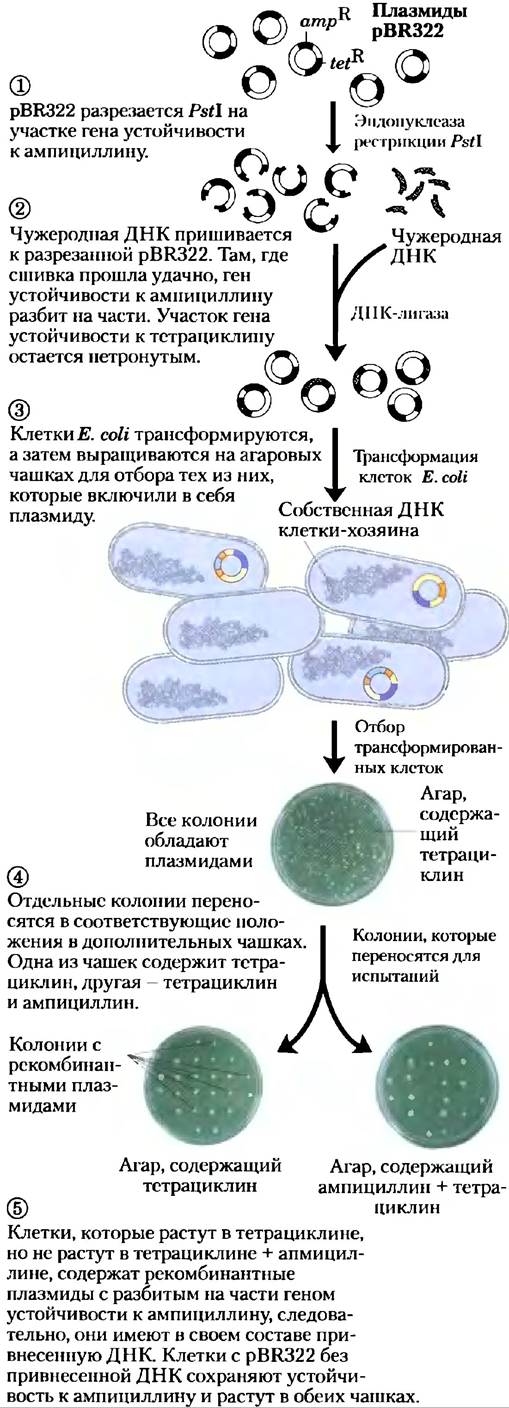
Независимо от метода, лишь немногие клетки поглощают плазмидную ДНК, поэтому требуется способ отбора удачно трансформированных. Распространенным подходом является использование плазмиды, содержащей ген, необходимый клетке-хозяину для роста в специфических условиях, такой как, скажем, ген устойчивости к какому-то антибиотику. Только клетки, трансформированные рекомбинантной плазмидой, могут расти в присутствии этого антибиотика, делающего всякую клетку, содержащую плазмиду, «селектируемой» при таких условиях роста. Такой ген называется селектируемым маркером.
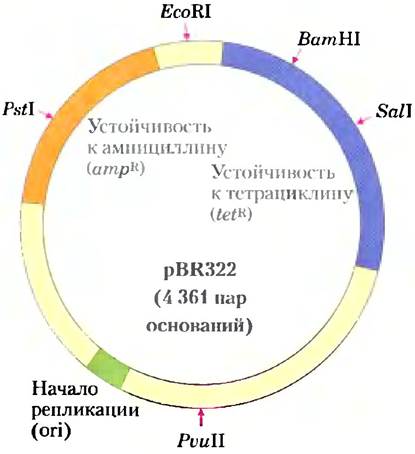
Путем изменения природных плазмид исследователи разработали много разных плазмидных векторов, пригодных для клонирования. Плазмида pBR322 Е. coli представляет собой хороший пример свойств, удобных для клонирующего вектора (рис. 9-3).
Рис. 9-3. Плазмида pBR322, созданная для Е. coli. Обратите внимание на расположение некоторых важных участков рестрикции — для PstI, EcoRI, BamHI, SalI и PvuII; на гены устойчивости к ампициллину и тетрациклину; и на репликатор (ori). Созданная в 1977 г., она стала одной из первых плазмид, специально разработанных для клонирования в Е. coli.
Важные преимущества плазмиды pBR322:
1. Участок начала репликации (ori) — последовательность, где инициируется репликация клеточными ферментами (гл. 25). Эта последовательность требуется для размножения плазмиды и поддержания ее в количестве 10-20 копий на одну клетку.
2. Два гена устойчивости к различным антибиотикам (tetR, ampR), что позволяет идентифицировать клетки, содержащие интактную плазмиду или ее рекомбинантный тип (рис. 9-4).
Рис. 9-4. Использование pBR322 для клонирования чужеродной ДНК в Е. coli и идентификации клеток, содержащих ее. Плазмидное клонирование
3. Некоторые уникальные последовательности распознавания (Pstl, EcoRI, BamHI, SalI, Pvull), которые являются мишенями различных эндонуклеаз рестрикции, предоставляя участки, где плазмида в дальнейшем может быть разрезана для вставки чужеродной ДНК.
4. Малый размер (4361 п. н.), который способствует ее проникновению в клетки и биохимическим манипуляциям с ДНК.
Трансформация типичных бактериальных клеток очищенной ДНК (сам по себе не очень результативный процесс) становится все менее успешной по мере увеличения размера плазмиды; поэтому становится трудно клонировать сегменты ДНК длиннее 15000 п. н. в том случае, если в качестве вектора используются плазмиды.
Бактериофаги. У бактериофага λ есть эффективный механизм переноса собственной ДНК (48502 п. н.) в бактерию, поэтому он может быть использован в качестве вектора при клонировании несколько больших сегментов ДНК (рис. 9-5). Удобным его применение делают две ключевые особенности:
1. Около трети генома λ несущественна и может быть заменена чужеродной ДНК.
2. ДНК упаковывается в частицы инфекционного фага только в том случае, если ее длина составляет от 40 000 до 53 000 п. н., причем это ограничение можно использовать для обеспечения упаковки только рекомбинантной ДНК.
Рис. 9-5. Клонирующие векторы на основе бактериофага λ. Для модификации генома бактериофага λ используются методы рекомбинантной ДНК — удаление ненужных для продуцирования фага генов и замена их ДНК-«наполнителем», чтобы сделать ДНК фага достаточно большой для помещения в частицы фага. Как показано здесь, в экспериментах по клонированию наполнитель заменяется чужеродной ДНК. Рекомбинанты упаковываются in vitro в жизнеспособные частицы фага только в том случае, если они содержат фрагмент чужеродной ДНК подходящего размера, а также принадлежащие бактериофагу λ концевые фрагменты ДНК.

Исследователи разработали векторы на основе бактериофага λ, которые могут быть легко расщеплены на три части таким образом, что две из них содержат необходимые гены бактериофага λ и вместе составляют всего около 30 000 п. и. в длину. Третья часть, ДНК-«наполнитель», выбрасывается, когда вектор нужно использовать для клонирования. Дополнительная ДНК вставляется между двумя необходимыми бактериофагу λ. сегментами, образуя лигированные молекулы ДНК длиной, достаточной для получения жизнеспособных частиц фага. Механизм упаковки используется для селекции рекомбинантных вирусных частиц.
Векторы на основе бактериофага λ позволяют клонировать фрагменты ДНК длиной до 23000 п. н. Как только фрагменты бактериофага λ лигированы с фрагментами чужеродной ДНК подходящего размера, получившиеся рекомбинантные ДНК могут быть упакованы в частицы фагов посредством добавления их к грубым экстрактам бактериальных клеток, которые содержат все необходимые белки для сборки целого фага. Этот процесс называется упаковкой in vitro (рис. 9-5). Все жизнеспособные частицы фагов будут содержать фрагмент чужеродной ДНК. Дальнейший процесс переноса рекомбинантной ДНК в клетки Е. coli чрезвычайно эффективен.
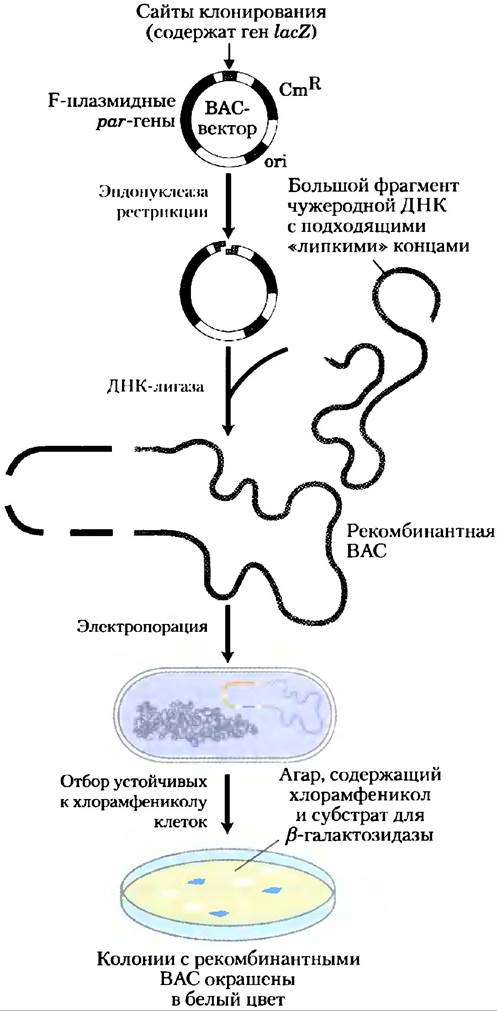
Искусственные бактериальные хромосомы (ВАС, от англ. bacterial artificial chromosomes). Искусственные бактериальные хромосомы представляют собой плазмиды, разработанные для клонирования очень длинных (как правило, от 100 000 до 300 000 н. н.) сегментов ДНК (рис. 9-6). Они обычно содержат селектируемые маркеры, такие как ген устойчивости к антибиотику хлорамфениколу (CmR), а также очень стабильный локус начала репликации (ori), поддерживающий число плазмид (1-2 копии на клетку). В ВАС-векторы встраивают фрагменты ДНК из нескольких сотен тысяч пар оснований. Затем огромные кольцевые ДНК с помощью электропореза помещают в бактерии-хозяева. В этих процедурах используют бактерии, содержащие мутации, которые изменяют структуру клеточной стенки, что позволяет поглощать большие молекулы ДНК.
Рис. 9-6. Искусственные бактериальные хромосомы (ВАС) в качестве клонирующих векторов. Вектор является относительно простой плазмидой с локусом начала репликации (ori), управляющим репликацией. Гены par, полученные из плазмид, называемых F-плазмидами, способствуют равномерному распределению плазмид по дочерним клеткам в процессе деления клетки. Это увеличивает вероятность того, что каждая дочерняя клетка будет нести одну копию плазмиды, даже при наличии малого числа копий. Небольшое число копий полезно при клонировании больших сегментов ДНК, поскольку это ограничивает возможности нежелательных реакций рекомбинации, которые могут со временем непредсказуемым образом менять большие клонируемые ДНК. Вектор ВАС включает селектируемые маркеры. Ген lacZ (необходимый для синтеза фермента β-галактозидазы) располагается в клонирующем участке так, что он дезактивируется вставками клонируемых ДНК. Встраиванию электропорацией рекомбинантных ВАС в клетки способствует использование клеток с измененными (более пористыми) клеточными стенками. Скрининг рекомбинантных ДНК производится на основании устойчивости клеток к антибиотику хлорамфениколу (CmR). Чашки, на которых выращивают бактерии, также содержат субстрат для β-галактозидазы, что приводит к окрашиванию продуктов. Колонии с активной β-галактозидазой и, следовательно, не имеющие вставок ДНК в ВАС-векторе становятся синими, а колонии не обладающие активностью β-галактозидазы — и, таким образом, имеющие искомые вставки ДНК, — белыми.
Искусственные хромосомы дрожжей (YAC, от англ. yeast artificial chromosomes). В генной инженерии клетки Е. coli вовсе не единственные клетки- хозяева. Дрожжи — чрезвычайно удобные эукариотические организмы для такой работы. Как и в случае с Е. coli, генетика дрожжей хорошо развита. Геном наиболее часто используемых дрожжей Saccharomyces cerevisiae содержит всего 14 • 106 п. н. (по эукариотическим стандартам, это небольшой геном, менее чем в четыре раза превышает размер хромосомы Е. coli);вся его нуклеотидная последовательность известна. Дрожжи также очень легко содержать и выращивать в больших масштабах в лабораторных условиях. Для дрожжей были созданы плазмидные векторы, используя те же самые подходы, которые лежали в основе применения рассмотренных выше векторов Е. coli. В настоящее время в распоряжении имеются удобные методы переноса ДНК как из дрожжевых клеток, так и в них, что способствует изучению многих аспектов биохимии эукариотической клетки. Некоторые рекомбинантные плазмиды содержат несколько ori и других элементов, что дает возможность использовать их более чем для одного вида организмов (например, в клетках дрожжей или Е. coli). Плазмиды, которые могут реплицироваться в клетках двух или более различных видов, называются шаттл- векторами.
Исследования больших геномов и связанная с этим потребность в клонирующих векторах высокой емкости привели к созданию искусственных хромосом дрожжей(YAC; рис. 9-7). YAC-векторы включают в себя все необходимые элементы для жизнеобеспечения эукариотической хромосомы в ядре дрожжей: дрожжевой ori, два селектируемых маркера и специфические последовательности (полученные из теломер и центромер, участков ДНК, о которых рассказывается в гл. 24), необходимые для стабильности и правильного расхождения хромосом при делении клетки. Прежде чем вектор будет использован в клонировании, он амплифицируется в виде кольцевой бактериальной плазмиды. Разрезание эндонуклеазой рестрикции (ВатШ на рис. 9-7) удаляет участок ДНК между двумя теломерными последовательностями (TEL), оставляя теломеры на концах линеаризованной ДНК. Разрезание в другом участке (EcoRI на рис. 9-7) делит вектор на два сегмента ДНК, называемых векторными «рукавами», каждый из которых содержит свой селектируемый маркер.
Рис. 9-7. Конструирование искусственной хромосомы дрожжей (YAC). YAC-вектор содержит локус начала репликации (ori), центромеру (CEN), две теломеры (TEL) и селектируемые маркеры (X и Y). Расщепление эндонуклеазами ВаmHI и EcoRI создает два отдельных ДНК-«рукава»; каждый из них имеет один теломерный конец и один селектируемый маркер. Один большой сегмент ДНК (например, до 2 • 106 п. н. из генома человека) лигируется с двумя «рукавами», образуя искусственную хромосому дрожжей. YAC трансформирует дрожжевые клетки (сферопласты с заранее удаленнымим клеточными стенками), и они селектируются по X и Y; уцелевшие клетки реплицируют ДНК-вставку.
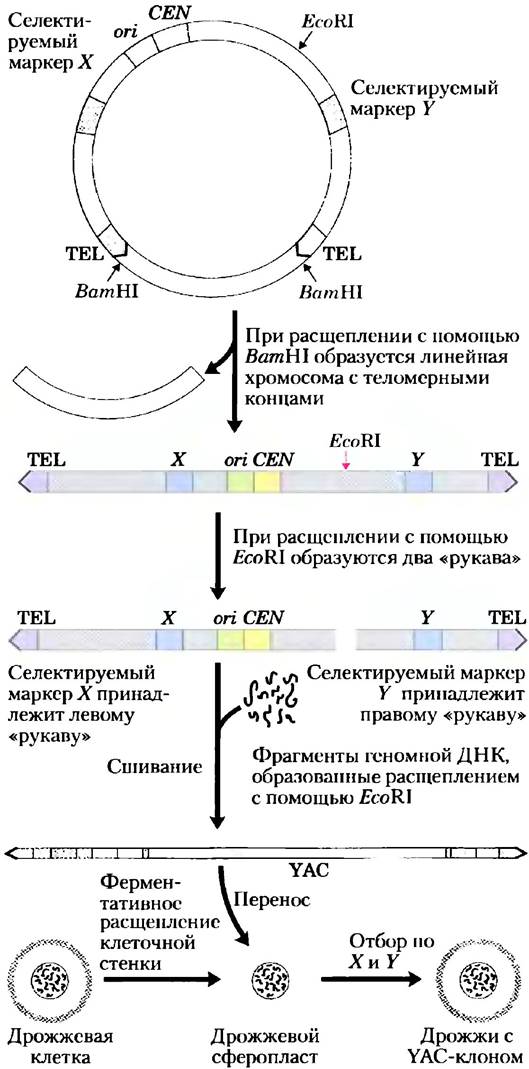
Геномная ДНК подвергается неполному расщеплению эндонуклеазами рестрикции (EcoRI на рис. 9-7), чтобы получился фрагмент подходящего размера. Затем геномные фрагменты разделяются пульсирующим гель-электрофорезом (разновидностью гель-электрофореза; см. рис. 3-18), который позволяет разделять очень большие сегменты ДНК. Фрагменты ДНК подходящего размера (до 2 • 106 п. н.) смешиваются с полученными «рукавами» векторов и лигируются. Затем полученная смесь используется для трансформации обработанных дрожжевых клеток очень большими молекулами ДНК. Выращивание на среде, для роста на которой необходимо наличие генов обоих селектируемых маркеров, обеспечивает отбор только тех дрожжевых клеток, у которых есть искусственная хромосома с большой вставкой между двумя «рукавами» вектора (рис. 9-7). Стабильность YAC-клонов увеличивается с размером. Те YAC-клоны, у которых размер вставки превышает 100 000 п. н., примерно так же стабильны, как и обычные хромосомы клетки, в то время как другие — со вставкой менее 100 000 п. н. — постепенно гибнут во время митоза (так, например, дрожжевые клоны, имеющие всего два векторных конца, сшитых вместе или с короткими вставками, обычно не обнаруживаются). YAC, теряющие теломеры на обоих концах, быстро разрушаются.
Специфические последовательности ДНК определяют при гибридизации
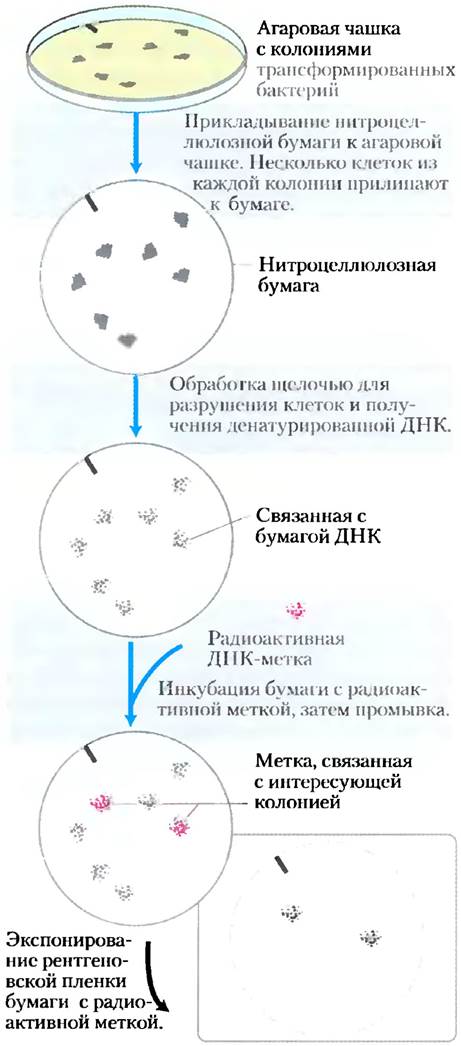
Гибридизация ДНК, о которой в общих чертах говорилось в гл. 8 (см. рис. 8-29), является наиболее доступным специфичным к последовательности нуклеотидов методом детектирования отдельных генов или сегментов нуклеиновых кислот. Существует много разновидностей основного метода, использующего, главным образом, меченые (например, радиоактивные) фрагменты ДНК или РНК (называемые зондами), комплементарные искомой ДНК. Согласно классическому подходу, для обнаружения определенной последовательности ДНК в клонотеке (совокупность клонов ДНК) в агаровую чашку, содержащую множество отдельных бактериальных колоний из клонотеки, с различной рекомбинантной ДНК в каждой, помещают нитроцеллюлозную бумагу. Несколько клеток из каждой колонии прилипают к бумаге, образуя реплику. Затем бумагу обрабатывают щелочью для разрушения клеток и денатурации ДНК; молекулы ДНК остаются связанными бумагой вблизи колоний, откуда они были получены. Добавленный радиоактивный ДНК-зонд связывается только с комплементарной ему ДНК. После вымывания несвязавшихся ДНК-зондов гибридизованную таким образом ДНК можно обнаружить авторадиографией (рис. 9-8).
Рис. 9-8. Использование гибридизации для идентификации клона с определенным сегментом ДНК. Радиоактивный ДНК-зонд гибридизируется с комплементарной ему ДНК, а затем обнаруживается авторадиографией. После идентификации меченых колоний соответствующие им колонии на исходной агаровой чашке можно использовать в дальнейших экспериментах в качестве источника клонируемой ДНК.
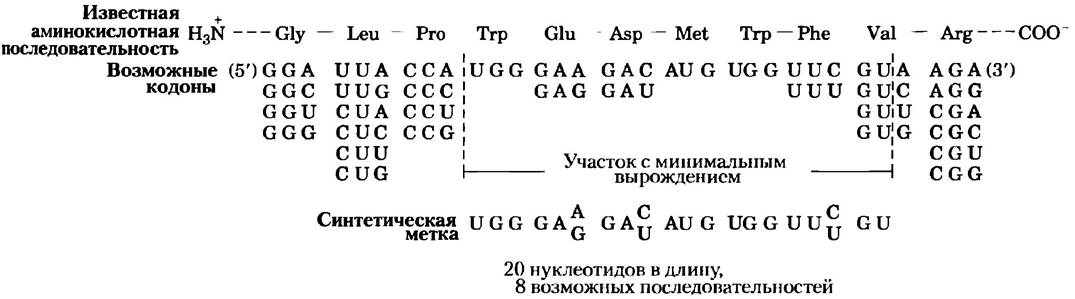
Как правило, создание комплементарной нуклеотидной цепи, используемой в качестве зонда, является лимитирующим шагом при определении и клонировании гена. Природа зонда зависит от того, что известно об исследуемом гене. Иногда подходящим зондом является гомологичный ген, клонированный из других видов. Или, если уже был получен белковый продукт гена, зонд можно спроектировать и синтезировать, исходя из аминокислотной последовательности (рис. 9-9). На сегодняшний день всю необходимую информацию о последовательности ДНК исследователи, как правило, получают из баз данных нуклеотидных последовательностей, в которых подробно описана структура миллионов генов широкого диапазона организмов.
Рис. 9-9. Метка для определения гена, кодирующего белок с известной нуклеотидной последовательностью. Из-за того, что более одной последовательности ДНК может кодировать любую данную аминокислотную последовательность, говорят, что генетический код «вырожден» (об этом говорится в гл. 27; каждая аминокислота кодируется набором из трех нуклеотидов — кодоном. Большинству аминокислот соответствуют два или более кодонов; см. рис. 27-7). Таким образом, искомая последовательность ДНК для известной аминокислотной последовательности не может быть заранее определена. Зонд проектируется так, чтобы быть комплементарным участку гена с минимальным «вырождением», т. е. участку с минимальным числом возможных кодонов для аминокислот; в приведенном здесь примере не более двух кодонов. Олигонуклеотиды синтезируются с селективно рандомизированными последовательностями, так что они содержат один из двух возможных нуклеотидов в каждом месте потенциального вырождения (окрашены в розовый цвет). Приведенный здесь олигонуклеотид представляет собой совокупность из восьми различных последовательностей, при этом одна из них полностью комплементарна гену, а все восемь вместе соответствуют, по крайней мере, 17 из 20 позициям.
Экспрессия клонированных генов дает значительное количество белка
Зачастую основной интерес представляет продукт клонированного гена, а не сам ген, особенно когда белок обладает коммерческой, терапевтической или исследовательской ценностью. Теперь, когда основы метаболизма и регуляции ДНК, РНК и белков в Е. coli становятся все более понятными, исследователи уже могут манипулировать клетками для экспрессии клонированных генов с целью изучения их белковых продуктов.
Большинство генов эукариот лишены элементов последовательности ДНК, необходимых для их экспрессии в клетках Е. coli (таких как промоторы — последовательности, указывающие ДНК-полимеразе места связывания). Поэтому в подходящие для эукариотических генов места в ДНК вектора должны быть помещены бактериальные регуляторные последовательности транскрипции и трансляции. Промоторы, регуляторные последовательности и другие аспекты регуляции генной экспрессии обсуждаются в гл. 28. В некоторых случаях клонированные гены экспрессируются так эффективно, что их белковый продукт составляет более 10% всего клеточного белка; тогда говорят, что они гиперэкспрессированы. При таких концентрациях некоторые чужеродные белки могут убить клетку Е. coli, так что экспрессию гена надо ограничить несколькими часами до сбора клеток.
Клонирующие векторы с сигналами транскрипции и трансляции, необходимыми для регулируемой экспрессии клонированного гена, часто называются экспрессирующими векторами. Скорость экспрессии клонированного гена контролируется при замене собственных промоторных и регуляторных последовательностей гена на более эффективные и подходящие варианты, предоставляемые вектором. Как правило, хорошо охарактеризованный промотор и его регуляторные элементы располагаются вблизи нескольких специфических участков рестрикции, используемых при клонировании, так, чтобы гены, вставляемые по этим участкам, экспрессировались регулируемым промоторным элементом (рис. 9-10). Некоторые из таких векторов включают в себя и другие характерные участки, например, место связывания бактериальной рибосомы для улучшения трансляции мРНК, полученной с гена, или последовательность окончания транскрипции.
Рис. 9-10. Последовательности ДНК в типичном экспрессирующем векторе Е. coli. Экспрессируемый ген вставляется на одном из участков рестрикции поли- линкера вблизи промотора (Р) так, чтобы ближайшим к промотору был конец, кодирующий N-концевую аминокислотную последовательность. Промотор способствует эффективной транскрипции вставленного гена, а последовательность терминатора транскрипции иногда увеличивает количество и стабильность получаемой мРНК. Оператор (0) осуществляет регуляцию с помощью репрессора который связывается с ним (гл. 28). Участок связывания рибосомы обеспечивает сигналы, необходимые для эффективной трансляции мРНК, полученной от гена. Селектируемый маркер позволяет отобрать клетки, содержащие рекомбинантную ДНК.

Гены можно также клонировать и экспрессировать в эукариотических клетках; обычно в качестве хозяев используют различные виды дрожжей. Эукариотический хозяин иногда может осуществлять посттрансляционные модификации (изменения в белковой структуре, происходящие после синтеза на рибосомах), которые могут потребоваться для функционирования клонированного белка эукариот.
Изменения в клонированных генах приводят к получению модифицированных белков
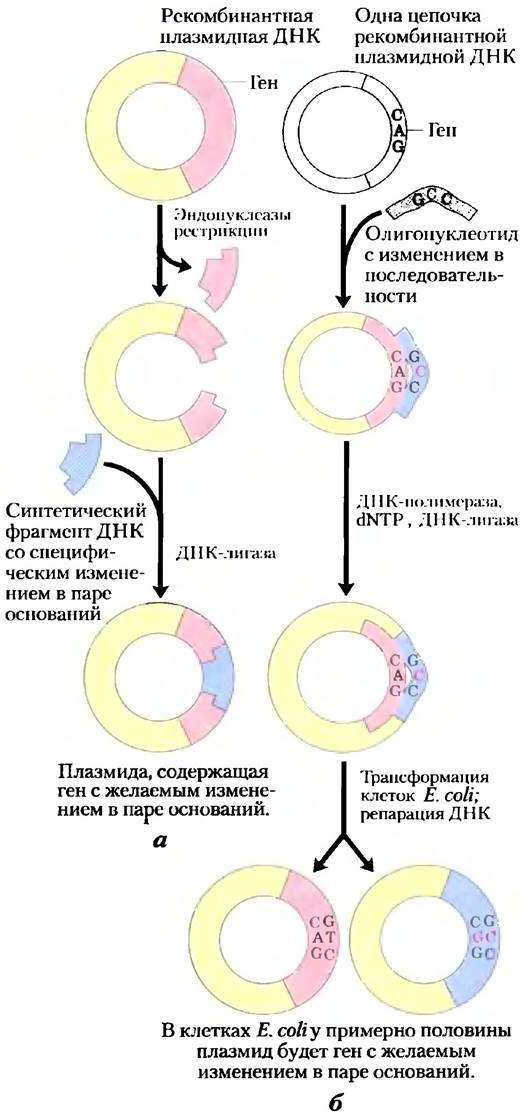
Технологии клонирования можно использовать не только для получения больших количеств белка, но и для синтеза белковых продуктов, слегка отличающихся от их природных форм. Специфические аминокислоты можно заменять по отдельности, используя сайт-специфичный мутагенез. В этом мощном методе изучения строения и функций белка производят изменение аминокислотной последовательности путем модификации последовательности ДНК в клонируемом гене. Если подходящие участки рестрикции расположены по обе стороны от модифицируемой нуклеотидной последовательности, тогда исследователи могут просто вырезать сегмент ДНК и заменить его синтетическим аналогом, идентичным оригиналу за исключением желаемого изменения (рис. 9-11, а). В том случае, если удобно расположенных участков рестрикции нет, можно произвести изменения в специфической ДНК-последовательности при использовании подхода, называемого олигонуклеотид-направленным мутагенезом (рис. 9-11, б). Короткая синтетическая цепочка ДНК со специфическим изменением в основании гибридизуется с одноцепочечной копией клонируемого гена в подходящем векторе. Несоответствие одной пары оснований на 15-20 пар не препятствует гибридизации, если это происходит при подходящей температуре. Эта цепочка служит праймером для синтеза цепи, комплементарной нлазмидному вектору. Затем такая двойная рекомбинантная плазмида с небольшим несоответствием (мисматчем, от англ, mismatch) используется для трансформации бактерий, в которых мисматчи исправляются клеточными ферментами репарации ДНК (гл. 25). Около половины актов репарации будут удалять и заменять модифицированное основание, возвращая ген к его исходной последовательности; другая половина удалит и заменит нетронутое основание, сохраняя желаемую мутацию. Трансформированные клетки тщательно проверяются (часто секвенированием их плазмидной ДНК), пока не будет найдена бактериальная колония, содержащая плазмиду с измененной последовательностью.
Рис. 9-11. Два подхода к сайт-специфичному мутагенезу. а) Синтетический сегмент ДНК заменяет фрагмент ДНК, который был удален вырезанием эндонуклеазой рестрикции. б) Синтетический олигонуклеотид с желаемым изменением последовательности в одном месте гибридизуется с одноцепочечной копией модифицируемого гена и действует как затравка для синтеза двухцепочечной ДНК (с одним мисматчем), которая затем используется для трансформации клеток. Репарирующие системы клеточной ДНК исправляют около 50% несоответствия отражая желаемое изменение в нуклеотидной последовательности.
Изменения можно также осуществить более чем в одной паре оснований. Большие участки гена могут быть удалены вырезанием сегмента эндонуклеазами рестрикции и сшиванием оставшихся частей с образованием меньшего по размерам гена. Части двух различных генов можно сшить, образуя новые комбинации. Продукт такого смешанного гена называется слитым белком (фьюжн-белком).
Теперь исследователи располагают оригинальными методами для осуществления практически любого генетического изменения in vitro. Встраивание измененной ДНК в клетку позволяет изучать результаты такого изменения. Сайт- специфичный мутагенез чрезвычайно продвинул исследование белков, поскольку дал возможность ученым производить специфические изменения в первичной структуре белка и изучать эффекты таких изменений на конформацию, трехмерную структуру и активность белка.
Концевые последовательности обеспечивают участки связывания для аффинной хроматографии
Аффинная хроматография является одним из наиболее эффективных методов очистки белка (см. рис. 3-17, в). К сожалению, для многих белков не найдено лигандов, которые можно было бы иммобилизовать на твердом носителе для проведения хроматографии. Создание слитых белков делает возможным проводить очистку методом аффинной хроматографии практически любого белка.
Сначала создают конструкцию ДНК, в которой ген искомого белка соединен с геном, кодирующим пептид или белок, связывающийся с высоким сродством и специфично с известным лигандом. Пептид или белок, используемый для этой цели, присоединяется к N- или С-концу; его называют концевой последовательностью, или тэгом (от англ. tag— метка, ярлык). В табл. 9-3 перечислены некоторые белки и пептиды, наиболее часто используемые в качестве концевых последовательностей, а также их лиганды.
Таблица 9-3. Часто используемые белковые тэги
Тэг/пептид |
Молекулярная масса, кДа |
Иммобилизованный лиганд |
Белок А |
59 |
Fc-участок IgG |
(His)6 |
0.8 |
Ni2+ |
Глутатнон-S- трансфераза |
26 |
Глутатион |
Мальтозосвязывающий белок |
41 |
Мальтоза |
β-Талактозидаза |
116 |
n-Аминофенил-β-D-тиога-лактозидаза (TPEG) |
Хитинсвязывающий домен |
5.7 |
Хитин |
Ход эксперимента проиллюстрирован на примере присоединения к выделяемому белку последовательности глутатион-5-трансферазы (ГСТ). ГСТ — небольшой фермент (Мr= 26 000), который с высоким сродством и специфичностью связывается с глутатионом (рис. 9-12). Если создать конструкцию, в которой ген ГСТ слит с геном искомого белка, такой химерный белок приобретет способность связывать глутатион. Химерный белок экспрессируют в бактериальных или других клетках, а затем готовят грубый клеточный экстракт. Хроматографическую колонку заполняют пористым носителем, представляющим собой микроскопические шарики из стойкого полимера (например, из перекрестно-сшитой агарозы), на котором иммобилизован лиганд (в данном случае глутатион). Все другие белки из экстракта проходят через колонку, не связываясь с носителем. ГСТ прочно связывается с глутатионом, но связь эта нековалентная, поэтому белок легко можно смыть с колонки с помощью раствора, содержащего либо высокую концентрацию соли, либо свободный глутатион, который конкурирует с иммобилизованным лигандом за связывание с ГСТ. Таким путем часто удается получить очищенный белок с высоким выходом. Иногда концевую последовательность частично или полностью отделяют от очищенного белка с помощью протеазы, которая расщепляет последовательность в области связывания с глутатионом.
Рис. 9-12. Использование химерных конструкций для очистки целевого белка. В качестве примера рассматривается очистка белка, слитого с глутатион-S-трансферазой (ГСТ). а) ГСТ представляет собой небольшой фермент (изображенный здесь в виде кружка фиолетового цвета), который связывает глутатион. (Глутатион — трипептид y-глутамилцистеинилглицин, содержащий необычную пептидную связь между аминогруппой цистеина и углеродом карбоксильной группы боковой цепи глутамата.) б) ГСТ присоединяют к С-концу выделяемого белка генно- инженерным методом. Химерный белок экспрессируется в хозяйской клетке и присутствует в грубом экстракте после лизиса клеток. Экстракт наносят на хроматографическую колонку с носителем, на котором иммобилизован глутатион. Химерный белок связывается с глутатионом и задерживается на колонке, тогда как остальные белки быстро вымываются. Химерный белок затем смывают с колонки с помощью раствора, содержащего высокую концентрацию соли или свободный глутатион.
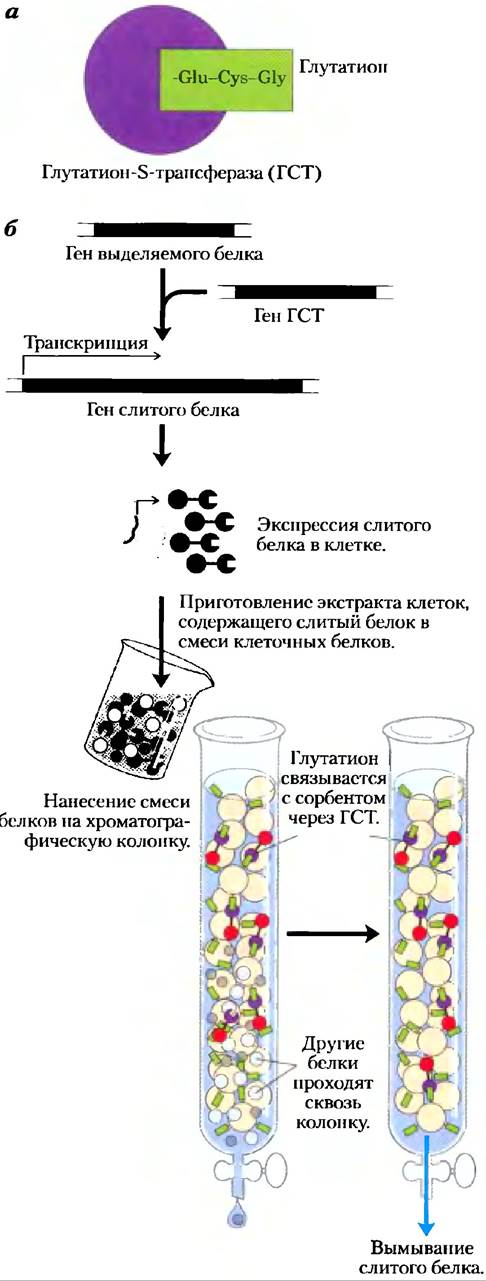
Примером короткой концевой последовательности, которая находит множество применений, может служить простая последовательность из шести или большего числа остатков гистидина (6 x His). Эта последовательность с высоким сродством и специфичностью связывается с ионами никеля. Колонка с иммобилизованными ионами никеля может применяться для эффективного отделения белка с гистидиновой последовательностью на конце от остальных белков в смеси. Более крупные концевые последовательности, такие как мальтозосвязывающий белок, могут улучшить растворимость и повысить устойчивость выделяемых белков, что позволяет выделять белки, которые не удается выделять другими методами.
Методы с использованием концевых последовательностей удобны и эффективны, что объясняет их очень широкое распространение. Однако в этой работе следует проявлять определенную осторожность. Дело в том, что концевые последовательности не являются инертными. Даже очень небольшие последовательности могут изменять свойства белков, к которым они прикреплены, и влиять на результаты эксперимента. Активность белка может измениться даже после удаления концевой последовательности с помощью протеазы, например, если один или несколько аминокислотных остатков остаются связанными с выделяемым белком. Экспериментальные результаты, полученные таким методом, всегда следует оценивать с помощью хорошо продуманного контроля, чтобы учесть влияние посторонней последовательности на функции изучаемого белка.
Краткое содержание раздела 9.1 Клонирование ДНК: основные понятия
■ В клонировании ДНК и генной инженерии используют разрезание ДНК и объединение сегментов ДНК в новую комбинацию — рекомбинантную ДНК.
■ Клонирование подразумевает разрезание ДНК ферментами на фрагменты; выбор и возможность модификации интересующего фрагмента; вставку фрагмента ДНК в подходящий клонирующий вектор; перенос вектора со вставленной в него ДНК в клетку- хозяина для репликации; идентификацию и отбор клеток, содержащих фрагмент ДНК.
■ Ключевые ферменты в клонировании гена — эндонуклеазы рестрикции (особенно ферменты типа II) и ДНК-лигаза.
■ Клонирующими векторами являются плазмиды, бактериофаги, для самых длинных вставок ДНК — искусственные бактериальные хромосомы (ВАС) и искусственные хромосомы дрожжей (YAC).
■ Клетки, содержащие определенные последовательности ДНК, можно идентифицировать методами ДНК-гибридизации.
■ Методы генной инженерии манипулируют клетками для экспрессии и/или изменения клонируемых генов.
■ Белки или пептиды могут быть генно- инженерным методом связаны с целевым белком, в результате чего образуется химерный белок. Дополнительный пептидный участок может быть использован для обнаружения целевого белка или для его очистки методом аффинной хроматографии.